
Abstract
Traditional Management and Morphological Patterns of Myrtillocactus schenckii (Cactaceae) in the Tehuacán Valley, Central Mexico Myrtillocactus schenckii is a columnar cactus endemic to central México and appreciated for its edible fruit. In the Tehuacán Valley it occurs wild in thorn-scrub forests, but it is also under silviculture management—tolerate in agroforestry systems and cultivated in home gardens. Ethnobotanical and morphometric studies were conducted to document its use and management forms, as well as consequences of artificial selection on morphological patterns in managed populations. In silviculture populations artificial selection occurs through selective tolerance of the phenotypes producing more, larger, and sweeter fruits, whereas in home gardens people plant branches of the preferred phenotypes. Morphological differences were found especially in fruit size and production, the main targets of artificial selection. The highest average values of fruit size and production were found in cultivated populations (0.993 cm3 and 204.45 fruits per branch, respectively), intermediate in silviculture populations (0.819 cm3 and 70.63 fruits per branch), and the lowest in the wild (0.68 cm3 and 59.75 fruits per branch). Multivariate statistical analyses differentiated populations according to their management type. Morphological diversity was higher in managed populations (0.703 ± 0.029 and 0.679 ± 0.019 in silviculture and cultivated populations, respectively) than in the wild (0.652 ± 0.016). Managed plants of M. schenckii do not show signs of depending on humans for survival and reproduction and, therefore, domestication should be considered incipient.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
The Tehuacán Valley is one of the areas of Mesoamerica where the history of plant management has been better documented. Archaeological studies by MacNeish (1967) reported information on plants used and managed from prehistory, including some of the oldest remains of agriculture in the New World. In addition, ethnobotanical studies (Casas et al. 2001; Lira et al. 2009) have documented a broad spectrum of forms of interactions between people and plants currently occurring in the region, among them a variety of agricultural and silviculture practices such as tolerance, promotion, special care, transplanting, and sowing of nearly 120 native plant species (Casas et al. 2008). Management of all these species is conducted with different intensities according to the human energy invested, tools, productivity, and practice of artificial selection, but how intensive the plant management can be is a question still analyzed by several authors (Casas et al. 1999a, 2007; González-Insuasti and Caballero 2007). Columnar cacti have been interesting study systems to analyze this research question, and studies available on these plants offer the possibility of documenting how the intensity of artificial selection may vary within and among related species (Casas et al. 1999a). Also, it is possible to analyze how the intensity of artificial selection, life history aspects, and interactions among the coexisting wild and managed plant populations may influence the degree of domestication (Casas et al. 2007).
Columnar cacti produce edible fruits that are generally gathered from wild populations, but some are also in situ or silviculture managed in association with agroforestry systems (Casas et al. 1999a). Silviculture is directed to maintain or increase availability of useful plant species or particularly good phenotypes of some species (with larger and sweeter fruits, and with thinner and fewer spines in the fruit peel), and this management form has been interpreted as artificial selection (Casas et al. 1999b). Some species of columnar cacti are also cultivated in home gardens, where artificial selection is conducted more intensely by planting both sexual and asexual propagules of the desirable phenotypes, and continually replacing plant materials in the cultivated areas (Casas et al. 2006, 2007).
Several species of columnar cacti have been studied from ethnobotanical, morphometric, reproductive, and population genetics approaches in order to document how artificial selection is conducted and its impacts on plant populations (Casas et al. 2007). The morphometric studies of Stenocereus stellatus (Casas et al. 1999b), Polaskia chende (Cruz and Casas 2002), P. chichipe, and Escontria chiotilla (Casas et al. 2007) suggest that morphological divergence between wild and managed populations within and between species is influenced by the intensity of artificial selection as well as characteristics of the plant species determining the degree of reproductive isolation between wild and managed populations (Casas et al. 2007). Within species, wild and cultivated populations are generally the most divergent, whereas the silviculture populations are intermediate. Among species, morphological divergence between wild and managed populations is higher in species under higher artificial selection intensity (Casas et al. 2007). But the degree of reproductive isolation between wild and managed populations may also influence the degree of differentiation (Otero-Arnaiz et al. 2005; Parra et al. 2008; Tinoco et al. 2005). Gene flow among populations occurs through movement of pollen, seeds, and asexual propagules, but it strongly depends on the establishing success of propagules. Pollen movement is determined by bats in species such as Stenocereus spp. (Casas et al. 2007), whereas bees are the most important pollinators of Escontria chiotilla (Oaxaca-Villa et al. 2006), Polaskia spp. (Cruz and Casas 2002; Otero-Arnaiz et al. 2003), and Myrtillocactus spp. (Ortíz et al. n.d.). Seeds of bat-pollinated species have higher water requirements to germinate and survive (Guillén et al. 2009), but vegetative propagation is more successful with faster growth than branches of bee-pollinated species. Bee-pollinated species have more limited pollen mobility but their seeds have lower water requirements (Guillén et al. 2009), thus establishing their seedlings more successfully; however, these species show limited capacity for vegetative propagation and grow slower than bat-pollinated species. But these capacities are variable among species; for instance, Polaskia chichipe has good capacity of seedling establishment and intermediate capacity of vegetative propagation and growth velocity. Differentiation between wild and managed populations has been found to be higher in S. stellatus compared with Escontria chiotilla, species representing the highest and lowest management intensity levels, respectively (Casas et al. 2007).
Management and domestication of Myrtillocactus schenckii has not been analyzed, but information already available indicates that this species is under relatively intermediate management intensity since, although it can be found under cultivation, it is scarcer than other species in home gardens (Casas et al. 1999a), and reproductive biology studies (Ortíz et al. in press) indicate that vegetative propagation is possible but only partially successful. M. schenckii is endemic to semi-arid regions of Puebla and Oaxaca, central Mexico, growing wild in tropical deciduous and thorn-scrub forests in areas of soils derived from volcanic rocks at elevations of 1,300 to 1,900 m. It is 3 to 5 m in height, with dark green branches. Flowers are small (an open corolla being nearly 2.5 cm diam), white, with diurnal anthesis. Blooming occurs from November to April, flower production being much higher in cultivated populations, where self-pollination also is more frequent (Ortíz et al. in press). Plabeia mexicana (stingless bee), Apis mellifera, Tabanus sp., and Xylocopa mexicanorum (carpenter bee) are the main flower visitors (Ortíz et al. in press). Fruits are spherical or ellipsoid, 0.8 to 1.5 cm in length and 0.6 to 1.5 cm in width. Fruiting occurs from May to July (Bravo-Hollis 1978). According to Ortíz et al. (in press), human management of M. schenckii influences differences in phenology, frequency of flower visitors, and breeding systems in populations, which may limit pollen flow between wild and managed populations.
In the Tehuacán Valley M. schenckii is called “garambullo” and is appreciated for its edible fruit. It occurs wild in the cactus forest called “garambullal,” where it is a dominant species and where people gather its fruit. But it also occurs in agroforestry systems, where people tolerate juvenile and adult plants, and in home gardens, where it is a cultivated plant. We expected to find higher differentiation in fruit features between wild and cultivated populations and intermediate differentiation between silviculture populations in agroforestry systems and the other populations. The information available also allows us to hypothesize that M. schenckii, similarly to P. chichipe, exhibits an intermediate degree of domestication. The purpose of this study was therefore to document how M. schenckii is used and managed, how artificial selection is conducted, and what are its effects on phenotype frequencies in populations.
Methods
Study Area
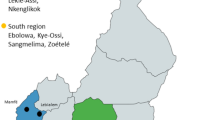
The study was conducted in the territory of the villages of San Luis Atolotitlán and Santiago Coatepec, in the state of Puebla, Mexico (Fig. 1), within the Tehuacán-Cuicatlán biosphere reserve. Climate of the area is semi-arid, with annual mean temperature of 18.5°C, and annual mean precipitation of 400 mm. Populations of M. schenckii were sampled in elevations from 1,800 to 2,000 m, and included three wild, three silviculture managed, and two cultivated populations. People of the villages are of Náhuatl origin, whose subsistence depends on rain-fed maize agriculture, raising goats, and extraction of forest products. Commercialization of products such as agave mescal, palm handcrafts, and fruits from their home gardens complement their economy.

Geographic location of the study area in the Tehuacan Valley. Populations studied in the municipalities of San Luis Atolotitlán and Santiago Coatepec, Puebla, Mexico.
Plant Populations Studied
Wild populations occur in natural vegetation called garambullal where M. schenckii is a dominant species along with trees and shrubs such as Polaskia chichipe, P. chende, Pittocaulon praecox, Agave salmiana, Mimosa luisana, Acacia cochlyacantha, Ipomoea arborescens, and Opuntia pillifera. Silviculture populations represent M. schenckii established in agroforestry systems derived from natural garambullal that have been under recurrent cycles of use and fallow for centuries. Polaskia spp., O. pillifera, I. arborescens, and A. salmiana were tolerated when the fields were cleared for cultivation. Cultivated populations are those within home gardens in the villages of San Luis Atolotitlán and Santiago Coatepec. Home gardens in these villages are approximately 200 m2 and frequently contain as many as 60 species of edible, medicinal, and ornamental plant species. Other columnar cacti are cultivated in home gardens, the most common being S. pruinosus (11 individual plants per plot on average) and S. stellatus (7 individual plants per plot on average), followed by P. chichipe (5 individual plants per plot on average) and M. schenckii (3 individual plants per plot on average). Samples of 30 adult individual plants selected randomly were studied per population (240 individual plants in total). In wild populations we established 1,000 m2 squares where reproductive individuals were sampled at random. We completed the sample of 30 individual plants in a total of 7 and 6 plots of agroforestry systems in San Luis Atolotitlán and Coatepec, respectively. For sampling home gardens, we assigned a number to each house in a village and selected the sample through random numbers. In the Tehuacán Valley, M. schenckii has a distribution associated with localized and scarce patches of soils derived from volcanic rocks, and the villages studied are representative of the environments (and populations) where M. schenckii is found in the region.
Ethnobotanical and Morphometric Studies
Information on use and management of garambullo was documented through ethnobotanical semistructured interviews and a survey to 30 and 35 households (nearly 20% of the total) randomly selected in the villages studied, respectively. Plant parts used, perception of morphological variation, the features preferred, and management techniques that favor survival and reproduction of the preferred phenotypes were investigated. Data were analyzed through descriptive statistical methods.
Morphological variation of 26 characters (10 of them vegetative and 16 reproductive) was analyzed (Table 1). Three to five structures were measured and averaged per individual plant. Flowers were collected and preserved in 70% ethanol, whereas fruits were collected and maintained in plastic bags, refrigerated, and then measured in the laboratory. Fruit size was calculated based on measures of two perpendicular diameters and using the formula of spherical bodies volume (v = 4/3 π a2b, where a and b are half of the diameters measured). Fruit sweetness was determined by measuring percentage of sugars in pulp (ºBrix) through a refractometer Atago (0–32 ºBrixs), and acidity was measured in pH units through a pH meter Denver Instrument 215. Measurements of vegetative parts were conducted in the field. General biomass of individual plants was measured using the formula of an inverted truncated cone v = πh/3 (R2 + Rr + r2), in which h is the plant height, R is the canopy diameter, which was estimated as the average of two perpendicular measures (north-south and east-west oriented) of canopy diameter, and r is the diameter of the main stem measured just below the first ramification. Branch diameter was measured in the mid-part of branches at the third level of ramification, where also number, width, and depth of ribs and distance between areoles were measured.
Data Analyses
Principal Component (PCA) and Discriminant Function Analyses (DFA, Sneath and Sokal 1973) were conducted in order to classify individual plants analyzed according to their general morphological similarity, and to explore if their similarity is related to the management regime. PCA was also used to identify those characters with higher contribution (higher eigenvectors scores) to the classification, and DFA was directed to test whether the wild, silviculture, and cultivated groups are significantly different among themselves and to identify the proportion of individual plants of each group resembling plants of different groups. A basic data matrix was constructed with individual plants as operative taxonomic units (OTUs) and morphological characters as variables. Considering the different types of variables and measurement units, the basic data matrix was standardized through the algorithm Y’ = (Y-a)/b, in which Y’ is the standardized value, Y is the real value of that character, a is its average value, and b is its standard deviation. PCA was performed based on a Pearson correlation between characters. These analyses were conducted through NYSYS (Rohlf 1993).
One-way ANOVA and Tukey (95% confidence) multiple-range tests were performed through JMP (JMP-SAS Institute 1996) to identify how morphological features differed among all populations studied and among populations pooled according to their management type. The purpose of these analyses was to visualize trends of variation among populations, but general differences among groups of populations were tested through DFA.
An index of morphological diversity (MD) was calculated based on the Simpson diversity index, in order to summarize information on the amount of variation of all variables in a single value. This index was defined as MD = 1-Σ1-s(pi)2 in which pi is the proportion of the total number of individual plants sampled in a population showing the ith state of morphological character and s is the number of states of that character (Casas et al. 2006). Frequencies of character states were first calculated; since all characters studied are quantitative, a previous conversion to qualitative states was necessary. Such transformation was conducted based on the significance intervals calculated through the ANOVA and Tukey multiple range tests, similar to the method used by Casas et al. (2006) and Blancas et al. (2006). Phenotypic differentiation between pools of wild, silviculture managed, and cultivated populations was analyzed using the algorithm of Nei’s genetic distance (Nei 1972), which, in our study, considered the types and frequencies of morphological character states in populations. The degree of differentiation of M. schenckii was compared with those calculated for Stenocereus stellatus, Polaskia chichipe, and P. chende, based on data by Casas et al. (2007) and Cruz and Casas (2002).
Results
Use, Management, and Artificial Selection
Garambullo is widely used by all people surveyed. Fruits are the main useful part, but dry branches and roots are occasionally used as fuelwood and the entire plant for living fences. Most people (nearly 67%) considered that consumption of garambullo fruits has been similar throughout time, but the remaining people interviewed considered that it was higher in the past, when “people lived using more wild resources and these were more abundant.” This last group said that at present some native resources (including garambullo fruits) have been replaced by other fruits obtained in the market. In the past, they said, the lack of roads limited commerce and people had to be more self-sufficient than at present. People depended more upon agriculture and gathering resources in the wild, whereas at present more people work in small industries in the neighboring villages and migrate to other regions of Mexico or the U.S.A. Nearly 67% of people interviewed considered that garambullo has decreased its abundance because clearing of land for agriculture has increased. Some people (15%) also perceived that rain has decreased and that it is more irregular than in the past.
Most people interviewed (83%) said that garambullo plants are tolerated when land is cleared to establish agricultural fields, whereas the rest said they remove them. The main reasons mentioned to tolerate them were that these plants (1) produce edible fruits (43%), (2) should be maintained according to customary norms (25%), (3) are part of an agricultural practice inherited from their parents (11%), (4) allow shade areas for people and domestic animals (8%), (5) help to prevent soil erosion (6%), (6) are used for stubble storing (5%), and (7) may be living fences to limit the parcels (2%). In contrast, people that remove garambullo plants argued that their presence makes agricultural labors more difficult and occupies space that could be used for maize; they said that because garamabullo is abundant in the area, it is not affected by its removal from agricultural fields.
Approximately 20% of people said they transplant garambullo plants from wild populations into their home gardens. Nearly 50% said that vegetative propagation is difficult or of uncertain success, whereas the others said that vegetative propagation is possible but more difficult than for other columnar cacti such as Stenocereus spp. However, only 10% recognized that individual plants producing more and larger fruits are preferred when making decisions on which plant to tolerate or to transplant.
All people interviewed agreed that garambullo plants occurring in home gardens and agroforestry systems do not receive special care or maintenance, but they indirectly benefit from some practices. For instance, in home gardens where M. schenckii grows in association with the dragon fruit locally called pitahaya (Hylocereus undatus), people observed that garambullo plants are more vigorous because of the occasional irrigation given to pitahayas during the dry season. According to respondents, garambullos occurring in agroforestry systems benefit from agricultural practices such as weeding and tillage.
Patterns of Morphological Variation
According to PCA, the first three principal components explain 93.32% of the variation. Individual plants under different management regimes were found forming a continuous gradient of similarity rather than discrete identifiable groups (Fig. 2). The first principal components classified most of the cultivated individual plants in the upper-right side of the plot and most of the wild and silviculture in situ managed individuals grouped at the left lower area of the plot. Eigenvectors showed that characters with higher contribution to the first principal components are dimensions of fruit (larger in cultivated population) and seed weight (smaller in cultivated individual plants), whereas in the second principal component the number of fruits per branch is the most relevant character (Table 1).

Projection of individuals of Myrtillocactus schenckii in the space of the first and second principal components. Black symbols are wild populations (▲ = San Luis Atolotitlán wild 1; ■ = San Luis Atolotitlán wild 2; ● = Coatepec wild); gray symbols are silviculture managed populations ( = San Luis Atolotitlán silviculture 1;
= San Luis Atolotitlán silviculture 1; = San Luis Atolotitlán silviculture 2;
= San Luis Atolotitlán silviculture 2; = Coatepec silviculture); white symbols are cultivated populations (∆ = San Luis Atolotitlán cultivated; □ = Coatepec cultivated).
= Coatepec silviculture); white symbols are cultivated populations (∆ = San Luis Atolotitlán cultivated; □ = Coatepec cultivated).
According to DFA, the multivariate morphological differences among the three population types were significant (Table 2), most individual plants being classified according to their management type. However, it is noteworthy that some trees from a population are similar to most trees in a population under different management type (Table 3). For example, nearly 27% of wild trees are similar to the silviculture group and only 2 wild trees (nearly 3%) are similar to the cultivated group; approximately 21% trees from silviculture populations are similar to the wild group, but only 6 of them (nearly 7%) are similar to the cultivated group; almost 85% cultivated trees were classified within the cultivated group and the few remaining trees were similar to the wild and silviculture groups. This information indicates that wild and silviculture in situ managed populations are similar among themselves and clearly different from the cultivated stands, as also illustrated in Fig. 3.

Discriminant function analysis. Classification of individuals of Myrtillocactus schenckii according to forms of management. Black symbols are wild populations (▲ = San Luis Atolotitlán wild 1; ■ = San Luis Atolotitlán wild 2; ● = Coatepec wild); gray symbols are silviculture managed populations ( = San Luis Atolotitlán silviculture 1;
= San Luis Atolotitlán silviculture 1; = San Luis Atolotitlán silviculture 2;
= San Luis Atolotitlán silviculture 2; = Coatepec silviculture); white symbols are cultivated populations (∆ = San Luis Atolotitlán cultivated; □ = Coatepec cultivated).
= Coatepec silviculture); white symbols are cultivated populations (∆ = San Luis Atolotitlán cultivated; □ = Coatepec cultivated).
One-way ANOVAs identified significant differences in 14 of the 26 characters analyzed, all of them showing significant differences between cultivated populations and the others. In particular 8 characters (stigma length, pericarpel diameter, rib depth, distance between areoles, fruit volume and weight, seed weight, and number of fruits per branch) showed marked differences according to the management type (Table 1).
The highest values of morphological diversity (MD) were found in the silviculture populations (Table 5), which averaged 0.768 ± 0.016, followed by the cultivated populations (MD = 0.679 ± 0.019) and the wild populations (MD = 0.652 ± 0.016). The highest diversity was found particularly in the population in situ managed 2 (MD = 0.842 ± 0.017) (Table 5) and the cultivated population 1 (MD = 0.764 ± 0.017), whereas the lowest values were found in the wild population 1 (MD = 0.547 ± 0.026).
Discussion
Compared with other columnar cacti species of the Tehuacán Valley, Myrtillocactus schenckii can be considered as a plant resource of relatively intermediate importance since quantities of fruit consumed are higher than those reported for Pachycereus hollianus (Rodríguez-Arévalo et al. 2006) and Myrtillocactus geometrizans, (Casas et al. 1999a, b), but lower than those reported for Stenocereus stellatus, Escontria chiotilla, and Polaskia spp. (Casas et al. 1999a, 2007). In addition, it should be mentioned that M. schenckii is cultivated (higher management intensity than E. chiotilla and P. chende, which are not cultivated), but trees of this species are scarcer than other columnar cacti, as mentioned above. Fruits are the main useful part of garambullo, and both silviculture management and cultivation involve artificial selection mainly directed to favor phenotypes producing larger fruits and more fruits per branch. Most people tolerate garambullo plants in agroforestry systems, some transplant individual plants or branches in both agroforestry systems and home gardens, but only 10% people mentioned practicing selection favoring the most productive plants. The cultural importance and management intensity appear to have decreased in the last decades as access to commercialized exotic fruits and out migration increased.
The multivariate statistical analyses performed showed that individual plants are morphologically similar among themselves according to the management type, which suggests that management influences the morphological patterns reported. The populations studied diverged according to the management intensity, the differences between wild and cultivated populations being higher than those between wild and silviculture populations. The one-way ANOVAs performed consistently found that fruit characters differ among populations according to their management regime (Table 1), suggesting that morphological patterns among populations differ because human management changes the frequencies of phenotypes in populations and, consequently, the average values of the characters analyzed. Fruit size and number per branch are the highest in cultivated populations and the lowest in the wild, a pattern similar to those documented in other species of columnar cacti (Casas et al. 2007). Long-term experiments (because of the long life cycle of M. schenckii) of quantitative genetics are still necessary to test whether the morphological characters analyzed are heritable, but the occurrence of morphological variation within a single site as reported in this study suggests that genes contribute significantly to the phenotypic variation.
Seed weight was the lowest in cultivated populations and the highest in wild populations, a finding which differs from the common pattern reported for other columnar cacti, in which seed number and size are directly proportional to fruit size (Casas et al. 2007). Number of ovules was not significantly different among populations; therefore, the fewer seeds produced by fruits of cultivated populations reveal a lower efficiency of the pollinators in the cultivated population, probably associated with a relative scarcity of pollinators in relation to the high number of flowers that cultivated garambullos produce. But also, as revealed by Ortíz et al. (in press), self-pollination is significantly higher in cultivated populations than in the wild, and this breeding system results in lower production of seeds than out-crossing. Therefore, differences in seed number and size among populations could be associated with frequency of self-pollination occurring in populations under different management regimes. Flower pericarpel was significantly larger in cultivated and silviculture populations than in the wild, and this difference could be related to artificial selection favoring larger fruits. Therefore, fruit size of M. schenckii appears to be related to size of flower parts rather than to seed number.
The populations of M. schenckii studied have the highest levels of morphological diversity compared with other species of columnar cacti (Table 4). As in other species, the highest levels of morphological diversity were found in managed populations, which indicates that human interaction with those populations favor variable stands, a common pattern in plants under domestication (Hawkes 1983), and silviculture and cultivated populations are reservoirs of variation that should be taken into account for conservation purposes.
Patterns of morphological diversity and phenotypic differentiation of M. schenckii compared with other species of columnar cacti are apparently related to both management intensity and biological features of the species (Table 5). Polaskia chende is the least morphologically diverse and has the lowest phenotypical differentiation among wild and managed populations. This species is also the least intensely managed of the species compared, and the enrichment of managed stands with selected new variants is more difficult than in the other species because of difficulties controlling vegetative propagation and its slow growth. Differentiation among wild and managed populations is low since artificial selection operating in managed stands is slighter than in the other species compared, and because gene flow among populations is high (Cruz and Casas 2002). Polaskia chichipe and M. schenckii, with higher morphological differentiation and phenotypic divergence, are comparable with P. chende since they are pollinated by diurnal insects, their seeds are dispersed by birds, and they have successful establishment in wild and managed populations (Otero-Arnaiz et al. 2003; Ortíz et al. in press). Therefore, both pollen and seed flow among wild and managed populations are high. In these cases, however, the higher morphological diversity and phenotypic differentiation between wild and managed populations could be explained by the higher management intensity.
Stenocereus stellatus is the most intensely managed of the species compared and shows also the highest levels of morphological diversity and phenotypic differentiation among wild and cultivated populations. The high variation can be explained because of the high replacement of variants in both agroforestry systems and home gardens (Casas et al. 2006). The high differentiation between wild and cultivated populations is associated with the relatively easy separation from their wild relatives since sexual reproduction is episodic (Casas et al. 2007).
It is possible to expect higher genetic diversity and gene flow in species with higher success of sexual reproduction and, consequently, a lower differentiation among populations. But morphological and population genetics data show the contrary. This pattern can be explained because human intervention through artificial selection may change the natural patterns in artificial populations of agroforestry systems and home gardens. Seeds and vegetative propagules are dispersed and cared for to different degrees by humans in artificial environments, and artificial selection changes phenotype frequencies by favoring abundance of individual plants preferred by people. These actions may alter gene flow and the genetic structure of populations and are more easily conducted in species with easier vegetative propagation and faster growth, intermediate in those species with intermediate capacity of vegetative propagation and growth, and more difficult in species with no vegetative propagation and slow growth. Although these patterns could be explored more deeply in additional cases to complete the panorama, this method appears to have merit as a means of analyzing the relation between management and artificial selection intensity, as well as biological and reproductive aspects of coexisting wild and managed populations of plant resources.
In conclusion, Myrtillocactus schenckii is a plant resource of relatively intermediate cultural value compared with other species of columnar cacti. It is gathered in wild populations but also managed in agroforestry systems and home gardens, artificial selection favoring individual plants producing larger and more fruits, which have increased their abundance in managed populations. This is a sign of an ongoing process of incipient domestication in which plants forming part of managed stands do not depend on humans to survive and reproduce. Gene flow among wild and managed populations appears to be high and, therefore, artificial selection appears to be the main factor explaining the morphological differentiation found in this study. Both agroforestry systems and home gardens are important scenarios for studying plant management, artificial selection, and domestication, and for conservation programs.
Literature Cited
Blancas, J. J., F. Parra, J. D. Lucio, M. E. Ruíz-Durán, E. Pérez-Negrón, A. Otero-Arnaiz, N. Pérez-Nasser, and A. Casas. 2006. Manejo tradicional y conservación de la diversidad morfológica y genética de Polaskia spp. (Cactaceae) en México. Zonas Áridas 10:20–40.
Bravo-Hollis, H. 1978. Las cactáceas de México. Universidad Nacional Autónoma de México, México, Volumen I.
Casas, A., J. Caballero, and A. Valiente-Banuet. 1999a. Use, Management and Domestication of Columnar Cacti in South-central Mexico: A Historical Perspective. Journal of Ethnobiology 19:71–95.
———, A. Valiente-Banuet, A. Rojas-Martínez, and P. Dávila. 1999b. Morphological Variation and the Process of Domestication of the Columnar Cactus Stenocereus stellatus in Central Mexico. American Journal of Botany 86:522–533.
———, ———, J. Viveros, J. Caballero, L. Cortés, P. Dávila, R. Lira, and I. Rodríguez. 2001. Plant Resources of the Tehuacán-Cuicatlán Valley, Mexico. Economic Botany 55(1):129–166.
———, J. Cruse, E. Morales, A. Otero-Arnaiz, and A. Valiente-Banuet. 2006. Maintenance of Phenotypic and Genotypic Diversity of Stenocereus stellatus (Cactaceae) by Indigenous Peoples in Central Mexico. Biodiversity and Conservation 15:879–898.
———, A. Otero-Arnaiz, E. Pérez-Negrón, and A. Valiente-Banuet. 2007. In Situ Management and Domestication of Plants in Mesoamerica. Annals of Botany 100:1101–1115.
———, S. Rangel-Landa, I. Torres-García, E. Pérez-Negrón, L. Solís, F. Parra, A. Delgado, J. J. Blancas, B. Farfán, and A. I. Moreno. 2008. In Situ Management and Conservation of Plant Resources in the Tehuacán-Cuicatlán Valley, Mexico: An Ethnobotanical and Ecological Perspective. Pages 1–25 in U. P. De Albuquerque and M. Alves-Ramos, eds., Current Topics in Ethnobotany. Research Signpost, Kerala, India.
Cruz, M. and A. Casas. 2002. Morphological Variation and Reproductive Biology of Polaskia chende (Cactaceae) under Domestication in Central Mexico. Journal of Arid Environments 51:561–576.
González-Insuasti, M. S. and J. Caballero. 2007. Managing Plant Resources: How Intensive Can It Be? Human Ecology 35:303–314.
Guillén, S., A. Casas, J. Benítez, and M. Martínez. 2009. Seed Germination of Wild, Managed in Situ and Cultivated Populations of Columnar Cacti in the Tehuacán-Cuicatlán Valley. Journal of Arid Environments 73:407–413.
Hawkes, J. G. 1983. The Diversity of Crop Plants. Harvard University Press, London.
JMP software Version 5.0 of the SAS System for Macintosh. Copyright © 1996. SAS Institute, Inc., Cary, North Carolina
Lira, R., A. Casas, R. Rosas, M. Paredes, E. Pérez-Negrón, S. Rangel-Landa, L. Solís, I. Torres, and P. Dávila. 2009. Traditional Knowledge and Useful Plant Richness in the Tehuacán- CuicatlánValley, Mexico. Economic Botany 63(3):271–287.
MacNeish, R. S. 1967. A Summary of Subsistence. Pages 290–309 in D. S. Byers, ed., The Prehistory of the Tehuacán Valley. Vo1. 1. Environment and Subsistence. University of Texas Press, Austin.
Nei, M. 1972. Genetic Distance between Populations. The American Naturalist 106:283–292.
Oaxaca-Villa, B., A. Casas, and A. Valiente-Banuet. 2006. Reproductive Biology in Wild and Silvicultural Management Populations of Escontria chiotilla (Cactaceae) in the TehuacánValley, Central México. Genetic Resources and Crop Evolution 53:277–287.
Ortíz, F., K. Stoner, E. Pérez-Negrón, and A. Casas. (in press). Pollination Biology of Myrtillocactus schenckii (Cactaceae) in Wild and Managed Populations of the Tehuacan Valley, Mexico. Journal of Arid Environments.
Otero-Arnaiz, A., A. Casas, M. C. Bartolo, E. Pérez-Negrón, and A. Valiente-Banuet. 2003. Evolution of Polaskia chichipe (Cactaceae) under Domestication in the Tehuacan Valley, Central México: Reproductive Biology. American Journal of Botany 90:593–602.
Otero-Arnaiz, A. C., J. L. Hamrick, and J. Cruse. 2005. Genetic Variation and Evolution of Polaskia chichipe (Cactaceae) under Domestication in the Tehuacán Valley, Central Mexico, Analyzed by Microsatellite Polymorphism. Molecular Ecology 14:1603–1611.
Parra, F., N. Pérez-Nasser, D. Pérez-Salicrup, R. Lira, and A. Casas. 2008. Population Genetics and Process of Domestication of Stenocereus pruinosus in the Tehuacán Valley, Mexico. Journal of Arid Environments 72:1997–2010.
Rodríguez-Arévalo, I., A. Casas, J. Campos, and R. Lira. 2006. Uso, manejo y proceso de domestication de Pachycereus hollianus (F.A.C. Weber) Buxb. (Cactaceae) en el Valle de Tehuacán-Cuicatlán, Mexico. Interciencia 31:677–685.
Rohlf, J. 1993. Numerical Taxonomy and Multivariate Analysis System for the PC Microcomputer (and Compatibles). Version 2.6. Applied Biostatistics, Inc., New York.
Sneath, P. H. A. and R. R. Sokal. 1973. Numerical Taxonomy. The Principles and Practice of Numerical Classification, Freeman, San Francisco, California.
Tinoco, A., A. Casas, R. Luna, and K. Oyama. 2005. Population Genetics of Wild and Silvicultural Managed Populations of Escontria chiotilla in the Tehuacán Valley, Central Mexico. Genetic Resources and Crop Evolution 52:525–538.
Acknowledgments
The authors thank the Dirección General de Asuntos del Personal Académico, UNAM, Mexico (DGAPA, research projects PAPIIT-IN220005 and IN219608), and the Royal Botanic Gardens at Kew (Millennium Seed Bank), U.K., for financial support. We also thank Edgar Pérez Negrón for support in field work, and people of San Luis Atolotitlán and Coatepec, Puebla, for their hospitality and valuable collaboration in the research.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article
Cite this article
Blancas, J., Casas, A., Lira, R. et al. Traditional Management and Morphological Patterns of Myrtillocactus schenckii (Cactaceae) in the Tehuacán Valley, Central Mexico1 . Econ Bot 63, 375–387 (2009). https://doi.org/10.1007/s12231-009-9095-2
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s12231-009-9095-2