
Abstract
The Inclusion and Selection of Medicinal Plants in Traditional Pharmacopoeias—Evidence in Support of the Diversification Hypothesis. An ethnobotanical study with phytochemical analyses was undertaken to examine the medicinal plants used by residents of a small rural community in northeastern Brazil. The present work tested two ideas that attempt to explain the inclusion and selection of medicinal plants in a given culture: the diversification hypothesis and the concept of versatility. The study involved 101 people and used semistructured interviews. A total of 61 plants were selected, including 25 exotic and 36 native species. Plants were classified according to their habit and analyzed for their phytochemical components. In addition, the relative importance (RI) of these plants was calculated, and a chemical diversity index (CDI) was created and applied to each of the species. Exotic and native plants were found to have significantly different occurrences of certain classes of compounds; this result supports the diversification hypothesis. It was therefore concluded that exotic plants are included in traditional pharmacopoeias to fill therapeutic vacancies that native plants cannot satisfy.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Few studies have attempted to address the intercultural exchange of traditional botanical knowledge or the adaptation and evolution of pharmacological traditions over time (Palmer 2004). Nonetheless, various studies have pointed out the expressive richness of exotic plants in traditional pharmacopoeias (Bennett and Prance 2000; Janni and J. W. Bastien, 2004; Palmer 2004), indicating the importance of intercultural contact to the incorporation of new knowledge, traditions, and costumes.
Many authors have attributed the loss of knowledge of medicinal plants to the acculturation of local communities and have indicated modernization and globalization as two important factors responsible for these events (Benz et al. 2000; Caniago and Siebert 1998; Müller-Schwarze 2006; Quinlan and Quinlan 2007). It is important to note, however, that access to modernity is not necessarily a threat to the perpetuation of traditional knowledge. According to Zarger and Stepp (2004), the traditional knowledge is resilient and mutable and can persist in some contexts.
There are communities that seem to have abandoned their knowledge of native plants and traditional values in exchange for greater social and economic opportunities (see Reyes-García et al., 2004 for review). This apparent disinterest in the preservation of traditional knowledge seems to be driven by governmental (and even nongovernmental) institutions that introduce programs for modernization and invest in technological progress (Benz et al. 2000). This phenomenon is complex, however. The construction of a “traditional pharmacopoeia” is quite complex, rich in details, and often not totally comprehensible to researchers. For example, allied to the fact that traditional communities often incorporate exotic species into their knowledge systems and practices (usually fruits of intercultural exchange) is the question of the origins of the resources that are exploited by these communities. A large proportion of the medicinal plants in traditional tropical pharmacopoeias are derived from secondary forests and anthropogenic zones; herbs tend to dominate among the species that are traditionally cited (Albuquerque et al. 2005; Caniago and Siebert 1998; Stepp 2004; Stepp and Moerman 2001; Voeks 1996, 2004; Voeks and Leony, 2004). Explanations for these patterns have been proposed but are still debated (see, for example, Albuquerque and Lucena 2005; Alencar et al. 2009; Almeida et al. 2005; Araújo et al. 2008; Stepp 2004). Curiously, this subject has attracted only limited interest among ethnobotanists and ethnopharmacologists.
The link between exotic plants and anthropogenic zones, which are important sources of medicinal plants, may point to a role for acculturation; however, this explanation is often excessively simplistic, ignoring complex phenomena and patterns and even discouraging more focused investigative actions. The present article contributes to the discussion on use-patterns of medicinal plants in the Caatinga drylands of northeastern Brazil. We examined two current hypothesis for how local cultures select medicinal plants: the versatility hypothesis and the diversification hypothesis.
We examined whether the exotic plants of folk pharmacopoeias have more uses than native plants. Bennett and Prance (2000) suggested that plants that serve a given culture as food sources or ornamentals may eventually be introduced into their pharmacopoeias—in large part due to their use-versatility. Accordingly, we might expect exotic plants to show more use-citations than native plants in medicinal as well as general use categories.
We also addressed whether exotic plants have a greater chemical diversity than native plants, perhaps contributing to a greater diversification of the local pharmacopoeia. Albuquerque (2006) proposed a hypothesis that has not yet been formally tested; this idea attempts to explain the manner in which exotic plants are incorporated into traditional medicinal pharmacopoeias. He argues that exotic plants make their way into traditional pharmacopoeias because they diversify the local repertoire of medicinal plants and enrich the pharmacological stock of those traditional communities. According to this idea, we should expect to find classes of bioactive compounds that occur exclusively, or in significantly greater amounts, in exotic plants.
Materials and Methods
Study Area
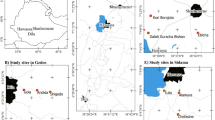
The present study was carried out in the rural community of Carão, which is situated in the municipality of Altinho, Pernambuco State, Brazil (8º 29′ 32′′ S; 36º 03′ 03′′ W) (Fig. 1). The municipality covers 45,449 km2 in the region of Pernambuco state in northeastern Brazil. According to data gathered from the weather station situated in the same municipality, the local annual precipitation is 622 mm, which is characteristic of a semi-arid climate; the months of June and July see the most rain. The average annual temperature is 23.1°C. Altinho is located 168 km from the state capital (Recife) in the physiographic subzone of the agreste in the “Brejo Pernambucano” microregion (IBGE 2007). The last census indicates that the municipality has 21,782 inhabitants, (IBGE 2007). The region is cut by various perennial rivers, but all have only small volume flows; there is little potential for significant ground water supply. The regional vegetation is dry hypoxerophilous arboreal caatinga, with arboreal species that are deciduous or semideciduous. The caatinga occupies a broad region of northeastern Brazil and is represented by different physiognomic types (Araújo et al. 2007). Its most striking feature is climatic seasonality, which completely changes the landscape at certain times of the year. In all physiognomic types of caatinga, the composition of the herbaceous layer is rich. The herbaceous layer is ephemeral, however, changing its composition and richness in time and space.

Study area: the Community of Carão, Altinho, Pernambuco (northeastern Brazil).
The community of Carão is located 16 km from the municipal seat, near the base of the Letreiro mountain range, although some residences are slightly distant from the village center. The community is primarily rural with many small-plot farmers. The principal economic activity is subsistence agriculture, principally monocultures of corn, beans, and manioc that can be planted near the residences and/or in cleared areas on the flat top of the nearby range (known as “chã da serra”). These small plots (“roçados”) represent the principal food supply for these families; excess production is sold in the open market in the town of Altinho. In addition to subsistence agriculture, cattle and goat husbandry are also important to these rural populations. The population is composed in large part of Catholics, with some evangelic Protestant sects. Health services consist of a small community health center that functions once a week with a nurse as well as monthly visits from a doctor. There is a grammar school in the village (Fundamental I), but students have to travel by bus to Altinho for continued studies (Fundamental II and High School); the youth of the community are largely literate (100% between 17 and 26 years) and have between 10 and 13 years of formal education; the large majority of elderly people in the community are illiterate or semi-illiterate (100% between 67 and 90 years).
The regional roads are unpaved, and the houses are made of bricks. While all of the houses have electricity, there is no running water or sewage system; water is collected from cisterns that catch rain water or is brought in barrels from the local rivers.
Data Collection
Before beginning the ethnobotanical survey, the legal representatives of the municipality were contacted and informed of the objectives of the project; a formal presentation of the project goals was also given to Family Health Program (PSF) agents and representatives of the local community. A meeting was then held with a majority of the adult inhabitants (above 18 years) from the village (mediated by the Rural Workers Association of the community), during which we presented the intentions of the research project. Everyone participating in the project was asked to sign a Free Consent and Understanding Agreement (TCLE) that would allow the collection and publication of data and other information, in accordance with the recommendations of the National Health Council (Resolution N° 196/96 CNS). The people who did not attend the public meeting were subsequently contacted with the help of the public health agents, and the same consent and information procedures were followed. A total of 101 people were interviewed (36 men and 65 women between the ages of 19 and 83); women accounted for 90.2% of the population older than 18.
Field work was undertaken between August 2006 and July 2007, with regular monthly visits lasting five days. Semistructured interviews (Albuquerque et al., 2008) were used to collect data about the medicinal plants used by and/or known to the local population for treating health problems, as well as for collecting complementary information about the parts of the plants that are used, the source of the material, how it is prepared, and the quantities that are used in medicinal preparations. The local therapeutic categories were generated based on information supplied by the informants (without any attempt on our part to reclassify them) in order to maintain the internal logic of the local classification system and to preserve its nosology (Albuquerque and Oliveira 2007). At the end of each interview session, a guided-tour was undertaken with the interviewee in order to collect the plants cited during the interview. During the guided tours we collected the plants that were cited in the interviews; however, plants that were not located adjacent to residences of the respondents, such as those in native vegetation areas, were collected during a separate visit.
In a second round of interviews with the informants (in a procedure analogous to that used by Medeiros et al. 2008), the plants that were cited by those same people were read back to them in an attempt to discover more details. In order to determine the local taxonomy of the useful plants and minimize any overestimation of the species richness, examples of fresh plant material were shown to members of the community, who were then asked to supply the local names (in a procedure analogous to the check list–interview) (Albuquerque et al., 2008).
After identifying the species cited during the interviews (261 ethnospecies corresponding to 199 identified species), we randomly selected plants for the phytochemical analyses. The selection was limited to those plants that were cited by three or more informants (98 species in total) and those that were seasonally available during the sampling period. We selected 61 of the 98 species for phytochemical analysis. We intended to examine as many plants as possible; however, because the climate of the region (the typical seasonality of the Caatinga), it was often not possible to find available material in sufficient quantity to be incorporated into the work.
The species cited during the interviews were classified according to their biogeographical distribution and according to whether they were native or exotic. Native plants were considered to be those that occur spontaneously in South America; exotics were considered to be those of extra-continental origin. Additionally, the plants were classified according to their habit (herbs, shrubs, or trees). Trees were considered to be plants with woody habits and a simple trunk, while herbs were plants without any lignification of the stem and aerial parts (Begon et al. 1988).
The species cited during the interviews were collected for botanical identification during the months of June and July of 2007 according to the usual techniques recommended by Mori et al. (1989). All primary specimens were incorporated into the collection at the Professor Geraldo Mariz Herbarium (UFP) of the Universidade Federal de Pernambuco (UFPE); duplicates were donated to the Vasconcelos Sobrinho Herbárium (PEUFR) of the Universidade Federal Rural do Pernambuco (UFRPE).
Phytochemical Analyses
Phytochemical analyses were performed on each of the 61 selected species. This data was important for testing the hypothesis that exotic plants have types of compounds that are not found in native plants (the diversification hypothesis). Analyses were performed on the part of the plant indicated during the interviews as being used for medicinal purposes (bark, leaves, fruits, or latex). For those species that had more than one part cited, material was collected from the plant organ that was mentioned the most. Plant material was collected from at least three adult individuals that showed no signs of predation or damage. Material was collected from plants harvested at the start of the rainy season (July and August 2007), as this was the time period during which plant material was available for both native and exotic plants; in the dry season, herbaceous and subshrub plants are not available due to the Caatinga’s seasonality and most of woody plants completely lose the leaves.
The collected material was dried at room temperature and subsequently ground up in preparation for methanol extraction (1 g/10 ml), according to the technique described by Matos (1997). The following classes of compounds were assayed: phenols, tannins, terpenes (mono-, di-, tri-, and sesquiterpenes), alkaloids (derived from yoimbine and caffeine), quinines (anthraquinones and naphthoquinones), and flavonoids (derived from rutin and quercetin). The analyses were performed using thin layer horizontal chromatography (CCD) with Merck silica gel 60 plates (0.2 mm) and F254 as the fluorescent indicator; solvents and indicator reagents specific for each class of compounds were employed as described by Wagner and Bladt (1996) and Harbone (1998). This chromatographic method was chosen as it provides reasonably rapid results and repeatability and can qualitatively detect the compounds chosen for evaluation.
Only the presence or absence of each chemical class was analyzed for each species; the intensity of the stain on the chromatogram was not considered.
Data Analysis
Williams’ G test (Sokal and Rholf 1995) was used to determine the percentage of species demonstrating positive results in terms of their habit (herb, shrub, or tree) and origin biogeographical (native or exotic) for each class of compounds evaluated (tannins, phenols, flavonoids, terpenes, triterpenes, quinines, and alkaloids).
The relative importance (RI) of the species was calculated according to Bennett and Prance (2000), where RI = NCS + NP; NCS is the number of corporeal systems treated by a given species (NSCS) divided by the total number of corporeal systems treated of the most versatile species (NSCSV). NP represents the number of therapeutic properties attributed to a species (NPS) divided by the number of therapeutic properties attributed to the most versatile species (NPSV). We considered the most important species to be the most versatile. These indices were important for testing the idea proposed by Bennett and Prance (2000) concerning the versatility of exotic plants as compared to native plants.
In order to evaluate the diversity of secondary metabolites present in each plant, a Chemical Diversity Index (CDI) was formulated that was calculated as follows: \( CDI = NOP/NCA \), where
- NOP:
-
number positive occurrences of each compound
- NCA:
-
total number of compounds evaluated.
For example: Myracrodruon urundeuva Allemão was analyzed for eight classes of compounds and gave positive results for phenols, tannins, flavonoids, terpenes, triterpenes, and anthraquinones. Thus, it had six positive occurrences, so its CDI would be six divided by eight (the total number of compound classes evaluated) or 0.75.
It is assumed that a species that has a higher diversity of secondary metabolites has a greater possibility to function in a wide variety of therapeutic settings. This index was used to compare the use-versatility of medicinal plants (measured by their RI) with their diversity of secondary compounds using Spearman’s correlation coefficient.
In order to evaluate the use-versatility of the medicinal plants as compared to their overall versatility, two sums were performed: all of the therapeutic indications of all of the species were summed and all of the use-citations within all of the various use categories of the plants (construction, biofuels, food, etc.). Subsequently, we evaluated the relationships of versatility of therapeutic and general uses among native and exotic plants using the Kruskal Wallis test (Sokal and Rholf 1995). All of the analyses were performed using the BioEstat 2.0 software package (Ayres et al. 2000).
Results
Of the 61 species chosen for analysis, 36 were native and 25 were exotic. Of the native plants, 52.78% were trees, 19.44% were shrubs, and 27.78% were herbs; that is, woody plants made up 72.22% of the sample. Among the exotic plants, 20.0% were trees, 8.0% were shrubs, and 72.0% were herbs, with woody plants therefore making up only 28.0% of the sample. The native species were distributed among 16 families (Table 1); the most species-rich families were: Caesalpiniaceae (5 species), Euphorbiaceae (5), Anacardiaceae (3), and Mimosaceae (3). The exotic species were distributed among 19 families, with the most species-rich families being: Lamiaceae (6), Apiaceae (2), Myrtaceae (2), and Rutaceae (2).
Compound Classes vs. Biogeographical Origin (“Diversification Hypothesis”)
In classifying the plants according to their biogeographical origin, native plants were observed to have more frequent occurrences of alkaloids (G=19.88; p < 0.0001). Exotic plants had more positive occurrences of terpenes (G = 9.03; p < 0.01) than native plants. No significant differences were observed between the two plant groups in the presence of other compounds assayed (Table 2).
Woody exotic plants had more flavonoids and triterpenes (G = 6.30; p < 0.05), but no other significant differences from the other plant classes were observed with the other classes of compounds (Table 2). Exotic herbs presented larger occurrence of phenols (G=10.07; p < 0.05) and tannins (G=12.53; p < 0.001; native herbs had more triterpenes (G = 48.25; p < 0.0001) and alkaloids (G=24.28; p < 0.0001) (Table 2). Flavonoids, terpenes, naphthoquinones, and anthraquinones were equally present in all plant biogeographical origins.
Relative Importance vs. Chemical Diversity Index (“Versatility”)
The plants with the greatest versatility, as measured by the Relative Importance (RI) values, were native species; the most important in this class were Caesalpinia ferrea Mart. (RI=1.81), Myracrodruon urundeuva Allemão (1.75), and Mimosa tenuiflora (Willd.) Poir. (1.50). Of the 10 species with the highest RI values, nine were woody, demonstrating once again that the use-versatility of arboreal and shrub plants is well known in the community studied. The exotic species with the highest RI values were Alpinia speciosa (Blume) D. Dietr. (1.19), Punica granatum L. (1.19), and Chenopodium ambrosioides L. (1.06) (Table 1).
There were no significant relationships between the RI and CDI values of the plants (rs = 0.10; p = 0.42), indicating that the diversity of bioactive compounds in any given species is not related to its versatility, as measured by its relative importance as a medicinal plant.
The arboreal-shrub species demonstrated medium-level values for the parameters measured, having an average RI = 0.954 (Standard Error = 0.39) and CDI = 0.60 (SE = 0.17); herbs had average values of RI = 0.58 (SE = 0.29) and CDI = 0.54 (SE = 0.21); these differences are statistically significant only for RI (H = 14.17; p < 0.001).
The native plants gave average values of RI = 0.85 (SE = 0.44) and CDI = 0.58 (SE = 0.18), while exotic plants gave values of RI = 0.68 (SE = 0.29) and CDI = 0.56 (SE = 0.20); none of these differences were statistically significant.
Native arboreal-shrub plants had an average RI = 1.03 (SE = 0.38) and CDI = 0.59 (SE = 0.17), while the average arboreal-shrub exotic plant values were RI = 0.66 (SE = 0.30) and CDI = 0.64 (SE = 0.20); these differences were significant only for RI (H = 3.93; p < 0.05). The exotic herbs had average values RI = 0.69 (SE = 0.29) and CDI = 0.53 (SE = 0.20), which were larger than the native herb values: RI =0.38 (SE = 0.19) and CDI = 0.56 (SE = 0.22). As such, exotic herbs demonstrated significantly greater versatility (H = 6.17; p < 0.05).
Native plants demonstrated a positive association between the number of medicinal properties of the plant and its versatility for other uses (rs = 0.61; p < 0.0001); a similar association was not observed for exotic plants. Native plants, as compared to exotic species, had more general use–citations (H = 5.27; p < 0.05). Only the native species had a positive relationship between versatility as food sources or ornamentals and medicinal uses (rs = 0.48; p < 0.01).
Discussion
Compound Classes vs. Biogeographical Origin (“Diversification Hypothesis”)
Our data indicate that the diversification hypothesis (Albuquerque 2006) best explains the inclusion of exotic plants into traditional pharmacopoeias in the Caatinga region. The diversification hypothesis suggests that traditional communities incorporate exotic medicinal plants into their medical repertoire in order to diversify the local medicinal stock. According to this hypothesis, we would expect that exotic plants contain large amounts of secondary compounds or types of secondary compounds that are distinct from those in native plants.
This hypothesis is consistent with our results: exotic plants, including trees and herbs, were found to contain large amount of flavonoids and terpenes, while the native group had higher amounts of triterpenes and alkaloids. Considering the plants in terms of their habit, exotic trees were found to contain more terpenes, triterpenes, anthraquinones, and flavonoids than native trees, while exotic herbs were positive for terpenes and anthraquinones. These results seem to indicate that exotic species are incorporated into the local pharmacopoeia in order to diversify therapeutic options and not to create competition with the native species (see Albuquerque 2006; Albuquerque and Oliveira 2007).
Several factors are associated with a decline in traditional botanical knowledge that leads to an “erosion” of knowledge. Globalization and the destructive exploitation of tropical forests are two examples. The first sign of an impending decline is often a growing disinterest in the traditional lifestyle of the community among the young members of the society (Begossi et al., 2002; Caniago and Siebert 1998; Estomba et al. 2006; Hanazaki et al. 2000; Phillips and Gentry 1993; Ugent 2000). This attitude puts the accumulated local knowledge at risk of being lost without continued transmission through the generations (Galeano 2000). Other authors believe that loss of knowledge in traditional communities occurs when two or more different groups enter into close contact, producing changes in both communities in a process known as acculturation (Foster 1974; Zent 2001). There are researchers who likewise see the expressive occurrence of exotic plants in traditional pharmacopoeias as the result of erosion or acculturation (Palmer 2004)
It is important to emphasize that the erosion of knowledge is considered to be a product of the acculturation of traditional communities, but there are also other products. The loss of knowledge of medicinal plants may be due in part to the fact that the young people now spend a good deal of their time at school or playing in the streets and much less time in the forest (Zent 2001). The degree of this loss will probably only be seen further in the future, as the current generation is only experiencing the first wave of globalization and still retains a large part of their older knowledge.
In order to make a more secure evaluation of the processes of acculturation, a diachronic approach must be taken, comparing data from the past with more recent research, although our results indicate that a local pharmacopoeia can incorporate new elements without losing knowledge. As such, an exotic plant inserted into a local pharmacopoeia does not necessarily supplant a native species but may simply be filling a therapeutic vacancy for which there were no equivalent (or as effective) native plants.
Relative Importance vs. Chemical Diversity Index (“Versatility”)
Bennett and Prance (2000) observed that exotic plants can be adopted into a culture due to their high versatility for general usage (for example, as an ornamental and/or food source); they would then be candidates for incorporation into the traditional knowledge of the culture and into the local pharmacopoeia. This idea may be equally valid in the Caatinga region, as exotic plants usually have other nonmedicinal uses (especially as food sources and ornamentals). The results of the present work do not corroborate the idea of versatility, however, as the native plants were more versatile in terms of their general uses and significant differences were not observed in the medical versatility of native versus exotic species. A majority of the native plants of the caatinga are in fact known for their multiple uses (Lucena et al. 2007a, b).
Chemical diversity, as measured by the CDI, could not explain the incorporation of exotic plants into the local pharmacopoeia: the results for exotic plants and native plants were not significantly different, and there was not an ample spectrum of secondary compounds that directly influenced the incorporation of these plants into the local pharmacopoeia. Estomba et al. (2006) observed that most exotic plants are used to treat gastrointestinal problems; this result indicates that exotic plants are chosen to occupy specific positions in the pharmacopoeia and they are not included in the local folk medicinal repertoire because they have particularly wide spectrums of use. Albuquerque and Oliveira (2007) observed that most exotic plants have therapeutic indications for ailments not covered by native plants; this observation supports our overall interpretation.
We conclude that the incorporation of exotic plants into traditional pharmacopoeias may not reflect acculturation events or use-versatility considerations. In the community studied, we found a scenario in which exotic plants appear to be incorporated due to the fact that they contain classes of secondary bioactive compounds that are absent from or present in small amounts in native plants. Exotic plants are thus selected for use in specific therapeutic treatments, while native plants are used in a more diversified manner.
Literature cited
Albuquerque, U. P. 2006. Re-examining Hypotheses Concerning the Use and Knowledge of Medicinal Plants: A Study in the Caatinga Vegetation of NE Brazil. Journal of Ethnobiology and Ethnomedicine 2:30.
———, L. H. C. Andrade, and A. C. O. Silva. 2005. Use of Plant Resources in a Seasonal Dry Forest (Northeastern Brazil). Acta Botanica Brasilica 19(1):27–38.
——— and R. F. Oliveira. 2007. Is the Use-Impact on Native Caatinga Species in Brazil Reduced by the High Species Richness of Medicinal Plants? Journal of Ethnopharmacology 113(1):156–170.
——— and R. F. P. Lucena. 2005. Can Apparency Affect the Use of Plants by Local People in Tropical Forests? Interciencia 30(8):506–510.
———, ———, and N. L. Alencar. 2008. Métodos e Técnicas na coleta de dados etnobotânicos. in U. P. Albuquerque, R. F. P. Lucena, and L. V. F. C. da Cunha, eds., Métodos e Técnicas na Pesquisa Etnobotânica, 2nd edition. NUPEEA/Comunigraf, Recife, Brazil.
Alencar, N. L., T. A. S. Araújo, E. L. C. Amorim, and U. P. Albuquerque. 2009. Can the Apparency Hypothesis Explain the Selection of Medicinal Plants in an Area of Coatinga Vegetation? A Chemical Perspective. Acta Botanica Brasílica 23:910–911.
Almeida, C. F. C. B. R., T. C. Lima e Silva, E. L. C. Amorim, M. B. S. Maia, and U. P. Albuquerque. 2005. Life Strategy and Chemical Composition as Predictors of the Selection of Medicinal Plants from the Caatinga (Northeast Brazil). Journal of Arid Environments 62(1):127–142.
Araújo, E. L., C. C. Castro, and U. P. Albuquerque. 2007. Dynamics of Brazilian Caatinga: A Review Concerning the Plants, Environment and People. Functional Ecosystems and Communities 1(1):15–28.
Araújo, T. A. S., N. L. Alencar, E. L. C. Amorim, and U. P. Albuquerque. 2008. A New Approach to Study Medicinal Plants with Tannins and Flavonoids Contents from the Local Knowledge. Journal of Ethnopharmacology 120:72–80.
Ayres, M., M. J. Ayres, D. L. Ayres, and S. A. Santos. 2000. BioEstat 2.0: Aplicações estatísticas nas áreas das ciências biológicas e médicas. Sociedade Civil Mamirauá, CNPq, Brasília, Brazil.
Begon, M., J. L. Harper, and C. R. Townsend. 1988. Ecologia: Individuos. Poblaciones y Comunidades, Ediciones Omega, Barcelona.
Begossi, A., N. Hanazaki, and J. Tamashiro. 2002. Medicinal Plants in the Atlantic Forest (Brazil): Knowledge, Use, and Conservation. Human Ecology 30:281–99.
Bennett, B. C. and G. T. Prance. 2000. Introduced Plants in the Indigenous Pharmacopeia of Northern South America. Economic Botany 54(1):90–102.
Benz, B. F., J. E. Cevallos, F. M. Santana, J. A. Rosales, and S. M. Graf. 2000. Losing Knowledge about Plant Use in the Sierra de Manatlán Biosphere Reserve, Mexico. Economic Botany 54(2):183–191.
Caniago, I. and S. F. Siebert. 1998. Medicinal Plant Ecology, Knowledge and Conservation in Kalimantan, Indonesia. Economic Botany 52:229–50.
Estomba, D., A. Ladio, and M. Lozada. 2006. Medicinal Wild Plant Knowledge and Gathering Patterns in a Mapuche Community from North-western Patagonia. Journal of Ethnopharmacology 103(1):109–119.
Foster, G. M. 1974. Antropología aplicada. Fondo de Cultura Económica, México D.F.
Galeano, G. 2000. Forest Use at the Pacific Coast of Chocó, Colombia: A Quantitative Approach. Economic Botany 54:358–376.
Hanazaki, N., J. Y. Tamashiro, H. F. Leitão-Filho, and A. Begossi. 2000. Diversity of Plant Uses in Two Caiçara Communities from the Atlantic Forest Coast, Brazil. Biodiversity and Conservation 9:597–615.
Harbone, J. B. 1998. Phytochemical Methods. Kluwer Academic Publishers, Dordrecht.
IBGE—Instituto Brasileiro de Geografia e Estatística. http://www.ibge.gov.br/ (2007)
Janni, K. D. and J. W. Bastien. 2004. Exotic Botanicals in the Kallawaya Pharmacopoeia. Economic Botany 58 (Supplement):S274–S279
Lucena, R. F. P., E. L. Araújo, and U. P. Albuquerque. 2007a. Does the Local Availability of Woody Caatinga Plants (Northeastern Brazil) Explain Their Use Value? Economic Botany 61:347–361.
———, U. P. Albuquerque, J. M. Monteiro, C. F. C. B. R. Almeida, A. T. N. Florentino, and J. S. F. Ferraz. 2007b. Useful Plants of the Semi-arid Northeastern Region of Brazil—A Look at their Conservation and Sustainable Use. Environmental Monitoring and Assessment 125(1):281–290.
Matos, F. J. A. 1997. Introdução à Fitoquímica Experimental, 2nd edition. Editora UFC, Fortaleza, Ceará, Brazil.
Medeiros, P. M., A. L. S. Almeida, R. F. P. Lucena, and U. P. Albuquerque. 2008. Uso de estímulos visuais na pesquisa etnobotânica. in U. P. Albuquerque, R. F. P. Lucena, and L. V. F. C. da Cunha, eds., Métodos e Técnicas na Pesquisa Etnobotânica, 2nd edition. NUPEEA/Comunigraf, Recife, Brazil.
Mori, A. S., L. A. M. Silva, and G. Lisboa. 1989. Manual de manejo do herbário fanerogâmico, 2nd edition. Centro de Pesquisa do Cacau, Ilhéus, Bahia, Brazil.
Müller-Schwarze, N. K. 2006. Antes and Hoy Día: Plant Knowledge and Categorization as Adaptations to Life in Panama in the Twenty-First Century. Economic Botany 60(4):321–334.
Palmer, C. 2004. The Inclusion of Recently Introduced Plants in the Hawaiian Ethnopharmacopoeia. Economic Botany 58 (Supplement):S280–S293.
Phillips, O. and A. H. Gentry. 1993. The Useful Plants of Tambopata. Peru: II. Additional Hypothesis Testing in Quantitative Ethnobotany. Economic Botany 47:33–43.
Quinlan, M. B. and R. J. Quinlan. 2007. Modernization and Medicinal Plant Knowledge in a Caribbean Horticultural Village. Medical Anthropology Quarterly 21(2):169–192.
Reyes-García, V., E. Byron, R. Godoy, V. Vadez, L. Apaza, E. Pérez, W. Leonard, and D. Wilkie. 2004. Measuring Culture as Shared Knowledge: Do Data Collection Formats Matters? Cultural Knowledge of Plant Uses among the Tsimane’ Amerindians of Bolivia. Field Methods 16:135–156.
Sokal, R. R. and F. G. Rholf. 1995. Biometry. Freeman and Company, New York.
Stepp, J. R. 2004. The Role of Weeds as Source of Pharmaceuticals. Journal of Ethnopharmacology 92(1):163–166.
——— and D. E. Moerman. 2001. The Importance of Weeds in Ethnopharmacology. Journal of Ethnopharmacology 75(1):25–31.
Ugent, D. 2000. Medicine, Myths and Magic: The Folk Healers of a Mexican Market. Economic Botany 54:27–438.
Voeks, R. A. 1996. Tropical Forest Healers and Habitat Preference. Economic Botany 50(4):381–400.
——— 2004. Disturbance Pharmacopoeias: Medicine and Myth from the Humid Tropics. Annals of the Association of American Geographers 94(4):868–888.
——— and A. Leony. 2004. Forgetting the Forest: Assessing a Medicinal Plant Erosion in Eastern Brazil. Economic Botany 58 (Supplement):S294–S306.
Wagner, H. and S. Bladt. 1996. Plant Drug Analysis—A Thin Layer Chromatography Atlas, 2nd edition. Springer, Berlin.
Zarger, R. K. and J. R. Stepp. 2004. Persistence of Botanical Knowledge among Tzeltal Maya Children. Current Anthropology 45(3):413–418.
Zent, S. 2001. Acculturation and Ethnobotanical Knowledge Loss among the Piaroa of Venezuela. In L. Maffi, On Biocultural Diversity—Linking, Knowledge, and the Environment. Smithsonian Institution Press, Washington, D.C.
Acknowledgments
The authors would like to thank the community of Carão for their gracious welcome and receptivity, as well as the following researchers from the Laboratory of Applied Ethnobotany (LEA) for their support during the ethnobotanical surveys: Ernani Machado de Freitas Lins Neto, Flávia Santos Almeida, Joabe Gomes de Melo, Alyson Luiz de Almeida, Miguel de Santana Almeida, Luciana Gomes de Sousa, Viviany Teixeira do Nascimento, and Lucilene Lima dos Santos. We also thank Victoria Lacerda of the Universidade Federal de Santa Catarina for help during her internship at the LEA; the Municipality of Altinho, especially the Secretary of Agriculture, Sr. Miguel Andrade Júnior, for logistic support; the Community Health Agents’ Carão, Srs. Inaldo, and Alexandre; and the CNPq for their financial support (“Edital Universal”) and for the productivity grant to U.P. Albuquerque.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article
Cite this article
Leal Alencar, N., de Sousa Araújo, T.A., de Amorim, E.L.C. et al. The Inclusion and Selection of Medicinal Plants in Traditional Pharmacopoeias—Evidence in Support of the Diversification Hypothesis. Econ Bot 64, 68–79 (2010). https://doi.org/10.1007/s12231-009-9104-5
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s12231-009-9104-5