
Abstract
The Mediterranean flora is spectacularly rich in orchid species that have evolved remarkable adaptations to their environment. Orchids have complex and delicate interactions with their pollinators, which makes them particularly prone to local extinction. Conservation actions should be encouraged for a range of endangered Mediterranean orchid species, but the current taxonomic confusion in several genera and the apparent disagreement among orchid taxonomists make the situation particularly confusing from a conservation perspective. In this review, we document how the different pollination syndromes of Mediterranean orchids (nectar reward, shelter offering, food deception and sexual deception) can have a profound impact on the type of reproductive barriers among species, on floral phenotypic variation as we perceive it, on potentially related processes of species sorting and extinction and, consequently, should have a strong influence on the related conservation management programs. We also highlight that the majority of Mediterranean orchids are pollinated by specialised bees often occupying otherwise narrow ecological niches (e.g. pollen specialisation, brood cell parasites, specific nesting site). This condition makes the orchid-pollinator interactions very fragile and several orchid species prone to local extinction. We illustrate this phenomenon by a selection of case studies that show how the adequate integration of the ecological requirements/traits of the orchids and their associated pollinators into conservation actions could help protect endangered species and ensure the sustainability of the often complex local pollination web.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
The Mediterranean region is geographically and ecologically heterogeneous (Quézel, 1985; Thompson, 2005) and is a difficult area to characterize because its political delineation does not coincide with any of its bioclimatic, biogeographically or floristic outlines. The whole Mediterranean region comprises approximately 24–25,000 plant species (or approximately 29–30,500 taxa including subspecies) (Quézel, 1985; Greuter, 1991; Heywood, 1995). This region is one of the world’s 25 biodiversity “hot spots” for plant biodiversity and encompasses a wide range of terrestrial orchids, mostly belonging to tribe Orchidinae and Limodorinae. Orchids are often thought of as delicate and colourful by the laymen, but scientists also regard them as perhaps the finest illustrative examples of reciprocal adaptation in insect-plant interactions (Darwin, 1862; Van der Pijl & Dodson, 1966; Alcock, 2005). While members of Limodorinae are typical of forest habitats and are generally rewarding, the members of Orchidinae are more common in open and sunny meadows and typically offer no reward to their pollinators. Among Orchidinae, with the interesting exception of the Alpine genera, such as Nigritella, Pseudorchis, Traunsteinera, which offer nectar as reward, most of the members of related genera, with a more Mediterranean-centred distribution, including Anacamptis, Dactylorhiza, Neotinea and Orchis, are food-deceptive, i.e. they have large and conspicuous flowers and attract and deceive mostly naive pollinators by emitting floral signals similar to those of non-orchid nectarifeous plants (Dafni, 1984, 1986; Ackerman, 1986).
Food deception is not the only deceptive mechanism present in Mediterranean orchids. In fact, species of genus Serapias mimic nests and shelter for insects (Dafni et al., 1981) while members of the genus Ophrys attract and deceive their pollinators through an elaborate and highly specific sexual mimicry that involves visual traits and odour signals that mimic the pheromone of female insects (Kullenberg, 1961). This ample variety of pollination systems has traditionally attracted evolutionary biologists since Darwin’s time. The literary backbone of The Various Contrivances by which Orchids are Fertilised by Insects (Darwin, 1862) is a long-standing argument describing the evidence for pollinator specialisations and adaptations to outcrossing. The orchids form a group that contains a long series of red-listed species (therefore important from a conservation perspective) and thus represents a challenging framework for addressing conservation problems and setting up management programs.
The Mediterranean region has been profoundly altered by human activities over the past 10,000 years. Much of its native vegetation has undergone dramatic changes during this process, leading to the formation of many secondary or subseral communities such as the characteristic shrubland communities (maquis, phrygana, matorral, garigue, etc.) that form a conspicuous part of contemporary Mediterranean landscapes (Thompson, 2005 and references therein). Orchids are particularly sensitive to such habitat changes. In fact, while dealing with orchid conservation there is a need to consider the physical as well the biological factors which determine orchids’ existence in relation to human disturbances and their effects. Swartz and Dixon (2009) have stressed the need to link drivers of rarity/decline in orchids to changes in habitats and pollinator requirements. In addition to all these factors, there is a need to consider the highly specific mycorrihza relations that are essential to the orchid’s survival (Rasmussen, 2002) and the relationship with their pollinators. Here we focus on the latter aspect of orchid biology and discuss the impact of pollination strategies, pollinator community changes and pollinator needs on orchid conservation.
The Taxonomy of Mediterranean Orchids and Species Delineation
The large controversy surrounding the taxonomy of orchid genera around the Mediterranean is a major obstacle towards planning conservation actions. The taxonomy of several Mediterranean orchid genera is still subject to intensive debates among orchidologists, primarily because of a general disagreement on the diagnostic value of different suites of morphological characters used for species delineation. Another reason is the parallel outbursts of new species described every year in non-refereed journals by some authors, which are almost immediately published again as synonyms by representatives of the opposite school of thought (Delforge, 1994, 2005 vs. Pedersen and Faurholdt, 2007; Devey et al., 2008).
This taxonomic ballet has far-reaching implications, not only from the purely practical perspective of ascribing a specimen to a given species, which sometimes turns out to be as close as it gets to “mission impossible”, but also from a conservation angle. In Ophrys, for example there are no less than about 530 names in the literature (Global Biodiversity Information Facility). Delforge (2005) recognised 252 species, while on the other extreme, Pedersen and Faurholdt (2007) recognise 16 species and 46 subspecies. Both approaches reflect two radically opposite perspectives but, admittedly, do not add a constructive contribution to the needed debate on the real evolutionary dynamics of local natural variation and of those selective factors as habitat constraints and pollinator-mediated selection. Moreover, even if in the recent years the molecular work on several genera (such as Orchis and Anacamptis) (Bateman, 2001; Cozzolino et al., 2003), Ophrys (Devey et al., 2008); Dactylorhiza (Pillon et al., 2006); Serapias (Bellusci et al., 2008), Epipactis (Squirrell et al., 2002) has largely contributed in recognizing patterns in systematics, we are still far from providing a clear and defined picture of the systematic relationship in the more complex genera (as Ophrys and Epipactis) that can help resolve the strong taxonomic controversy.
These taxonomic inconsistencies are obviously an obstacle to conservation because there is often a lack of communication between taxonomists and conservationists due to the largely non-overlaypping relevant literature. Theoretical debates between evolutionary biologists on species delimitation may have significant effect on conservation attention devoted to specific taxa. If a taxon (or several taxa) has been named as a species then it is expected to be considered seriously by conservationists. Each author, by using different criteria, has often proposed his own diagnostic criteria and the discrepancy between several different taxonomic treatments for the same orchid taxon (particularly for the sexually deceptive Ophrys) has generated uncertainty.
A large number of debatable taxa make it difficult to produce any checklist, which should be the first practical step towards characterizing and quantifying the local biodiversity. Given that micro-species are by definition, rare as well as narrow endemics, they should receive high conservation attention (Pillon & Chase, 2007). Thus, taxonomically difficult groups tend to have more endangered species and tend to be over-represented in red data lists (Pilgrim et al., 2004). As a result of this long-standing taxonomical confusion, critical issues with far-reaching implications from a conservation angle have emerged:
-
1.
The accurate identification of a given taxon as endemic, micro-species or local variant is subject to controversy. For example, Ophrys fusca ssp. fusca which includes at least 42 synonyms (sensu Pedersen and Faurholdt, 2007);
-
2.
Any decision concerning species delineation may instantly change the “identity” as well as the distribution range of a given taxon and, as a result, the policy and measures to be taken concerning its conservation;
-
3.
The confusion renders useless any categories threatened (Lucas & Synge, 1978; Mace & Lande, 1991) and rarity (Rabinowitz, 1981; Sapir et al., 2003) that are key to any conservation action.
Pollination Specialisation and Floral Variation
The taxonomic value attributed to the natural morphological polymorphism within and among populations is a decisive aspect that should be taken into account by taxonomists and conservationists. From a conservation perspective there will be striking differences if this variation reflects species differences, or if it is solely regarded as intra-specific polymorphism from a local biological/ecological perspective. To reach a better understanding of the processes generating such natural variation (both morphological and genetic), we should pay more attention to orchid-pollinator interactions. A deeper knowledge of species biology, particularly species-specific pollination strategies and pollinator behaviour, may help reaching a better understanding of the potential evolutionary consequences of natural variation that occurs among individuals and populations. Thus, adequate conservation efforts should be taken for preserving such variation. Indeed, estimating and understanding the forces shaping natural variation may allow us to distinguish among the even subtly distinct morphological variants (often emphasized in species-rich classifications) generated by stochastic forces in species and populations dynamics (such as genetic drift and population demographic history) and those generated by natural selection and local adaptation. All these factors deserve conservation attention.
In this regard, the different nature of pollination strategies has a direct impact on constraining floral forms and in generating and maintaining of local variation. Nevertheless, to estimate the functional role of natural variation in flower shape, it is important to dissect the relative contribution that each floral trait (scent, colour, floral morphological display) has for pollination attraction. For instance, food deceptive orchids typically attract a wide range of pollinators and their species boundaries are well maintained by strong post-mating isolation rather than by strong floral isolation (Scopece et al., 2007). A single food deceptive species typically attracts a broad spectrum of pollinator species (however, typically from one or few functional groups) and an effective pollinator overlap among different food deceptive orchids has been often reported (Dafni & Ivri, 1979; Cozzolino et al., 2005). Here, visual floral signal (colour and shape) rather than floral scent are thought to play a major role in mediating pollinator attraction (Dafni, 1987; Gumbert & Kunze, 2001). However, very few cases of Batesian mimicry involving a single model are known (Dafni & Ivri 1981; Nilsson, 1983). The strategy of generalised food deception (or perceptual exploitation) is likely to be more effective when orchids grow in habitats that are home to a wide range of foraging insects.
Given that several partners are included in this plant-animal interaction (see Fig 2), it is less likely that a single pollinator species will exert a strong directional selection on floral traits. In fact, excluding the flower colour, where a magnet effect from surrounding rewarding flower has been seen, there is no strong morphological specialisation in any single floral trait (nectar spur, labellum) to improve pollination performance of a single specific pollinator species (Ushimaru & Nakata, 2001). More importantly, it has often been suggested that individual variation in floral traits may retard the learning behaviour of the pollinators (Wright & Schiestl, 2009).
Surprisingly, empirical data testing the action of pollinator-mediated natural selection on floral traits are still lacking for food deceptive Mediterranean orchids (but see Jacquemyn & Brys 2009). The few studies that are available from other non-Mediterranean food deceptive orchid groups (O’Connell & Johnson, 1998; Sabat & Ackerman, 1996) have failed to detect strong directional selection on floral traits. Comparative analysis of scent emission in rewarding and food deceptive orchids revealed larger scent variation in food deceptive orchids than rewarding ones and high intra-individual variation (Salzmann et al., 2007a, b). These findings suggest the absence of any significant directional selection on floral traits for food deceptive orchids, because several different pollinators were involved in this interaction (i.e. relaxed selection).
Thus, for food deceptive orchids, local variation in floral traits, particularly in morphological traits but also in floral scent among distant populations, are more likely to reflect stochastic processes such as genetic drift or relaxed selection on these traits. Under this scenario, this variation should not be favoured both from a taxonomic perspective and from the relative conservation priorities.
In contrast, in the genus Ophrys, due to the dominance of scent emission in pollinator attraction, the relative contribution of floral traits variation in scent and morphology should be more carefully dissected. While floral scent emission is usually highly specific to match the female partner of pollinator, other floral traits such as labellum and lateral tepals, play a minor role in pollinator attraction (Vereecken & Schiestl, 2009; but see by Spaethe et al., 2007). Thus, stochastic variations in traits are expected to be greater in the latter than in the former and have less evolutionary importance than corresponding variation in flower scent emission. As consequence, (and in spite of what has been traditionally documented in Ophrys), small morphological variations in flower traits should not deserve any enthusiastic taxonomic interest or strong conservation attention.
By contrast, different odour variants in Ophrys may be the result of an adaptation of spatially-varying models or pollinator preferences (Vereecken & Schiestl, 2008). Isolated Ophrys populations that are subject to strong selection by local pollinators may experience strong local differentiation of odour bouquet that may represent a first step in the evolution of stronger reproductive isolation (and thus speciation). By using GC analyses of virgin female bee extracts Vereecken et al., (2007) found significant differences in sex pheromone blends among populations in the solitary bee Colletes cunicularius (Hym. Colletidae), an important pollinator of Ophrys species, especially in the O. exaltata group (Delforge, 2005; Mant et al., 2005). This study also revealed that the divergence in female sex pheromone blends was positively correlated with the geographic distance among populations, which suggests that genetic divergence among distant insect populations can affect their sex pheromone chemistry. By performing field bioassays with synthetic sex pheromones, Vereecken et al., (2007) showed that males of C. cunicularius had a strong preference for sex pheromone extracts of allopatric female bees, which led to testing the prediction that the orchids should evolve “exotic” odour bouquets that slightly differ from the female bees within populations, and thus, be preferred by C. cunicularius males. These authors also found that the floral scent of O. exaltata was adapted to the local preferences of patrolling C. cunicularius males for “exotic” odour blends, and that the higher behavioural activity in male bees in response to the floral odours was caused by the deviant ratio in the odour components between the female bees and the sympatric orchids (Vereecken & Schiestl, 2008). In this context, local “crosstalk” between orchids and its pollinator partner represents a crucial step in the evolutionary dynamics of sexually deceptive species and deserves special attention for conservation perspective.
Pollination Specificity, Hybridization and Hybrid Conservation
Pollination strategies and their specificities also have a strong impact on the frequency and amount of possible gene flow among different orchid species. From a conservation perspective, natural hybridization has traditionally been seen with contrasting points of view. More often, hybridization has been considered a threat for the endangered species because of the risk of genetic assimilation with the widespread one (Rhymer & Simberloff, 1996). However, the role of hybridization as an evolutionary force driving speciation and adaptation through increasing genetic diversity within taxa has been recognised in several studies, highlighting the need for a more rigorous evaluation of this phenomenon by conservationists (Arnold, 1997). Clearly, from a traditional perspective, if a positive rule in generating natural biodiversity is acknowledged, then hybrid zones can be seen as evolutionary arenas where novel genetic combination arose and increase natural variation.
Therefore, in order to make accurate evaluations of the conservation status of hybridization and related hybrid zones, it is imperative to properly identify the nature of the ongoing evolutionary processes. Obviously, the strength, frequency and output of hybridization strongly depend on the nature of species boundaries and on the type of reproductive isolation existing among orchid species. Consequently, a better appraisal of these barriers and on their interactions in contact zones represents a crucial tool for the understanding of the dynamic and evolutionary outputs of hybrid zones. Consequently, Mediterranean orchids, depending on their pollination biology, differ strikingly in the amount of pre-mating and post-mating isolation.
Hybridization, particularly in the Mediterranean basin, is a relatively common phenomenon for food deceptive orchids (Dafni, 1987; Cozzolino & Widmer, 2005). Due to the nature of the pollination strategy in deceptive orchids, where increased pollinator sharing is expected, hybrid individuals mainly represent the secondary product of a food deceptive pollination syndrome, and are continuously produced when parental species occur sympatrically. The evidence from the literature largely indicates that in the case of food deceptive orchids, hybridization is unlikely to be a threat, and does not lead to a loss of orchid biodiversity as suggested for other plant groups. Due to strong post-zygotic reproductive isolation among parental species, interspecific hybrids are relatively unfit, so strong post-zygotic reproductive isolation barriers (i.e. chromosomal differences (Cozzolino et al., 2004)) consistently maintain and protect parental species from large introgression. Furthermore, the risk of species extinction through genetic assimilation must be considered an extremely uncommon event, due to the presence of strong post-zygotic isolating barriers that prevents significant level of genetic introgression. Thus, hybridization in food deceptive orchids has no evolutionary or conservation importance.
However, sympatric zones (particularly those where several different orchid species occur together, such as the Mediterranean basin) could represent places in which to test the strength of reproductive isolation barriers against species coexistence. In some cases it has also be noted that these sympatric areas represent instances where natural selection appears to promote divergence in species traits leading to pollinator-mediated fitness benefits and possibly contributing to the further build-up of reproductive isolation (secondary character displacement; Cozzolino et al., 2005). Under these circumstances, sympatric areas, rather than hybrid zones and hybrid plants per se, can be considered as an evolutionary laboratory to reinforce reproductive isolation of orchid species, warranting an elevated conservation status. This is also true for the few clades of strictly Mediterranean-distributed rewarding orchids (i.e. A. coriophora clade) which when hybridizing with related food deceptive species (A. collina), are always strongly sheltered from extensive introgression (Dafni and Ivri, 1979).
A very different interpretation of the hybridization process and its evolutionary output and related conservation effort should be considered when dealing with sexually deceptive orchids. Here, postmating isolation is highly limited and the species boundary is mainly maintained by premating isolation due to specificity of pollinator attraction. However, when hybridizing, due to absence of other late postmating barriers that halt interspecific gene flow, sexually deceptive hybrid zones are formed by hybrids of different genetic categories and large introgression occurs among parental species (Stökl et al., 2008). Under this scenario, the hybrid zone may produce a wide genetic merging of parental species. However, the occurrence of several recombination events and the make-up of new genetic combinations may also provide the substrate for new evolutionary trajectories and speciation avenues. In fact, while in food deceptive orchids hybrids are less fit than parental species and thus unable to acquire novel pollinator niches (Dafni & Baumann, 1982), in sexually deceptive orchids the genetic admixture of parental species may produce novel scent combinations in hybrid individuals. Furthermore, these novel scents may also be more attractive to different pollinators than the scents produced by parental species, thus allowing an immediate speciation (Vereecken et al., 2010).
Finally, it should be stressed that even if hybridization does not lead to speciation, it does not imply that it may not provide some significant contribution to promote and facilitate the local genetic adaptation of orchid species. Hybridization may also represent a genetic bridge across the species boundary to transfer highly advantage alleles for local adaptation, as in the case of species that recently colonised new habitats. This could be the case for the North European Dactylorhiza species, where extensive hybridization (often followed by polyploidization) generated the genetic background for promoting species distribution in northern European hemisphere (Hedrén, 2002).
Pollinator Types and Pollen Dispersal
Orchid dispersion and interpopulation gene flow is traditionally considered to occur primarily through seeds (Ackerman & Ward, 1999). This view is largely based on the empirical observation of their minute, almost dust-like seeds, but recent studies have provided evidence that most seeds typically disperse in the near vicinity of the maternal plant (Chung et al., 2004). These results are consistent with mathematical models (e.g., a simple ballistic model) for anemochorous (i.e., wind-dispersed) seeds and wind tunnel experiments, indicating that most orchid seeds are expected to germinate close to the maternal plant (<6 m), which in turn supports a prediction of significant fine-scale genetic structure within populations (Murren & Ellison, 1998). Parallel studies on the genetic structure of orchid colonies (such as Cephalanthera longibracteata, Orchis purpurea, etc.) support the prediction that the majority of seeds are dispersed over distances of less than 10 m for terrestrial orchids (Chung et al., 2004).
These and other recent advances challenge the mainstream hypothesis that orchid seeds represent the main (and eventually the only) gene dispersal mean. In particular, estimations of the ratio between pollen and seed flow among populations confirmed this finding, as in the north America introduced Epipactis helleborine, where this value reached the extreme value 1.43 and in Anacamptis palustris where the calculation yielded a value of 0.48 (Cozzolino et al., 2003). A similar experimental approach showed that the pollen to seed flow ratio varies among populations and, in some circumstances, pollen flow contributes much more to gene flow than seed dispersal (see evidence in the neotropical epiphytic orchid, Laelia rubescens,Trapnell et al., 2004). These results have important implications for conservation programmes, since they challenge the popular belief that pollen flow contributed less to population connectedness than seed dispersal. Thus, it can be postulated that even in instances where pollen dispersal seems less important than seed dispersal (in fact, orchids are among seed plants with lower values for pollen/seed flow (Cozzolino et al., 2003)), the relative contribution of pollen dispersal might be very important for the maintenance of population genetic diversity within related orchid patches while long distance seed dispersion remains the basic mechanism for the invasion of new habitats and gene flow among distant populations.
Pollen dispersal distances within and among populations are therefore likely to vary according to the pollen vector species and pollinator-mediated pollen flow is determined by the pollination strategy of the orchid (e.g. sexually deceptive vs. food deceptive vs. rewarding). On average, female solitary bees are thought to forage within a few hundreds metres around their nest (Gathmann & Tscharntke, 2002). The patrolling area of males in search of females has been largely overlooked in wild bees, but Peakall & Schiestl (2004) have shown that Colletes cunicularius males are mostly found within a very close range (i.e. 50m) of the nesting/emergence site during their reproductive period. It can therefore be postulated that individuals of sexually deceptive orchids such as O. exaltata growing nearby and flowering at time when the males are active are more likely to attract pollinators than other individuals growing a few hundred meters or several kilometers apart. Overall, it seems that body size does well at predicting the foraging range of solitary bees (Greenleaf et al. 2007), although it seems fair to hypothesize that the foraging/patrolling distance is likely to vary from one species to another, between males and females, according to the diet breadth of each gender and the local abundance of suitable forage plants.
To date, the most reliable methods to assess pollinator movements within their habitat are the harmonic radar (Riley, 1980; Osborne et al., 1999; Riley & Smith, 2002) and direct radio-tracking (Pasquet et al., 2008). The results of the few studies using these tools indicate that bees are likely to forage in a range not farther than a few hundred meters. Several ground nesting species are known to have spread dramatically and rapidly across Europe over the past decade or so (see Goulson & Williams, 2001 on Bombus hypnorum and Vereecken et al. 2009 on Colletes hederae), which implies that estimating their foraging range does not provide the full picture about dispersal capabilities of wild bees, their inter-population connectedness and their capability to invade new territories.
Different representatives of local pollinators also influence pollen dispersal. Moths and butterflies are the main pollinator of the deceptive Anacamptis pyramidalis and their relative representation has been found to vary between local orchid populations: butterflies are the main pollen vector in forested areas while burnet moths dominate more open areas. In mark-release-recapture studies, the two main pollen vectors exhibited different flight behaviour and consequently, they differed in relation to pollen dispersal. The diurnal Aproia crategi was generally found moving much longer distances than Zygaena minos and transferred pollen among more distant orchid patches. Thus, while Zygaena was more effective in moving pollen among close plants, Aproia was a more important vector for transferring pollen among distant patches (Lind et al., 2007). Since orchid pollen vectors do not depend on deceptive orchid for their needs, clearly environmental variations (e.g. induced by human activities) that produce changes in local pollinator community can alter the representation of those pollinators that export pollen to longer distances, thus producing an indirect effect on among-populations gene flow and consequently increase genetic drift and inbreeding in fragmented populations.
More experimental studies are needed in order to understand the effective link between distance in pollen transfer and genetic diversity in orchid populations but this can effectively represents an important limiting factor, particularly for those endangered orchid species that are patchily distributed.
Identity and Decline of Orchid Pollinators in Europe
Most orchid species in the Mediterranean are strictly dependent upon insect pollinators to ensure their reproduction. There are a few exceptions to this rule, notably in the genera Epipactis, Cephalanthera, Serapias (S. parviflora), Ophrys (O. apifera), Neotinea (N. maculata), where spontaneous selfing is common, but overall, the flowers of most Mediterranean orchid species are entomophilous. Pollinator records include representatives primarily in the orders Diptera, Hymenoptera and Lepidoptera. The ecology of these pollen vectors differs to a large extent, but they nevertheless share one very important common denominator: the decline of their natural populations all over Europe over the past few decades. This phenomenon has been particularly well documented in wild bees (e.g. Murray et al., 2009; Williams & Osborne, 2009). These reports on decline have documented changes in the pollinator fauna way beyond the expected spatio-temporal variation observed in the overall distribution and local abundance of these insects in their natural habitats (Rosenzweig, 1995; Roubik, 2001; Oertli et al., 2005; Wojcik et al., 2008).
An appraisal of the major drivers of decline has pinpointed habitat loss, habitat fragmentation and the associated decline in suitable forage plants as the main threats to the sustainability of pollinator’s populations and pollinator services (Shepherd et al., 2003; Biesmeijer et al., 2006). Habitats rich in plant species are an essential prerequisite to harbour a diverse array of insect pollinators, pointing towards bee’s climatic and habitat specialisation.
To illustrate all orchid-pollinator interactions and how to conserve them by incorporating relevant data on autecology of both orchids and their pollinators is beyond the scope of this article. Furthermore, the details of the ecology of many orchid pollinator species remains poorly understood. Therefore, we will focus this review on a selection of case studies where endangered pollinators and their associated orchids need to be considered with a particular emphasis on the “partial habitat” principle described by Westrich (1996). Specialised pollinators often require a unique combination of nesting sites (along with nest building material) and food plants (including nectar plants, host plants for egg-laying or forage plants for the collection of pollen) within their flight range (Pemberton, this volume). Furthermore, it appears that the sustainability of pollinator populations requires the simultaneous maintenance of all partial habitats (nesting sites and patches of forage/egg-laying plants) that are sometimes isolated by sections of inhospitable environment, rather than the conservation of a geographically-restricted portion of the habitat where pollinators are found to occur on native forage plants (Westrich, 1996).
Pollinator Network—A Selection of Case Studies
Ophrys Orchids and Their Specialised Bees
By and large, the pollination of Ophrys orchids is mediated by male bees (Hym. Apoidea), although several species of wasps, beetles and even flies have been reported as well. The male insects bring about cross-pollination as they attempt copulation with the orchid labellum whose floral scent, size and shape resemble females of the targeted species (Kullenberg, 1961; see also Dötterl & Vereecken 2010). Overall, the spectrum of Ophrys pollinators can be divided into three functional groups: food generalist species, food specialists and parasitic specialists. Although the conservation of generalist pollinator species is not strictly dependent upon a narrow range of host plants, it requires sufficient supplies in food along with an appropriate nesting site. However, we will primarily focus on the last two categories, as they provide illustrative examples of how orchid pollinators link specifically to particular resources in their environment.
A large proportion of Ophrys pollinators are solitary bees that have evolved pollen specialisation, i.e. oligolecty (see Dötterl & Vereecken 2010), the females of these species collect from a narrow taxonomic range of host plants (records compiled in Delforge, 2005). Males of these bee species frequently patrol on the preferred host plants for females, and mating takes place on these “rendez-vous” flowers. This strategy has had an impact on the floral display in some of the associated Ophrys, particularly on their perianth that sometimes matches the spectral reflectance of the host plants visited by the females bees, resulting in an increased attractiveness towards patrolling male bees (see Spaethe et al. 2007 on O. heldreichii; but see Vereecken & Schiestl 2009 on O. arachnitiformis). There are several representatives of this kind in the genus Ophrys, and O. fuciflora (Fig. 1a) is presumably no exception. This species with its conspicuous pink perianth is pollinated by males of the long-horned bees Eucera nigrescens (Fig. 1b) and E. longicornis, two species oligolectic on Fabaceae (Westrich, 1989) that have undergone a massive decline in England (Falk, 1991) and in the Netherlands (Peeters & Reemer, 2003). As a result of the decline of its pollinators and habitat loss, this orchid species is severely threatened in UK (Wood, 1989; Lang, 2004), extinct in the Netherlands (Kreutz & Dekker, 2000) and very rare in Belgium (Delforge, 2007). This is the ongoing situation undergone by one of the many endangered orchid species in the Mediterranean, which shows that the decline of pollinators and their host plants can have dramatic snowball effects and drive the decline of the associated orchids over a large geographic scale.

Case studies illustrating the relationships between orchids and their pollinators, and the ecological requirements of the latter. a. Ophrys fuciflora (= O. holoserica), an endangered orchid species in northwestern Europe; b. Female of Eucera nigrescens (Hym. Apidae) on Trifolium pratense (Fabaceae), one of its preferred host plants; c. Male of Ceratina cucurbitina (Hym. Apidae) with pollinia on its head, entering a flower of the orchid Serapias lingua; d. Male of C. cucurbitina inspecting a hollow stem of Rubus sp. (Rosaceae) in which the females build their nest; e. Flower detail of the red helleborine, Cephalanthera rubra (Orchidaceae); f. Female of Chelostoma rapunculi (Hym. Megachilidae), the pollinator of the red helleborine, Cephalanthera rubra, visiting its preferred host plant, a Campanula species (Campanulaceae); g. Male of Chelostoma rapunculi (Hym. Megachilidae) patrolling for females on a Campanula species, the preferred host plant of the females; h. Females of Chelostoma (here, C. florisomne) nest in hollow stems and close their nest entrance with a plug of mud, pebbles and occasionally resin. All photos by NJ Vereecken except E. by Y. Wilcox
Another example of this phenomenon concerns the pollinators characterised by parasitic habits. Several Ophrys pollinators are strictly dependent upon the presence of their insect host(s) to complete their life cycle. For example, females of the scoliid wasp Dasyscolia ciliata (the pollinator of the mirror orchid, O. ciliata = O. speculum) lay their eggs on beetle grubs of several lamellicorn (Scarabaeidae) species living under ground, and the newly hatched wasp larvae slowly consume their paralyzed host in their underground cell (see Clausen, 1940; Vereecken & Carrière, 2003). The digger wasps Argogorytes mystaceus and A. fargeii prey virtually exclusively upon the spittlebug Philaenus spumarius (Hemipt. Cercopidae) (Bitsch et al., 1997), a polyphagous species common in Europe and introduced in North America (Hamilton, 1982).
Finally, males of several cleptoparasitic bee species, so-called “cuckoo bees” because they have evolved the habit of laying their eggs into the nests of other species (Michener, 1974; 2007), are recorded as primary pollinators of orchids that are of particular conservation concern, such as the two Sardinian endemics O. chestermanii and O. normanii, both pollinated specifically by males of Bombus (Psithyrus) vestalis (Paulus & Gack, 1995; Gögler et al., 2009). The queens of these cuckoo or inquiline bumblebees produce no workers, but instead rely on the invasion of colonies of their host (here Bombus (Terrestribombus) terrestris) and subsequently overtake the host queen to force and the “enslave” the host workers of that colony to feed her and her developing young (Sladen, 1912; Goulson, 2003; Benton, 2006). Because their population sizes are generally much smaller than those of their host(s), cuckoo bees are thought to be more vulnerable and more prone to local extinction than their hosts. Evidence from different countries in Western Europe indicates that several Bombus (Psithyrus) species are indeed on the decline primarily as a consequence of anthropogenic activities such as habitat loss and fragmentation affecting both the host bumblebees (Bombus excluding Psithyrus) (Biesmeijer et al., 2006; Kosior et al., 2007; Williams & Osborne, 2009) and their cuckoos (Bombus (Psithyrus)) (Kosior et al. 2007 and references therein).
Serapias lingua and Ceratina cucurbitina
The pollination biology of Serapias orchids in the Mediterranean is usually viewed as a generalised mimicry of nests and shelter (Dafni et al., 1981, Dafni 1984, 1987). This hypothesis rests upon the wide taxonomic range of insect pollinators, both males and females, found immobile into the tubular flowers overnight with pollinia attached to their head. Dafni et al. (1981) found that the dark-coloured flowers of Serapias species accumulate warmth via the sun beams at dawn, providing the pollinators with a morning dose of heat presumably sufficient for them to start foraging earlier than others kept at a lower ambient temperature. Taxonomic studies on the range of their pollinators are urgently needed to uncover if more signals are involved in pollinator attraction. At least one species in the genus Serapias, namely S. lingua, has received more attention so far, and preliminary observations indicate that Ceratina cucurbitina males are key pollinators (Fig. 1c).
This phenomenon, along with the finding of large amounts of alkenes (mono-unsaturated hydrocarbons) in the floral odour extracts of this orchid species (Schiestl & Cozzolino, 2008; Vereecken et al., 2010) suggest that it might have evolved a pollination strategy based on sexual deceit similar to what is observed in Ophrys orchids (Kullenberg, 1961; Van der Cingel, 1995; Delforge, 2005). The females of C. cucurbitina nest in hollow stems where both sexes (virgin females and males) can be found overwintering after the reproductive season (Westrich, 1989; Carrière, 1995). Males of this species patrol nest entrances early in the reproductive season (Fig. 1d), and they are occasionally found pulling females out of their nest to initiate copulation. Contrary to what is observed in the case of several Ophrys pollinators (see “Ophrys Orchids and Their Specialised Bees”), the females of this species are largely polylectic (Westrich, 1989), i.e. they collect pollen from a wide taxonomic range of plant taxa to provision the brood cells of their developing young. Consequently, it appears that the local availability of plants such as Rubus spp. (Rosaceae), Foeniculum sp. (Apiaceae) and other species whose dried stems contains easily excavated pith is essential to the local maintenance of C. cucurbitina and therefore to the pollination of S. lingua (see also Potts et al., 2005).
Cephalanthera rubra and its Pollinators
Cephalanthera rubra (Fig. 1e), a food-deceptive species, attracts several species of bees as pollinators, particularly Chelostoma rapunculi and C. campanularum (Hym. Megachilidae) (Nilsson, 1983), two species whose females are known to be strongly associated with (i.e., oligolectic on) Campanula flowers (Campanulaceae) (Westrich, 1989; Sedivy et al., 2008) (Fig. 1f). Pollinator attraction is reported to be based on the spectral similarity between the rose-coloured flowers of C. rubra and the Campanula species (Nilsson, 1983). Here too, males investigate the Campanula flowers for the presence of conspecific females early in the reproductive season (Fig. 1g) and initiate copulation on these “rendez-vous” flowers. The females require hollow stems in which they make their nest, as well as a supply of resin from neighbouring trees which they collect to craft a plug used to fill the nest entrance (Fig. 1h) when the nest has been provisioned with Campanula pollen and eggs have been laid. The maintenance of large populations of Campanula species whose anthesis coincides more or less phenologically with the activity period of C. rapunculi and C. campanularum, as well as with the flowering time of C. rubra is likely to have a significant positive effect on the reproductive success of this orchid, perhaps through a “magnet effect”, i.e. the ecological facilitation of the pollination in this food-deceptive species through the presence of its “rewarding” model (see Jersáková et al., 2009 for a review).
Orchis caspia and O. israelitica and Their Pollinators
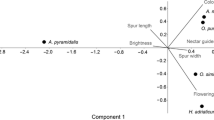
A simplified “fraction” of the pollination web in which Orchis caspia and O. israelitica are involved are presented in Fig. 2. The pollination of the deceptive species depends on the subsidiary of the pollinators by the rewarding ones (Dafni, 1983: Ackerman, 1986; Jersáková et al., 2006, 2009). The data shows that both species are deceptive and share pollinators that are subsidised by several other rewarding species. The species-specific mimic (O. israelitica) has few pollinators and is completely dependent on the pollinators of its rewarding model Bellevalia flexuosa (Dafni and Ivri, 1981; Galizia et al., 2005), while O. caspia which is a general food mimic (Dafni, 1983) attracts several pollinators of various rewarding species, and may have different models in different habitats (Dafni, pers. observations). One pollinator (Eucera clypeata) is also responsible for hybridization and introgression between the two orchids (Dafni and Baumann, 1982). It should be noted that the deceptive species are used by only a few bee species, while some of the rewarding species, which may be visited by some tens of others bee species and are, thus, more flexible in their demands of pollination services.

Fraction of a pollination web involving Anacamptis israelitica, a batesian mimic orchid (*), A. caspia, a generalised food mimic (**) and their hybrid, A. feinbruniae. Each orchid taxon attracts several insect pollinators that are themselves dependent upon certain food plants in their habitat
Analysis of the needs of the pollinators of the main Mediterranean orchid genera and their specificity shows marked differences. The food deceptive orchids attract both males and female insects while Ophrys deception is highly specific due to species –specific chemical sexual attraction of only male bees (Fig. 3). The male bees’ resources, in general, are nectar, sleeping holes and females, while the female needs are pollen as well as nectar, males and suitable nesting microsites. Thus, it is clear that the female floral resources are expected to be more variable than the male’s that are using only nectariferous plants. As a result, the maintenance and the subsidy of the deceptive flowers (as well as of Serapias, which offer a night shelter as a reward for male solitary bees) became dependent on plant species which supply all the resources of both bee sexes as well as the availability of nesting resources.

Interrelationships between deceptive orchids with different pollination syndromes, wild bees and the ecological requirements of the latter, including food plants (nectar and pollen host plants) and nesting sites/materials. The thin lines in the figure represent less frequent instances where e.g. female solitary bees pollinate Serapias orchids or where male bees pollinate food-deceptive orchids such as Anacamptis papilionacea
Discussion and Conclusions
While dealing with orchid conservation there is a need to consider the physical as well the biological factors which determine orchid’s existence in relation to human disturbances and their effects. Swartz and Dixon (2009) pointed out that “although a significant literature exists on the many causes of rarity in plants, drives of rarity in orchids are more often than not linked to their habitats and pollinator requirement…Extrinsic rarity in orchids is a reflection of anthropogenic processes directly limiting or reducing the distribution and abundance of species…”. Fragmentation of habitats, removal of key species critical to the functional sustainability of ecosystems, increased susceptibility to fire threats, pollinator decline and introduction of feral animals are also documented to result in drastic losses in orchid population and diversity (Sosa & Platas, 1998; Coates & Dixon, 2007; Bidartondo & Read, 2008).
Roberts (2003) suggested that orchids are naturally pollinator—limited within a season but resource-limited over their lifetime. Pollination limitation or failure in plants can be attributed to absence of pollinators, failure of removal pollen to reach a recipient, poor quality or insufficient pollen quantity and dispersion of heterospecific pollen (Wilcock & Neiland, 2002), whereas resource limitation may be a reflection of the fitness costs associated with sexual reproduction in the previous season (Ackerman & Montalvo, 1990). Although orchids are ecologically adapted to these limitations, anthropogenic processes such as habitat reduction or fragmentation and over collection may interrupt with a subsequent decrease in fruit set and seedling recruitment (Swartz & Dixon, 2009).
Pollinator limitation appears to be extremely common among orchids (Ackerman, 1986; Calvo, 1993; Tremblay et al., 2005). In an extensive survey of orchid reproductive biology, Tremblay et al. (2005) showed that although resource constraints can sometimes affect orchids (Montalvo & Ackerman, 1987) the majority of evidence pointed to access to pollinators as the dominant limiting factor for reproductive success. A literature survey, of studies in which flowers were subject to supplementary cross-pollination, showed an increase in fruit-set compared with natural populations in 56 out of 57orchid species (Tremblay et al., 2005). Given that orchids are far more reliant on their pollinators than the latter are on the orchid (see Vereecken, 2009), careful conservation planning is required to ensure pollinators requirements such as a food plants, nest sites, larval host species or larval host plants (Kearns et al., 1998; Roberts, 2003; Potts et al., 2005). Thus, understanding of the limitation on reproductive success in orchids and impact of anthropogenic and environmental changes is important in conservation planning and management (Roberts, 2003).
From a conservation perspective, the approach of considering only the habitats physical constraints (i.e. habitat loss, fragmentation and deterioration) overlooks the dependency of rewardless species on the nectariferous ones and the resource needs for the bees’ community service which may dictate also by the patch size as a result of fragmentation. Neiland and Wilcock (1998) found indirect evidence for a dependency of deceptive orchids on other rewarding plants and showed that most rare orchid species do not produce nectar. These findings are consistent with that the fact that non-rewarding orchids have lower fruit set than rewarding ones (Dafni and Ivri; 1979; Gill, 1989; Wilcock & Neiland, 1998), as predicted by Darwin (1862).
Due to the current taxonomical confusion, the large and still unexplained floral variation (the main diagnostic characteristic for any taxonomy) means that the still evolving status of many taxa, including hybrids, makes it impossible to follow any generally recognized conservation policy. The common tools applied for red list data, endemics, rare species and narrow distribution etc. are often rendered useless in the most specific situations, therefore, any decision regarding what to conserve at the local level tends to be far from any common “objective” criteria. Each endangered taxon has to be evaluated under its local and specific circumstances beyond any taxonomic debate with a focus on local variation and spatiotemporal distribution. The conservation approach, at least for those aspects concerning floral biology, has to be converted from “species-oriented approach” to “a pollination web-oriented approach”.
Literature Cited
Ackerman, J. D. 1986. Mechanisms and evolution of food-deceptive pollination systems in orchids. Lindleyana 1: 108–113.
——— & A. M. Montalvo. 1990. Short- and long-term limitation to fruit production in a tropical orchid. Ecology 71: 263–272.
——— & S. Ward. 1999. Genetic variation in a widespread, epiphytic orchid: Where is the evolutionary potential. Systematic Botany 24: 282–291.
Alcock, J. 2005. An enthusiasm for orchids: Sex and deception in plant evolution. Oxford University Press, Oxford.
Arnold, M. L. 1997. Natural hybridization and evolution. Oxford University Press, Oxford.
Bateman, R. M. 2001. Evolution and classification of European orchids: Insight from molecular and morphological characters. Journal Europäischer Orchideen 33: 33–119.
Bellusci, F., G. Pellegrino & A. M. Palermo. 2008. Phylogenetic relationships in the orchid genus Serapias based on monocoding regions of the chloroplast genome. Molecular Phylogenetics and Evolution 47: 986–991.
Benton, T. 2006. Bumblebees, the natural history and identification of the species found in Britain. Collins, London.
Bidartondo, M. I. & D. J. Read. 2008. Fungal specificity bottlenecks during orchid germination and development. Molecular Ecology 17: 3707–3716.
Biesmeijer, J. C., S. P. M. Roberts, M. Reemer, R. Ohlemueller, M. Edwards, T. Peeters, A. Schaffers, S. G. Potts, R. Kleukers, C. D. Thomas, J. Settele & W. E. Kunin. 2006. Parallel declines in pollinators and insect-pollinated plants in Britain and the Netherlands. Science 313: 351–354.
Bitsch, J., T. Barbier, S. F. Gayubo, K. Schmidt & M. Ohl. 1997. Hyménoptères Sphecidae d’Europe Occidentale (Vol. 2.). Faune de France 82: 1–427.
Calvo, R. N. 1993. Evolutionary demography of orchids: Intensity and frequency of pollination and the cost of fruiting. Ecology 74: 1033–1042.
Carrière, J. 1995. Généralités, comportement nidificateur et cycle biologique artificiel pour le genre Ceratina (Latreille, 1802) dans l’Hérault (Hymenoptera, Xylocopinae). Lambillionea XCV 2: 244–254.
Chung, M. Y., J. D. Nason & M. G. Chung. 2004. Spatial genetic structure in populations of the terrestrial orchid Cephalanthera longibracteata (Orchidaceae). American Journal of Botany 91: 52–57.
Clausen, C. P. 1940. Entomophagous insects. McGraw-Hill, New York.
Coates, D. J. & K. W. Dixon. 2007. Current perspective in plant conservation biology. Australian Journal of Botany 55: 187–193.
Cozzolino, S. & A. Widmer. 2005. Orchid diversity: An evolutionary consequences of deception? Trends in Ecology and Evolution 20: 487–494.
———, S. D’Emerico & A. Widmer. 2004. Evidence for reproductive isolate selection in Mediterranean orchids: Karyotype differences compensate for the lack of pollinator specificity. Proceedings of the Royal Society B 271: 259–262.
———, Cafasso D, Pellegrino G, Musacchio A. & Widmer A. 2003. Fine—scale phylogeographical analysis of Mediterranean Anacamptis palustris (orchidaceae) populations based on chloroplast minisatellite and microsatellite variation. Molecular Ecology 12: 2783–2793.
———, F. P. Schiestl, A. Müller, O. De Castro, A. M. Nardella & A. Widmer. 2005. Evidence for pollinator sharing in Mediterranean nectar-mimic orchids: Absence of premating barriers. Proceedings of the Royal Society B 272: 1271–1278.
Dafni, A. 1983. Pollination of O. caspia—a nectarless plant which deceives the pollinators of nectariferous species from other plant families. Journal of Ecology 71: 467–474.
——— 1984. Mimicry and deception in pollination. Annual Review of Ecology and Systematics 15: 259–278.
——— 1986. Floral mimicry—mutualism and unidirectional exploitation of insects by plants. Pp 81–90. In: T. R. F. Southwood & B. E. Juniper (eds). The plant surface and insects. Arnold, London.
——— 1987. Pollination in Orchis and related genera: Evolution from reward to deception. Pp 80–104. In: J. Arditti (ed). Orchid biology: Reviews and perspectives IV. Cornell University Press, Ithaca.
——— & H. Baumann. 1982. Biometrical analysis in populations of Orchis israelitica Baumann and Dafni, O. caspia Trautv and their hybrids. Plant Systematics and Evolution 140: 87–94.
——— & Y. Ivri. 1979. Pollination ecology of and hybridization between Orchis coriophora L. and O. collina. Sol. ex. Russ (Orchidaceae) in Israel. New Phytologist 83: 181–187.
——— & ———. 1981. Floral mimicry between Orchis isrealitica Baumann and Dafni (Orchidaceae) and Bellevalia flexuosa Boiss. (Liliaceae). Oecologia 49: 229–232.
———, ——— & N. B. M. Brantjes. 1981. Pollination of Serapias vomeracea Briq. in Israel by imitation of holes for sleeping solitary male bees (Hym.). Acta Botanica Neerlandica 30: 69–73.
Darwin, C. 1862. The various contrivances by which are fertilized by insects. Murray, London.
Delforge, P. 1994. Guide des Orchide´es d’Europe, d’Afrique du Nord et du Proche-Orient. Delachaux et Niestle, Lausanne.
——— 2005. Guide des orchidées d’Europe, d’Afrique du Nord et du Proche—Orient. Delachaux & Niestlé, Paris.
——— 2007. Guide des orchidées de France, de Suisse et du Bénélux. Delachaux & Niestlé, Paris.
Devey, D. S., R. M. Bateman, M. F. Fay & J. A. Hawkins. 2008. Friends or Relatives? Phylogenetics and species delimitation in the controversial European orchid genus Ophrys. Annals of Botany 101: 385–402.
Dötterl, S. & N.J. Vereecken. 2010. The chemical ecology and evolution of bee-flower interactions: A review and perspectives. Canadian Journal of Zoology, in press.
Falk, S. 1991. A review of scarce and threatened bees, wasp and ants of Great Britain. Research and Survey in Nature Conservation 35, Nature Conservancy Council, UK.
Galizia, C. G., J. Kunze, A. Gumbetrt, A.-K. Borg-Karlson, S. Sachs, C. Markl & R. Menzel. 2005. Relationship of visual and olfactory signal parameters in food-deceptive flower mimicry system. Behavioural Ecology 16: 159–168.
Gathmann, A. & T. Tscharntke. 2002. Foraging ranges of solitary bees. Journal of Animal Ecology 71: 757–764.
Gill, D. E. 1989. Fruiting failure, pollination efficiency and speciation in orchids. Pp 458–481. In: D. Otte & J. A. Endlet (eds). Speciation and its consequences. Sinauer, Sundeland.
Gögler, J., J. Stökl, A. Sramkova, R. Twele, W. Francke, S. Cozzolino, P. Cortis, A. Scrugli & M. Ayasse. 2009. Ménage À Trois—Two Endemic species of deceptive orchids and one pollinator species. Evolution 63: 2222–2234.
Goulson, D. 2003. Bumblebees; their behaviour and ecology. Oxford University Press, Oxford.
——— & P. H. Williams. 2001. Bombus hypnorum (Hymenoptera: Apidae), a new British bumblebee? British Journal of Entomological Natural History 14: 129–131.
Greenleaf, S. S., N. M. Williams, R. Winfree & C. Kremen. 2007. Bee foraging ranges and their relation to body size. Oecologia 153: 589–596.
Greuter, W. 1991. Botanical diversity, endemism, rarity, and extinction in the Mediterranean area: An analysis based on the published volumes of Med-Checklist. Botanical Chronicle 10: 63–79.
Gumbert, A. & J. Kunze. 2001. Colour similarity to rewarding model plants affects pollination in a food deceptive orchid. Biological Journal of the Linnaean Society 72: 419–433.
Hamilton, K. G. A. 1982. Insects and arachnids of Canada Part 10—Homoptera : Cercopidae. Biosystematics Research Institute Publication (Canada Agriculture Canada). PLACE
Hedrén, M. 2002. Speciation patterns in Dactylorhiza incarnata/maculata polyploidy complex (Orchidaceae): evidence from molecular markers. Journal Europäischer Orchideen 34: 707–731.
Heywood, V. H. 1995. The Mediterranean flora in the context of world diversity. Ecologia Mediterranea 21: 11–18.
Jacquemyn, H. & R. Brys. 2009. Temporal and spatial variation in flower and fruit production in a food-deceptive orchid: A five-year study. Plant Biology 12(1): 145–153.
Jersáková, J., S. D. Johnson & P. Kindlmann. 2006. Mechanisms and evolution of deceptive pollination in orchids. Biological Reviews 81: 219–235.
———, ——— & A. Jürgens. 2009. Food deception by plants: From generalized systems to specialized floral mimicry. Pp 223–246. In: F. Baluška (ed). Plant-environment interactions, signaling and communication in plants, from sensory plant biology to active plant behaviour. Springer-Verlag, Berlin Heidelberg.
Kearns, C. A., D. W. Inouye & N. M. Waser. 1998. Endangered mutualisms: The conservation of plant-pollinator interactions. Annual Review of Ecology and Systematics 29: 83–112.
Kosior, A., W. Celary, P. Olejniczak, J. Fijał, W. Król, W. Solarz & P. Płonka. 2007. The decline of the bumble bees and cuckoo bees (Hymenoptera: Apidae: Bombini) of Western and Central Europe. Oryx 41: 79–88.
Kreutz, C. A. J. & H. Dekker. 2000. De orchideeen van Nederland—ecologie, verspreiding, bedreiging, beheer. Uitgave Kreutz and Seckel, Landgraaf & Raalte, The Netherlands.
Kullenberg, B. 1961. Studies in Ophrys pollination. Zoologiska Bidrag Uppsala 34: 1–340.
Lang, D. 2004. Britain’s orchids. WILDguides, English Nature. PLACE
Lind, H., M. Frazen, B. Petterson & L. A. Nilsson. 2007. Metapopulation pollination in the deceptive orchid Anacamptis pyramidalis. Nordic Journal of Botany 25: 176–182.
Lucas, G. B. & H. Synge. 1978. The IUCN plant red data book. International Union for Conservation. WWF.UNEP, Paris.
Mace, G. M. & R. Lande. 1991. Assessing extinction threats: Towards a re-evaluation of IUCN threatened species categories. Conservation Biology 5: 148–157.
Mant, J., R. Peakall, F. P. Schiestle & C. F. Williams. 2005. Does selection on floral odor promote differentiation among populations and species of the sexually deceptive orchid genus Ophrys. Evolution 59: 1449–1463.
Michener, C. D. 1974. The social behaviour of bees—a comparative study. Harvard University Press, Cambridge.
——— 2007. The bees of the world, ed. 2nd. Johns Hopkins University Press, Baltimore.
Montalvo, A. V. & J. D. Ackerman. 1987. Limitations to fruit production in Ionopsis utricularoides (Orchdaceae). Biotropica 19: 24–31.
Murray, T. E., M. Kuhlmann & S. G. Potts. 2009. Conservation ecology of bees: Populations, species and communities. Apidologie 40: 211–236.
Murren, C. J. & A. M. Ellison. 1998. Seed dispersal characteristics of Brassavola nodosa (Orchidaceae). American Journal of Botany 85: 675–680.
Neiland, M. R. M. & C. C. Wilcock. 1998. Fruit set, nectar reward, and rarity in the Orchidaceae. American Journal of Botany 85: 1657–1671.
Nilsson, L. A. 1983. Mimesis of bellflower (Campanula) by the red helleborine orchid Cephalanthera rubra. Nature 305: 799–800.
O’Connell, L. M. & M. O. Johnson. 1998. Male and female pollination success in a deceptive orchid, a selection study. Ecology 79: 1246–1260.
Oertli, S., A. Müller & S. Dorn. 2005. Ecological and seasonal patterns in the diversity of a species-rich bee assemblage (Hymenoptera: Apoidea: Apiformes). European Journal of Entomology 102: 53–63.
Osborne, J. L., S. J. Clark, R. J. Morris, I. H. Williams, J. R. Riley, A. D. Smith, D. R. Reynolds & A. S. Edwards. 1999. A landscape scale study of bumblebee foraging range and constancy, using harmonic radar. Journal of Applied Ecology 36: 519–533.
Pasquet, R. S., A. Peltier, M. B. Hufford, E. Oudin, J. Saulnier, L. Paul, J. T. Knudsen, H. R. Herren & P. Gepts. 2008. Long-distance pollen flow assessment through evaluation of pollinator foraging range suggests transgene escape distances. Proceedings of the National Academy of Sciences of the USA 105: 13456–13461.
Paulus, H. F. & C. Gack. 1995. Zur Pseudokopulation und Bestäubung in der Gattung Ophrys (Orchidaceae) Sardiniens und Korsikas. Jaarbericht Naturwissenschaften Verein. Wuppertal 48: 188–227.
Peakall, R. & F. P. Schiestl. 2004. A mark-recapture study of male Colletes cunicularius bees: Implications for pollination by sexual deception. Behavioural Ecology and Sociobiology 56: 579–584.
Pedersen, H. A. & N. Faurholdt. 2007. The bee orchids of Europe. Kew Publishing, Kew.
Peeters, T. M.J. & M. Reemer. 2003. Bedreigde en verdwenen bijen in Nederland (Apidae s.l.)—Basisrapport met voorstel voor de Rode Lijst. Sichting European Invertebrate Survey—Nederland.
Pilgrim, E. S., M. J. Crawley & K. Dolphin. 2004. Patterns of rarity in the British Flora. Biological Conservation 210: 161–170.
Pillon, Y., M. F. Fay, A. B. Shipunov & M. W. Chase. 2006. Species diversity versus phylogenetic diversity: A practical study in the taxonomically difficult genus Dactylorhiza (Orchidaceae). Biological Conservation 129: 4–13.
——— & M. W. Chase. 2007. Taxonomic exaggeration and its effect on orchid conservation. Conservation Biology 21: 263–265.
Potts, S. G., B. Vulliamy, S. Roberts, C. O’Toole, A. Dafni, G. Ne’eman & P. Willmer. 2005. Role of nesting resources in organising diverse bee communities in a Mediterranean landscape. Ecological Entomology. 30: 78–85.
Quézel, P. 1985. Definition of the Mediterranean region and the origin of its flora. Pp 9–24. In: C. Gomez-Campo (ed). Plant conservation in the Mediterranean area. W. Junk, Dordrecht.
Rabinowitz, D. 1981. Seven forms of rarity. Pp 205–218. In: H. Synge (ed). The biological aspects of rare plant conservation. Wiley, New York.
Rasmussen, M. N. 2002. Recent development in the study of orchid mycorrhiza. Plant and soil 244: 149–163.
Rhymer, J. M. & D. Simberloff. 1996. Extinction by hybridization and introgression. Annual Rreview of Ecology and Systematics 27: 83–109.
Riley, J. R. 1980. Radar as an aid to the study of insect flight. Pp 131–140. In: C. J. Amlaner, C. J. Amlaner, & D. W. Macdonald (eds). Handbook on biotelemetry and radio tracking. Pergamon, Oxford.
——— & A. D. Smith. 2002. Design considerations for an harmonic radar to investigate the flight of insects at low altitude. Computers and Electronics in Agriculture 35: 151–169.
Roberts, D. L. 2003. Pollination biology: The role of sexual reproduction in orchid conservation. Pp 113–136. In: K. W. Dixon, S. P. Kell, R. L. Barrett, & P. J. Cribb (eds). Orchid conservation. Natural History Publications, Kota Kinabalu.
Rosenzweig, M. L. 1995. Species diversity in space and time. Cambridge University Press, Cambridge.
Roubik, D. W. 2001. Ups and downs in pollinator populations: When is there a decline? Conservation Ecology 5(1): 2. URL: http://www.consecol.org/vol5/iss1/art2.
Sabat, A. M. & J. D. Ackerman. 1996. Fruit set in a deceptive orchid: The effect of flowering phenology, display size, and local floral abundance. American Journal of Botany 83: 1181–1186.
Salzmann, C. C., A. M. Nardella & S. Cozzolino. 2007a. Variability in floral scents in rewarding and deceptive orchids: The signature of pollinator-imposed selection? Annals of Botany 100: 757–765.
———, S. Cozzolino & F. P. Schiestl. 2007b. Floral scent in food-deceptive orchids: Species specificity and source of variability. Plant Biology 9: 270–729.
Sapir, Y., A. Shmida & O. Fragman. 2003. Constructing Red Numbers for endangered plant species—Israeli flora as a test case. Journal for Nature Conservation 11: 91–107.
Schiestl, F. P. & S. Cozzolino. 2008. Evolution of sexual mimicry in the Orchidinae: The role of preadaptations in the attraction of male bees as pollinators. BMC Evolutionary Biology 8: 27.
Scopece, G., A. Musacchio, A. Widmer & S. Cozzolino. 2007. Patterns of isolation in Mediterranean deceptive orchids. Evolution 61: 2623–2642.
Sedivy, C., C. J. Praz, A. Müller, A. Widmer & S. Dorn. 2008. Patterns of host-plant choice in bees of the genus Chelostoma: The constraint hypothesis of host-range evolution in bees. Evolution 62: 2487–2507.
Shepherd, M., S. L. Buchmann, M. Vaughan & S. H. Black. 2003. Pollinator conservation handbook. The Xerces Society, Portland Oregon.
Sladen, F. W. L. 1912. The humble-bee, its life history and how to domesticate it, with descriptions of all the British species of Bombus and Psithyrus. MacMillan, London.
Sosa, V. & T. Platas. 1998. Extinction and persistence of rare orchids in Veracruz, Mexico. Conservation Biology 12: 451–455.
Spaethe, J., W. H. Moser & H. F. Paulus. 2007. Increase of pollinator attraction by means of a visual signal in the sexually deceptive orchid, Ophrys heldreichii (Orchidaceae). Plant Systematics and Evolution 264: 31–40.
Squirrell, S., P. M. Hollingsworth & R. M. Bateman. 2002. Taxonomic complexity and breeding system transitions: Conservation genetics of Epipactis leptochila complex (Orchidaceae). Molecular Ecology 11: 1957–1964.
Stökl, J., P. M. Schluter, T. F. Stuessy, H. F. Paulus & G. Assum. 2008. Scent variation and hybridization cause the displacement of a sexual deceptive orchid species. American Journal of Botany 95: 472–481.
Swartz, N. D. & K. W. Dixon. 2009. Terrestrial orchid conservation in the age of extinction. Annals of Botany 104: 543–556.
Thompson, J. D. 2005. Plant evolution in the Mediterranean. Oxford University Press, Oxford.
Trapnell, D. W., J. L. Hamrick & J. D. Nason. 2004. Three- dimensional fire-scale genetic structure of the neotropical epiphytic orchid Laelia rubescens. Molecular Ecology 13: 1111–1118.
Tremblay, R. L., J. D. Ackerman, J. K. Zimmerman & R. N. Calvo. 2005. Variation in sexual reproduction in orchids and its evolutionary consequences: A spasmodic journey to diversification. Biological Journal of the Linnaean Society 84: 1–54.
Ushimaru, A. & K. Nakata. 2001. Evolution of flower allometry and its significance for pollination success in the deceptive orchid Pogonia japonica. International Journal of Plant Science 162: 1307–1311.
Van der Cingel, N. 1995. An atlas of orchid pollination—European orchid. Balkema, Roterdam.
Van der Pijl, N. & C. H. Dodson. 1966. Orchid flowers: Their pollination and evolution. University of Miami Press, Coral Gables.
Vereecken, N. J. 2009. Deceptive behaviour in plants. I. Pollination by sexual deception in orchids: A host-parasite perspective. In: F. Baluska (ed). 203–222 Plant–Environment Interactions—From Sensory Plant Biology to Active Behaviour. Springer Verlag.
——— & J. Carrière. 2003. Contribution à l’étude éthologique de la grande scolie à front jaune, Megascolia maculata flavifrons (F., 1775) (Hymenoptera, Scoliidae) en France méditerranéenne. Notes Fauniques de Gembloux 53: 71–80.
——— & F. P. Schiestl. 2008. The evolution of imperfect floral mimicry. Proceedings of the National Academy of Sciences of the USA 105: 7484–7488.
——— & ———. 2009. On the roles of colour and scent in a specialized floral mimicry system. Annals of Botany 104(6): 1077–1084.
———, J. Mant & F. P. Schiestl. 2007. Population differentiation in female sex pheromone and male preferences in a solitary bee. Behavioural Ecology and Sociobiology 61(5): 811–821.
———, H. Schwenninger, A. Gogala & S. P. M. Roberts. 2009. Mise à jour de la distribution géographique de l’abeille du lierre, Colletes hederae Schmidt & Westrich (Hymenoptera, Colletidae) en Europe. Osmia 3: 2–3.
———, S. Cozzolino & F. P. Schiestl. 2010. Hybrid floral scent novelty drives pollinator shift in sexually deceptive orchids. Editorial Discussion.
Westrich, P. 1989. Die Wildbienen Baden-Württembergs, Teil 1 & 2. Eugen Ulmer, Stuttgart.
——— 1996. Habitat requirements of central European bees and the problem of partial habitats. Pp 1–16. In: A. Matheson, S. L. Buchmann, C. O’Toole, P. Westrich, & I. H. Williams (eds). The Conservation of Bees. Academic, London.
Wilcock, C. & R. Neiland. 1998. Fruit set, nectar reward, and rarity in the Orchidaceae. American Journal of Botany 85: 1657–1671.
——— & ———. 2002. Pollination failure in plants: Why it happens and when it matters. Trends in Plant Science 7: 270–277.
Williams, P. H. & J. L. Osborne. 2009. Bumblebee vulnerability and conservation worldwide. Apidologie 40: 367–387.
Wojcik, V. A., G. W. Frankie, R. W. Thorp & J. L. Hernandez. 2008. Seasonality in bees and their floral resource plants at a constructed urban bee habitat in Berkeley, California. Journal of the Kansas Entomological Society 81: 15–28.
Wood, J. J. 1989. British orchids in their European context. Pp 141–145. In: H. Pritchard (ed). Modern methods in orchid conservation: The role of physiology, ecology and management. Cambridge University Press, Cambridge.
Wright, G. A. & F. Schiestl. 2009. The evolution of floral scent: The influence of olfactory learning by insect pollinators on the honest signaling of floral rewards. Functional Ecology 23: 841–851.
Acknowledgements
We wish to thank Robert Pemberton, Stella Watts and Hong Liu for their critical reading of the paper and for their constructive comments. NJV thanks Yves Wilcox for providing the photograph of the red helleborine used in Fig. 1, and the FRS-FNRS (Belgium) for financial support via a post-doctoral grant.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article
Cite this article
Vereecken, N.J., Dafni, A. & Cozzolino, S. Pollination Syndromes in Mediterranean Orchids—Implications for Speciation, Taxonomy and Conservation. Bot. Rev. 76, 220–240 (2010). https://doi.org/10.1007/s12229-010-9049-5
Published:
Issue Date:
DOI: https://doi.org/10.1007/s12229-010-9049-5