
Abstract
The effects of reduced tidal flushing on post-hurricane mangrove recovery were measured across a gradient of hurricane disturbance (in order of decreasing wind intensity: Captiva, North Sanibel, Central Sanibel, and East Sanibel). Each region consisted of replicate study plots with either reduced tidal exchange (tidally restricted location) or an open tidal connection (tidally unrestricted location). Locations with reduced tidal exchange displayed significantly lower (two-way ANOVA, p ≤ 0.0001) tidal amplitude, decreased seedling densities, and decreased productivity (recruitment, growth, and litter fall) when compared to the tidally unrestricted locations. Results also indicated significant regional variations in measures of mangrove stand structure (seedlings and canopy) and productivity (recruitment, growth, and litter fall) up to 4-years post-hurricane disturbance. These findings suggest that the legacy effects from hurricane disturbance vary with degree of wind intensity, acting both independently and synergistically with the effects of tidal restriction to influence post-hurricane mangrove structure and function.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
Mangrove structural and functional attributes reflect ecosystem productivity, and can be limited by environmental stressors (Lugo 1980; Ellison and Farnsworth 1996; Duke et al. 1998; Twilley et al. 1999; Kathiresan and Bingham 2001). Because of the high metabolic demands associated with growth and reproduction, it is possible that natural disturbances, in combination with anthropogenic disturbance, can cause enough changes in structure and function to lead to an altered steady state (Everham and Brokaw 1996), particularly when ecosystems experience frequent overlapping disturbances. Ultimately, the frequency, intensity, duration, and spatial extent of each disturbance dictates the characteristics that control ecosystem recovery (Boose et al. 1994; Lugo and Scatena 1996), but the legacy effects of a disturbance can leave a system more vulnerable to additional disturbances (Kathiresan and Bingham 2001) and decrease resiliency.
Depending on the frequency and intensity, hurricane disturbances can have catastrophic effects on mangrove communities (Lugo and Scatena 1996). Both mortality and recovery from wind disturbance have a large temporal range lasting weeks to decades (Everham and Brokaw 1996). During hurricane and tropical storm events, mangrove stands can be lost or severely damaged by intense winds and storm surge (Sadd et al. 1999; Lugo 2000; Granek and Ruttenberg 2007). Hurricanes directly impact mangroves through defoliation, uprooted trees, subsidence, broken branches, and eventual mortality (Imbert et al. 1996). This form of direct damage typically decreases with distance from the hurricane eye-wall (Doyle et al. 1995; Milbrandt et al. 2006). In general, the severity of a disturbance in forested areas is measured by structural or compositional loss (Everham and Brokaw 1996) and is governed by biotic (e.g., stand conditions) and abiotic site factors (e.g., storm intensity and disturbance history). Measures of seedling and stem densities are typically used to determine post-hurricane mangrove stand structure (Baldwin et al. 2001; Ross et al. 2006), while seedling recruitment patterns reflect the functionality of mangrove reproduction and productivity (Proffitt et al. 2006). Post-hurricane mangrove ecosystem resiliency is dependent, in part, on initial post-disturbance forest structure, nutrient availability, and post-disturbance seedling recruitment rates (Chen and Twilley 1998), with a lag time noted in both mortality (Everham and Brokaw 1996; Baldwin et al. 2001) and recovery (Proffitt et al. 2006).
Reduced tidal flushing is also a potential stressor to mangrove ecosystems (Lugo and Snedaker 1974). Hydrologic alterations can redirect natural water flow, decrease the degree of tidal inundation, and create habitat fragmentation (Milbrandt et al. 2006). Tidal restriction occurs when obstructions restrict the natural flow dynamics and reduce or manipulate tidal exchange; roadways, dams, and reservoirs are examples of such obstructions (Boumans et al. 2002). Modification of local hydrographic patterns affects mangrove propagule dispersal, seedling establishment, and growth (Ellison and Farnsworth 1996; Duke et al. 1998; Milbrandt and Tinsley 2006), while drainage and entrained water stress decrease mangrove litter fall and productivity (Carter et al. 1973). An initial study by Milbrandt et al. (2006) investigated the effects of tidal restrictions and found that areas with an obstruction or other barrier to impede tidal flushing also had significantly lower Rhizophora mangle seedling densities and other structural differences.
The purpose of this research was to determine whether these obstructions (e.g., roadways, water retention ditches) caused decreased tidal amplitude using simultaneous measurements of water level at both tidally restricted and unrestricted sites. Secondly, mangrove ecosystem structure and function were expected to differ in tidally restricted locations due to decreases in net primary productivity associated with reduced tidal flushing (Carter et al. 1973; Twilley et al. 1986). Finally, post-hurricane mangrove recovery was expected to reflect the degree of disturbance along a decreasing wind intensity gradient, with slower recovery in regions that had experienced intense hurricane winds and a more rapid recovery in areas that had experienced minimal hurricane winds.
Previous research has documented the structural damage caused by hurricane disturbance, but the dynamics of hurricane recovery are not well understood. The alteration of hydrology on a Florida barrier island and the recent hurricane landfall (Hurricane Charley in 2004) created a unique opportunity to study the effects of hydrological impairment and a regional post-disturbance legacy effects on mangrove ecosystem recovery. The effects of reduced tidal variability on mangrove structure (seedlings and canopy) and function (productivity, recruitment, etc.) were determined along a post-hurricane disturbance gradient. Tidal amplitude was used as a proxy for tidal restriction (tidally restricted as having lower tidal amplitude and reduced or manipulated tidal exchange). Seedling densities, seedling recruitment, growth rates, litter fall, and canopy closure were compared by wind disturbance and tidal restriction to determine ecosystem health and productivity, while tidal amplitude was used to quantify the degree of tidal restriction.
Materials and Methods
Study Location
This study was conducted on Sanibel and Captiva Islands, in southwest Florida. Sanibel and Captiva are a part of the Caloosahatchee estuary, surrounded by Pine Island Sound, San Carlos Bay, and the Caloosahatchee River to the east, and the Gulf of Mexico to the west. Pine Island Sound and San Carlos Bay are estuarine waters with salinities dependent upon freshwater inflow from the Caloosahatchee River watershed, which varies with seasonal precipitation patterns and freshwater releases from Lake Okeechobee. Because there is no clear source of freshwater found on Sanibel or Captiva Islands, the freshwater influence on Sanibel–Captiva’s wetlands depends on local precipitation patterns, rather than freshwater discharge (Clark 1976). Average rainfall on Sanibel has been recorded at 127 cm/year mostly occurring in the wet season (June–October) and a mean tidal range of 47 cm (NOAA 2009). Since the early 1900s, both Sanibel and Captiva have been subject to human population, settlement, and infrastructure development, including impoundment, drainage, or land use alteration, which led to the degradation of submerged aquatic vegetation, oyster reefs, and tidal wetlands (Clark 1976; Corbett 2006; Meyers et al. 2006). Tidal wetlands in the study area are principally composed of mangroves and contain a mix of Rhizophora mangle (red mangrove), Avicennia germinans (black mangrove), and Laguncularia racemosa (white mangrove) (Table 1).
Experimental Design
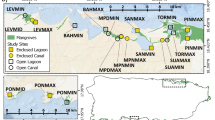
Hurricane Charley, a category 4 storm (Saffir–Simpson hurricane scale), made landfall north of Captiva, FL, on August 13, 2004 causing wind damage to all forested areas and providing an opportunity to study mangrove damage and subsequent recovery. Study plots were established following a 2 × 4 factorial design to compare measures of structure and function by tidal restriction (unrestricted and restricted) and region (Captiva, North Sanibel, Central Sanibel and East Sanibel). The four regions experienced decreasing hurricane wind intensity from Captiva to East Sanibel (Fig. 1). Captiva was the closest region to the hurricane eye-wall, and endured the greatest hurricane wind damage (Milbrandt et al. 2006). Following Captiva, wind disturbance (as assessed from change in canopy pre- and post-hurricane) decreased moving further away from the hurricane eye-wall, with North Sanibel, then Central Sanibel, and East Sanibel, the lowest disturbance and the furthest region from the hurricane eye-wall (Fig. 1). Each region consisted of one tidally restricted and one tidally unrestricted location, described below. Locations had three replicate plots (6-m radius) in which all measurements were taken. Plots were established in 2003 and have been surveyed annually until 2007, as described by Milbrandt et al. (2006).

Hurricane Charley (2004), location of the eye-wall and studied regions. Captiva (CT) was the closest region to the hurricane eye-wall followed by North Sanibel (NS), Central Sanibel (CS), and East Sanibel (ES) (Milbrandt et al. 2006)
In all regions, tidally ‘unrestricted’ plots were located landward of open water with no barriers blocking tidal flow. Tidally ‘restricted’ locations were in areas bisected by a roadway and were thought to have decreased tidal amplitude. Each tidally restricted location displayed an observable impoundment (via roadway) and reduced tidal exchange when compared to the tidally unrestricted counterpart. The study plots all had historical connections to open water through tidal wetlands, bayous, and backwater creeks and were characterized as mixed species stands (R. mangle, A. germinans, and L. racemosa); specific species distributions between regions and locations are further described in Table 1.
The first objective of this study was to quantify tidal flushing, as there were patterns detected in the stand structure that suggested functional differences between tidally restricted and unrestricted locations (Milbrandt et al. 2006). High and low tides were measured during a spring tide in January 2008 using temporary observational piezometers (Ridd et al. 1997; Hughes et al. 1998) positioned at a random distance and bearing from the center of 6-m2 plots. Low tides were recorded simultaneously by measuring from the top of the piezometers down to the water table every 5 min for approximately 1 h after the predicted low tide. High tide measurements were taken by applying Kolor Kut (Channel Supplies, TX, USA) water finding paste to each piezometer, then measuring from the top of the piezometer to the highest high tide mark. The difference between the maximum high and minimum low tides were averaged per location (n = 9, restricted; n = 10, unrestricted) and are expressed as tidal amplitude.
In 2006, one permanent 1-m2 quadrat was randomly selected (bearing and distance from central pin) within each 6-m2 plot (n = 13, restricted; n = 13, unrestricted) to estimate seedling recruitment. Seedling recruitment was calculated by annually tagging individual seedlings with color coded zip ties in 1-m2 permanent quadrats and enumerating the new seedlings established the previous year; during the 2007 and 2008 site visits, new unmarked seedlings were counted and tagged to represent the number of new recruits per 2006 and 2007 reproductive seasons. Seedling densities were also recorded in each 6-m2 plot for all species and calculated as the average (one permanent and eight random) per 1-m2 quadrat.
Diameter of stem at breast height (DBH; 1.3 m) was measured for all mangrove stems greater than 1 cm DBH within each 6-m2 plot (Milbrandt et al. 2006). In 2006, stems were individually tagged for identification and marked at DBH. Stems were then re-measured in 2007 and relative growth rate (RGR) per individual was calculated. RGRs were calculated to standardize growth by stem size (Sherman et al. 2000). RGRs were determined using the following equation:
Mangrove productivity was estimated using rates of litter fall (Twilley et al. 1986; Day et al. 1996). One litter trap was positioned at a random distance and bearing from the center of each 6-m2 plot; a total of 26 litter traps were established in the summer of 2007. Litter collection boxes were constructed of 25 cm by 25 cm plywood, underlain with 2 mm mesh screening to catch an area of 0.0625 m2 of litter fall. The litter collection boxes were xelevated on wooden posts, leaving the litter trap approximately 0.85 m above ground. Litter fall was collected five times from spring 2007 to spring 2008, which falls within the monthly (Twilley et al. 1986; Day et al. 1996) and quarterly (Ellison and Simmons 2003) sampling procedures for mangrove litter fall collection. Within 1 week of collection, litter fall (including leaves, stems, reproductive parts, and misc.) was taken to the laboratory and dried at 70°C for 72 h (Twilley et al. 1986; Day et al. 1996). The dried mass was weighed and dry weight (DW) per area was determined for each plot. All samples were placed in Thermolyne 1400 (Barnstead International, IA, USA) muffle furnace for 3 h at 500°C (Mfilinge et al. 2002; Bouchard et al. 2003), and percent organic matter (OM) was calculated. All DW and OM are reported in g m−2 year−1.
Canopy closure was recorded at each site in 2007 using a spherical densitometer (Lemmon 1956). Canopy closure was measured at the center of each plot. Canopy closure was averaged from north, south, east, and west heading, and is reported as a percent canopy closure (includes leaves, branches, or other obstructions).
Data Analysis
All statistical analyses were conducted using SPSS 12.0. Tidal amplitude, seedling densities, seedling recruitment, growth rates, litter fall, and canopy closure were considered dependent variables and analyzed against two independent variables, tidal influence (restricted and unrestricted) and region (Captiva, North Sanibel, Central Sanibel, and East Sanibel). Data were analyzed for tidally restricted and unrestricted locations, all regions, all plots. Normality of the dataset was tested using the Shapiro–Wilk test. Homogeneity of variance was tested by residual analysis. If the assumptions of normality and homogeneity were not violated then a two-way ANOVA was used to compare tidal influence and regions with a Scheffé post hoc test to view specific significant differences among regions. If data was heteroscedastic, then the data was square-root (√) transformed and analyzed with a two-way ANOVA. If data were still found to be not normally distributed, then analysis was conducted via nonparametric tests. For non-normally distributed data, the Wilcoxon rank sum nonparametric test and descriptive statistics were used to look at differences in tidal influence, and a Kruskal–Wallis test was used to compare the four regions of hurricane intensity. Averages are expressed as the mean ± Standard Error (SE).
Results
Interaction
The interaction of tidal influence by region was examined for measures of tidal amplitude, seedling densities, seedling recruitment, litter fall, and canopy closure. For each measure, the tidal influence by region interaction was not significant (tidal amplitude, two-way ANOVA, p = 0.909; seedling densities, two-way ANOVA, p = 0.354; seedling recruitment (2007) two-way ANOVA, p = 0.180; DW of litter fall, two-way ANOVA, p = 0.059; canopy closure, two-way ANOVA, p = 0.249), indicating that the differences due to tidal restriction did not vary regionally.
Tidal Influence
Measures of tidal amplitude were consistently lower in tidally restricted locations (Fig. 2). Tidally restricted locations had significantly (two-way ANOVA, p ≤ 0.0001) lower tidal amplitude than unrestricted locations, with an average 10.61 cm lower tidal amplitude in restricted locations. Measurements of R. mangle, A. germinans, and L. racemosa seedling densities were also significantly (two-way ANOVA, p ≤ 0.0001) lower in tidally restricted locations (Fig. 3a). However, there was no significant (M–W test, p = 0.113) difference between mean 2006 seedling recruitment in tidally restricted versus unrestricted locations (Table 2). Mean 2007 seedling recruitment was significantly (two-way ANOVA, p = 0.003) lower in tidally restricted locations than in the corresponding unrestricted location (Fig. 3b). Recruitment in unrestricted locations increased from 16.6 ± 3.9 seedlings m−2 to 28.2 ± 6.2 seedlings m−2 from 2006 to 2007 (69.4%), while the restricted locations increased from 6.9 ± 2.1 seedlings m−2 to 9.1 ± 4.0 seedlings m−2 from 2006 to 2007 (31.2%) over the same period (Table 2).

Tidal amplitudes (mean ± SE) from unrestricted ( ) and restricted (
) and restricted ( ) locations are displayed for each of the four regions. X-axis represents increased distance from hurricane eye-wall and decreased wind intensity. Captiva (CT) was the closest region to the hurricane eye-wall followed by North Sanibel (NS), Central Sanibel (CS), and East Sanibel (ES)
) locations are displayed for each of the four regions. X-axis represents increased distance from hurricane eye-wall and decreased wind intensity. Captiva (CT) was the closest region to the hurricane eye-wall followed by North Sanibel (NS), Central Sanibel (CS), and East Sanibel (ES)

Mean seedling densities a seedling recruitment b dry weight (DW) of litter fall c and percent canopy closure d in 2007 from unrestricted ( ) and restricted (
) and restricted ( ) locations. The four regions represent a disturbance gradient where Captiva (CT) was the closest region to the hurricane eye-wall followed by North Sanibel (NS), Central Sanibel (CS), and East Sanibel (ES). The X-axis, from right to left represents increased distance from hurricane eye-wall and decreased disturbance. Error bars are ±1 standard error (SE)
) locations. The four regions represent a disturbance gradient where Captiva (CT) was the closest region to the hurricane eye-wall followed by North Sanibel (NS), Central Sanibel (CS), and East Sanibel (ES). The X-axis, from right to left represents increased distance from hurricane eye-wall and decreased disturbance. Error bars are ±1 standard error (SE)
Mean RGRs of 0.08 cm year−1 and 0.03 cm year−1 were calculated for tidally unrestricted and restricted locations, respectively. Mean RGRs were significantly (M–W test, p ≤ 0.0001) lower in tidally restricted locations (Table 3). Mean DW of the litter fall was significantly (two-way ANOVA, p ≤ 0.0001) lower in tidally restricted locations than in the corresponding unrestricted location (Fig. 3c). OM was similar for both restricted (87.8%) and unrestricted (86.5%) locations (Table 4), as expected given similar species compositions (Table 1). Because there were few to no surviving mangrove stems in North Sanibel’s restricted plots, the canopy closure data from that region was excluded to provide a more meaningful comparison. There were no significant differences in canopy closure between tidally restricted and unrestricted locations (two-way ANOVA, p = 0.799).
Regions
Significant (two-way ANOVA, p = 0.001) differences in tidal amplitude were observed among regions. Measurements of tidal amplitude exhibited a regional trend of decreasing mean tidal amplitude from Captiva to East Sanibel, with each of the unrestricted locations showing greater tidal amplitude than the corresponding restricted locations (Fig. 2). Mean tidal amplitude decreased from north to south so that regions with the greatest disturbance from the 2004 hurricane also had the greatest tidal amplitude, while the least disturbed regions had also had the least tidal amplitude (Fig. 2).
Seedling densities were higher in areas with minor hurricane disturbance, while lower densities were found in areas with a greater degree of hurricane damage (two-way ANOVA, p ≤ 0.0001) (Fig. 3a). There were significant (K–W test, p = 0.005) regional differences in 2006 seedling recruitment, but there were no significant (two-way ANOVA, p = 0.291) regional differences in 2007 seedling recruitment (Table 5). Captiva showed the largest increase in seedling recruitment (260.5%) from 5.5 seedlings m−2 in 2006 to 19.8 seedlings m−2 in 2007 (Table 5). East Sanibel’s seedling recruitment decreased (−7.3%) from 22.8 seedlings m−2 in 2006 and 21.2 seedlings m−2 in 2007 (Table 5). RGRs revealed significant (K–W test, p = 0.003) differences among regions (Table 6). Central Sanibel showed the highest RGRs in unrestricted plots, while Captiva showed the highest RGRs for restricted locations. The mean DW of litter fall in unrestricted locations was lowest in Captiva where hurricane damage was the greatest (Fig. 3c). Tidally restricted locations followed a similar pattern with the exception of North Sanibel where no trees remained standing after the 2004 hurricane. A significant difference (two-way ANOVA, p = 0.003) in DW was seen among regions, demonstrating a regional pattern of increasing mean litter fall from Captiva to East Sanibel (Table 7). Percent canopy closure also revealed significant (two-way ANOVA, p ≤ 0.0001) regional differences (Fig. 3d). Canopy closure in unrestricted, as well as restricted locations, was higher in areas with low to moderate hurricane damage.
The Scheffé post hoc test identified significant differences between Captiva (heavily damaged) and East Sanibel (lightly damaged) for tidal amplitude, seedling densities, 2006 seedling recruitment, DW of litter fall, and percent canopy closure (Fig. 3). Differences were also detected between Captiva and Central Sanibel in tidal amplitude, seedling densities, and canopy closure. North Sanibel and East Sanibel differed in tidal amplitude, seedling densities, 2006 seedling recruitment, DW of litter fall, and percent canopy closure. North Sanibel and Central Sanibel differed in seedling densities and 2006 seedling recruitment. Central Sanibel and East Sanibel differed for percent canopy closure only.
Discussion
Previous research has identified tidal flushing as an important factor regulating mangrove productivity (Carter et al. 1973; Lugo and Snedaker 1974; Twilley et al. 1986). Based on the earlier studies, we would have expected higher productivity in regions with greater tidal amplitude. Analysis confirmed that the average of each measure of mangrove stand structure and productivity was higher in tidally unrestricted plots. However, our results indicated that regional differences in stand structure and productivity appeared to be the result of a regional legacy effect from wind intensity, rather than from differences in tidal amplitude. The gradient of disturbance reported previously from the 2004 hurricane (Milbrandt et al. 2006) was still evident in stand structure and measures of ecosystem function. These results suggest that the legacy effects from wind disturbance are long lasting, leaving the system more susceptible to additional disturbances and creating an opportunity for multiple disturbance interactions.
Three years after hurricane Charley, seedling densities in tidally unrestricted locations (32.0 seedlings m−2) were significantly greater than in restricted locations (10.1 seedlings m2). Seedling densities in unrestricted locations are comparable to the sum of all three mangrove species in South Florida 4 years after Hurricane Andrew (1992) (Ross et al. 2006). There, seedling densities of R. mangle were 9.5 seedlings m−2; L. racemosa were 20.8 seedlings m−2; and A. germinans were 0.5 seedlings m−2 (sum of 30.8 seedlings m−2). Prior to this study, Milbrandt et al. (2006) also observed significantly lower R. mangle seedlings in tidally restricted locations. After more clearly defining ‘tidally restricted’ by measuring tidal amplitude, this pattern continued to apply throughout the current study area. Higher net primary productivity associated with increased tidal inundation has been documented (Carter et al. 1973; Lugo and Snedaker 1974; Twilley et al. 1986) and supports the conclusion that areas with an open tidal connection have greater productivity and greater functional resilience and are therefore more likely to recover from future catastrophic disturbances.
One-year post-hurricane disturbance, minimal R. mangle seedling recruitment was reported within Central Sanibel, demonstrating a lag in reproduction after a hurricane disturbance as observed and predicted by Proffitt et al. (2006). The current study included two additional years of recruitment data indicating a regional increase in recruitment from North to Central to East Sanibel for both the third and fourth reproductive seasons after the 2004 hurricane. Captiva, the most heavily damaged region, had a pulse of recruitment in the fourth reproductive season post-hurricane, suggesting that areas experiencing the most intense hurricane winds were showing signs of recovery, but only after a 3-year lag. Unrestricted locations experienced a greater change in seedling recruitment from the third to fourth post-hurricane reproductive seasons, thus, indicating greater resiliency and potential for long term recovery.
The average RGRs in tidally unrestricted locations (0.08 cm year−1) are analogous to measures of R. mangle under closed canopy (0.086 cm year−1) in the Dominican Republic (Sherman et al. 2000). Sherman et al. (2000) found that all three mangrove species (R. mangle, L. racemosa, and A. germinans) had higher relative growth rates under canopy gaps. Research has suggested that increased salt stress yields reduced mangrove growth (Ball et al. 1997; Duke et al. 1998), while flooded, well-drained soils, support growth (Kathiresan and Bingham 2001). Results from the current study reveal that tidal restriction can also impede growth (lower average RGRs in tidally restricted locations, 0.03 cm year−1). The RGRs did not show the same pattern of regional recovery as measures of seedling densities, litter fall, or canopy closure. Average RGRs increased regionally from Captiva to North Sanibel to Central Sanibel, then decrease in East Sanibel. Crowding has been reported to negatively impact R. apiculata growth rates in Malaysian mangroves (Putz and Chan 1986), and could explain the decreased growth rates found in East Sanibel where stem density is high (unpublished data). East Sanibel also had the greatest percent canopy closure, and while this might have impacted growth, it was not a regional trend. Everham and Brokaw (1996) noted leaf area and reproduction increases in forested areas as a recovery response following catastrophic wind disturbance. When compared to the other measures of productivity, the growth rates recorded in the current study were relatively low, suggesting that during post-hurricane disturbance recovery, more energy is allocated to canopy growth and reproduction, opposed to individual stem growth.
Average annual litter fall was comparable to previous studies (Lugo and Snedaker 1974; Putz and Chan 1986; Twilley et al. 1986; Twilley et al. 1997), falling within the range (130–1,870 g m−2 year−1) established by Kathiresan and Bingham (2001). Litter fall in unrestricted locations (616.18 g m−2 year−1) resembles the mean annual litter fall recorded for mixed species basin stand mangrove forests in Southwest Florida (661 g m−2 year−1, see Twilley et al. 1997). The litter fall in restricted locations (167.13 g m−2 year−1) was much lower and more comparable to the less productive scrub mangroves 186 g m−2 year−1) in Southeast Florida (Twilley et al. 1997). A regional examination of average annual litter fall had similar results with increasing productivity from Captiva (105.5 g m−2 year−1) to East Sanibel (682.67 g m−2 year−1), which supports regional recovery reflecting wind intensity and a reduction in productivity associated with reduced tidal influence.
Recovery time from catastrophic wind damage varies with frequency, and the severity of the disturbance with a lag time noted for mortality (Everham and Brokaw 1996; Baldwin et al. 2001) and recovery (Proffitt et al. 2006). Although each region was impacted by differing degrees of hurricane disturbance, recovery mirrors the degree of disturbance. It has been suggested that recovery from natural disturbance can leave a system more vulnerable to additional disturbances (Kathiresan and Bingham 2001) and when disturbances become more frequent or overlap, the additional stress could throw the system out of balance (Everham and Brokaw 1996). In the current study, legacy effects from the 2004 hurricane increased the vulnerability of mangroves, which had reduced tidal amplitude due to human activities.
The current study suggests that when areas are tidally restricted from road building, mosquito ditching, or other human activities, mangroves are degraded and become less resilient to disturbance events. The results of this study apply to areas impacted by reduced tidal flushing and may not apply to all areas designated as impoundments, especially areas that are monitored for flood control. A previous study of J. N. Ding Darling Wildlife Refuge (Central Sanibel) suggested that when managed for flood control, impounded areas may be functionally degraded but provide an important benefit by serving as wading bird foraging area (Meyers et al. 2006). The current study included measures of mangrove stand structure, productivity, and function but did not examine the use of mangroves as habitat by any specific species. Mangroves provide habitat to wildlife, but additional research is required to determine how structural or functional changes within mangrove systems would impact their role as a nursery ground and habitat.
A better understanding of the interface between natural and anthropogenic disturbance and recovery within mangroves is essential for barrier island pan-tropical coastal management and restoration practices. Mangrove ecosystems face a variety of environmental stressors, including both natural and anthropogenic disturbances. Prolonged stress applied to a mangrove ecosystem may result in an altered system with lower productivity, recruitment, and potential species replacement (Lugo and Snedaker 1974). This study is unique in that the synergistic effects of multiple disturbances, both natural and human-caused, provide insight into the mangrove recovery process. Multiple disturbance interactions must be considered when making management choices that will impact mangrove ecosystems and barrier island systems, especially in the face of future land use changes. Incorporating predictions of land use change, global climate change, and sea level rise is essential to understanding how isolated episodic disturbances events will interact with continual stressors (e.g., decreased tidal amplitude) to affect impending mangrove ecosystem health.
References
Ball, M.C., M.J. Cochrane, and H.M. Rawson. 1997. Growth and water use of the mangroves Rhizophora apiculata and R. stylosa in response to salinity and humidity under ambient and elevated concentrations of atmospheric CO2. Plant, Cell and Environment 20: 1158–1166.
Baldwin, A., M. Egnotovich, M. Ford, and W. Platt. 2001. Regeneration in fringe mangrove forests damaged by Hurricane Andrew. Plant Ecology 157: 149–162.
Boose, E.R., D.R. Foster, and M. Fluet. 1994. Hurricane impacts to tropical and temperate forest landscapes. Ecological Monographs 64: 369–400.
Bouchard, V., D. Gillon, R. Joffre, and J. Lefeuvre. 2003. Actual litter decomposition rates in salt marshes measured using near-infrared reflectance spectroscopy. Journal of Experimental Marine Biology and Ecology 290: 149–163.
Boumans, R.M.J., D.M. Burdick, and M. Dionne. 2002. Modeling habitat change in salt marshes after tidal restoration. Restoration Ecology 10: 543–555.
Carter, M.R., L.A. Burns, T.R. Cavinder, K.R. Dugger, P.C. Fore, D.B. Hicks, H.L. Revells, and T.W. Schmidt. 1973. Ecosystem analysis of Big Cypress swamp and estuaries. U.S. Environmental Protection Agency Report 904/9-74-002 Atlanta, GA.
Chen, R., and R.R. Twilley. 1998. A gap dynamic model of mangrove forest development along gradients of soil salinity and nutrient resources. The Journal of Ecology 86: 37–51.
Clark, J. 1976. The Sanibel Report: formulation of a comprehensive plan based on natural systems. Washington, D.C.: The Conservation Foundation.
Corbett, C.A. 2006. Seagrass coverage changes in Charlotte Harbor, Florida. Florida Scientist 69: 7–23.
Day, J.W., C. Coronado-Molina, F.R. Vera-Herrera, R. Twilley, V.H. Rivera-Monroy, H. Alvarez-Guillen, R. Day, and W. Conner. 1996. A 7 year record of above-ground net primary production in a southeastern Mexican mangrove forest. Aquatic Botany 55: 39–60.
Doyle, T.W., T.J. Smith, and M.B. Robblee. 1995. Wind damage effects of Hurricane Andrew on mangrove communities along the southwest coast of Florida, USA. Journal of Coastal Research 21: 159–168.
Duke, N.C., M.C. Ball, and J.C. Ellison. 1998. Factors influencing biodiversity and distributional gradients in mangroves. Global Ecology and Biogeography Letters 7: 27–47.
Ellison, A.M., and E.J. Farnsworth. 1996. Anthropogenic disturbance of Caribbean mangrove ecosystems: past impacts, present trends, and future predictions. Biotropica 28: 549–565.
Ellison, A.M., and Simmons. 2003. Structure and productivity of inland mangrove stands at Lake MacLeod, Western Australia. Journal of the Royal Society of Western Australia 86: 25–30.
Everham, E.M., and N.L. Brokaw. 1996. Forest damage and recovery from catastrophic wind. The Botanical Review 6: 114–149.
Granek, E.F., and B.I. Ruttenberg. 2007. Protective capacity of mangroves during tropical storms: a case study from Wilma and Gama in Belize. Marine Ecology Progress Series 343: 101–105.
Hughes, C.E., P. Binning, and G.R. Willgoose. 1998. Characterization of the hydrology of an estuarine wetland. Journal of Hydrology 211: 34–49.
Imbert, D., P. Labbe, and A. Rousteau. 1996. Hurricane damage and forest structure in Guadeloupe, French West Indies. Journal of Tropical Ecology 12: 663–680.
Kathiresan, K., and B.L. Bingham. 2001. Biology of mangroves and mangrove ecosystems. Advances in Marine Biology 40: 81–251.
Lemmon, P.E. 1956. A spherical densitometer for estimating forest overstory density. Forest Science 2: 314–320.
Lugo, A.E. 1980. Mangrove ecosystems: sucessional or steady state? Biotropica, supplement: Tropical Succession 12: 65–82.
Lugo, A.E. 2000. Effects and outcomes of Caribbean hurricanes in a climate change scenario. The Science of the Total Environment 262: 243–251.
Lugo, A.E., and F.N. Scatena. 1996. Background and catastrophic tree mortality in tropical moist, wet, and rain forests. Biotropica 28: 585–599.
Lugo, A.E., and S.C. Snedaker. 1974. The ecology of mangroves. Annual Review of Ecology and Systematics 5: 39–64.
Mfilinge, P.L., N. Atta, and M. Tsuchiya. 2002. Nutrient dynamics and leaf litter decomposition in a subtropical mangrove forest at Oura Bay, Okinawa, Japan. Trees 16: 172–180.
Milbrandt, E.C., and M.N. Tinsley. 2006. The role of salt wort (Batis maritima L.) in regeneration of degraded mangrove forests. Hydrobiologia 568: 369–377.
Milbrandt, E.C., J.M. Greenawalt-Boswell, P.D. Sokoloff, and S.A. Bortone. 2006. Impact and response of Southwest Florida mangroves to the 2004 Hurricane Season. Estuaries and Coasts 29: 979–984.
Meyers, J.M., C.A. Langtimm, T.J. Smith III, and K. Pednault-Willett. 2006. Wildlife and habitat damage assessment from Hurricane Charley: recommendations for recovery of the J. N. “Ding” Darling National Wildlife Refuge Complex. U.S. Geological Survey Open File Report 2006-1126. http://sofia.usgs.gov/publications/ofr/2006-1126/index.html Accessed 09 Dec 2009.
NOAA. 2009. Tides & currents. Tarpon Bay, Fl: Station Information. Station ID 8725362. http://www.tidesandcurrents.noaa.gov Accessed on 09 Dec 2009.
Proffitt, C.E., E.C. Milbrandt, and S.E. Travis. 2006. Red mangrove (Rhizophora mangle) reproduction and seedling colonization after Hurricane Charley: comparisons of Charlotte Harbor and Tampa Bay. Estuaries and Coasts 29: 972–978.
Putz, F.E., and H.T. Chan. 1986. Tree growth, dynamics, and productivity in a mature mangrove forest in Malaysia. Forest Ecology and Management 17: 211–230.
Ridd, P., R. Sam, S. Hollins, and G. Brunskill. 1997. Water, salt and nutrient fluxes of tropical tidal saltflats. Mangroves and Salt Marshes 1: 229–238.
Ross, M.S., P.L. Ruiz, J.P. Sah, D.L. Reed, J. Walters, and J.F. Meeder. 2006. Early post-hurricane stand development in fringe mangrove forests of contrasting productivity. Plant Ecology 185: 283–297.
Sadd, S., M.D. Hausain, R. Yaacob, and T. Asano. 1999. Sediment accretion and variability of sedimentological characteristics of a tropical estuarine mangrove: Kemaman, Terengganu, Malaysia. Mangroves and Salt Marshes 3: 51–58.
Sherman, R.E., T.J. Fahey, and J.T. Battles. 2000. Small-scale disturbance and regeneration dynamics in neotropical mangrove forests. Journal of Ecology 88: 165–178.
Twilley, R.R., A.E. Lugo, and C. Ratterson-Zucca. 1986. Litter production and turnover in basin mangrove forests in Southwest Florida. Ecology 67: 670–683.
Twilley, R.R., M. Pozo, V.H. Garcia, V.H. Rivera-Monroy, R. Zambrano, and L. Botero. 1997. Litter dynamics in riverine mangrove forests in the Guayas River estuary, Ecuador. Oecologia 111: 109–122.
Twilley, R.R., V.H. Rivera-Monroy, R. Chen, and L. Botero. 1999. Adapting to ecological mangrove model to simulate trajectories in restoration ecology. Marine Pollution Bulletin 37: 404–419.
Acknowledgements
This research was supported by funds from the Explorers Club of Southwest Florida and Florida Gulf Coast University Office of Research and Sponsored Programs. The authors wish to thank Sanibel–Captiva Conservation Foundation, J. N. Ding Darling National Wildlife Refuge, Rookery Bay National Estuarine Research Reserve, and Florida Gulf Coast University Coastal and Watershed Institute. This is contribution 0019 from the Sanibel–Captiva Conservation Foundation Marine Laboratory.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article
Cite this article
Harris, R.J., Milbrandt, E.C., Everham, E.M. et al. The Effects of Reduced Tidal Flushing on Mangrove Structure and Function Across a Disturbance Gradient. Estuaries and Coasts 33, 1176–1185 (2010). https://doi.org/10.1007/s12237-010-9293-2
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s12237-010-9293-2