
Abstract
Recent bamboo collections from Brazil, and the study of specimens from several Brazilian herbaria revealed that the species described as Eremocaulon setosum, subtribe Guaduineae, in fact fits within the Arthrostylidiinae genus Aulonemia, based on floral and anatomical characters. Several characters found in this species, such as paniculate determinate synflorescences, solitary spikelets, an abaxial marginal green stripe on the foliage leaf blades, absence of stomata on the entire adaxial side of the leaf blade, and intercostal fibers in the mesophyll are typical features of the subtribe Arthrostylidiinae. The new combination Aulonemia setosa is here proposed, an epitype is designated, and a broader distributional range of this species is documented.
Resumo
Recentes coletas de bambus no Brasil e o estudo de espécimes de diversos herbários brasileiros revelaram que a espécie descrita como Eremocaulon setosum, subtribo Guaduineae, enquadra-se na circunscrição do gênero Aulonemia, da subtribo Arthrostylidiinae, com base nas características florais e anatômicas aqui apresentadas. Algumas características desta espécie, tais como sinflorescências em panículas determinadas, espiguetas simples, uma faixa marginal verde escura na face abaxial das folhas de ramos, ausência de estômatos em toda a superfície adaxial da lâmina e presença de fibras intercostais no mesofilo são características típicas da subtribo Arthrostylidiinae. Com base nos dados apresentados, propõe-se aqui a nova combinação Aulonemia setosa e um epítipo é designado. A ampliação da distribuição geográfica desta espécie é também documentada.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Aulonemia Goudot is a genus of tropical woody bamboo comprising at least 40 species (Judziewicz et al., 1999; Judziewicz & Riina, 2005). The genus is distributed from Mexico to southern Brazil and almost all its species are found in wet high elevation sites. In Brazil, approximately 15 species of Aulonemia are formally known and almost all of them occur in the Atlantic Rain Forest.
The genus is characterized by a set of morphological features (McClure, 1973; Judziewicz et al., 1999) including the ramification type (with one divergent branch per node that occasionally bears some branchlets on its first contracted internodes); hollow internodes (usually); culm leaf blades that are constricted at the base and usually reflexed; foliage leaf sheaths with fimbriae; and paniculate synflorescences bearing solitary spikelets with two to six glumes, two to ten fertile florets, and a rudimentary apical floret represented by a sterile lemma.
Aulonemia is placed within the subtribe Arthrostylidiinae, a neotropical clade of woody bamboos (Bambuseae tribe) that is composed of 13 genera (McClure, 1973; Judziewicz et al., 1999). The species of this subtribe share morphological and anatomical characters that serve to distinguish the subtribe from the Guaduinae subtribe, a closely related clade that is also endemic to the neotropics (Zhang & Clark, 2000; Ruiz-Sanchez et al., 2008). According to Londoño and Clark (2002) and Judziewicz et al. (1999), the following differences distinguish these subtribes : (1) the presence of stomata on both leaf surfaces in Guaduinae versus stomata usually found only on the abaxial surface in other woody bamboos; (2) presence of intercostal layers of sclerenchyma in Arthrostylidiinae; (3) a marginal green stripe on the abaxial surface of foliage leaves of Arthrostylidiinae; (4) midrib of the foliage leaves in median position that is usually prominent in the Guaduinae (this usually associated with the complex vascular system in the midrib of the Guaduinae), but often undetectable in the Arthrostylidiinae (which usually have a single vascular bundle on the midrib; Soderstrom & Ellis, 1987).
Recent bamboo collections from the Atlantic Rain Forest (by P.L. Viana) and the study of specimens from several Brazilian herbaria revealed a distinctive “Aulonemia-like” species with spicate panicles that was initially thought to be a new taxon related to A. setigera (Hack.) McClure, from Rio de Janeiro state.
Further analysis of the literature (Londoño & Clark, 2002) and examination of type collections of Brazilian bamboos showed that this species had been previously described as Eremocaulon setosum Londoño & L. G. Clark, based only on vegetative characters. Some vegetative characters of this taxon that are unusual in Aulonemia led the authors to include it tentatively within the Guaduinae genus Eremocaulon, although its anatomical characters are typically Arthrostylidiinae-like. Its leathery culm leaves with a shiny copper-red adaxial surface and prominent oral setae, thick-walled internodes, and the prominent midrib of the foliage leaf blades are also found in many genera of Guaduinae, such as Guadua and Eremocaulon. Londoño and Clark (2002) stated that the “... generic placement of E. setosum is therefore provisional and may be revised once flowering material and/or molecular data are available”.
A complete morphological characterization of this species, including vegetative characters, floral parts, and anatomical features is provided here. An epitype is designated because the floral parts are critical for the correct application of the name (Art. 9.7 of the Code). The evidence we present should be sufficient to place the species within Aulonemia, following the current morphological concept of the genus (McClure, 1973; Judziewicz et al., 1999). A wider distributional range of this species in the Brazilian Atlantic Forest is documented here.
Materials and methods
This work is based on intensive fieldwork and a review of the main Brazilian herbaria as part of a taxonomic revision of the genus Aulonemia in Brazil by the senior author. For anatomical studies in transverse sections, samples were taken from the central portion, about half-way between the apex and the base of the mature leaf lamina. These samples were fixed in FAA50 (Johansen, 1940) and stored in 70% ethanol, but in some cases they were used immediately after collection, in order to obtain fresh hand-cut sections. For transverse views, hand-cut sections were obtained from the central portion, and submitted to vacuum in order to remove air from intercellular spaces, then double-stained with safranin-astrablue (Bukatsch, 1972), and photographed on an Olympus BH-2 light microscope.
To observe the vasculature in cleared material, fixed samples were processed according to Fuchs (1963), that is, stained with 1% alcoholic fuchsin, soaked in 5% sodium hydroxide solution until clearing was complete, washed and mounted in glycerin jelly. Paradermal sections were obtained by abrasion, from both leaf surfaces, then washed and stained with 1% alcoholic safranin. The following histochemical tests were used: an aqueous solution of 0.02% ruthenium red to stain pectic compounds (Jensen, 1962) and acid phloroglucinol to detect lignin (Johansen, 1940).
For scanning electron microscopy (SEM), leaf samples were removed from the leaf marginal green stripe and midrib regions of herbarium specimens. These samples were glued on aluminum stubs exposing both the adaxial and abaxial surfaces. The mounted and dried samples were coated with gold, and examined on a Quanta 200 SEM (FEI Company) at 20 kV. Terminology of anatomical and micromorphological features follow Ellis (1976, 1979).
Results
Aulonemia setosa (Londoño & L. G. Clark) P. L. Viana & Filg. comb. nov. Basionym: Eremocaulon setosum Londoño & L.G. Clark. Syst. Bot. 27(4): 719, f. 7. 2002. Type: Brazil. São Paulo: Salesópolis, Estação Biológica de Boracéia / Reserva Hidrológica de Rio Claro-SABESP, trail to Campos de Boracéia, 850 m, 45°54′S, 22°36′W, 25 Feb 1991 (sterile), L. G. Clark & M. Morel 829 (holotype: SP; isotypes: ISC, MBM, MO, NY, RB, SJRP, US-n.v.). Epitype, designated here: Brazil. Minas Gerais: Mun. Felício dos Santos, APA Felício, 18° 10′S, 43° 17′W, 1000–1400 m, 8 Oct 2004 (fl), P. L. Viana, N. F. O. Mota, T. E Almeida, A. Salino, H. Bispo, F. M. G. Santos 2229 (BHCB; isoepitypes: IBGE, ISC, RB, US, UWSP). (Figs. 1 and 2)

Aulonemia setosa. A. Rhizomes. B. Section of mid-culm nodal region showing buds. C. Ligular region of foliage leaves. D. Flowering shoots with synflorescences. E. Detail of first order branch with spikelets appressed to main axis of synflorescence. F. First glume. G. Second glume. H. Third glume. I. Fertile floret. J. Gynoecium and androecium. K. Lodicules. L. Caryopsis. (From Viana et al. 2229, BHCB.)

Aulonemia setosa: A. Profusely flowering clump. B–D. Synflorescences. E. Branch complements. F. Apex of sheaths showing fimbriae. G. Ligular region of culm leaf, showing conspicuous auricles (A–D from Viana et al 2229 population; E–G Viana & Leite 3760 population.)
Rhizomes sympodial, pachymorph, short-necked. Culms 2–13 m × 0.5–3.5 cm, clambering with pendent fractiform apices. Internodes 12.5–35(–40) cm long, hollow, thick-walled, terete, slightly ridged above the bud, finely mottled, glabrescent with white velvet adpressed trichomes below the nodal line. Nodes solitary, the nodal line horizontal, glabrous, sometimes with white velvety trichomes on the scars of the culm leaf sheaths; intranodal area 0.6–1.6 × 0.9–2.3 cm, glabrous; supranodal line smooth. Buds broadly elliptic to ovate-lanceolate, 11–18 × 8–11 mm, glabrous, the cataphylls ciliolate towards the apex. Culm leaf sheaths 8–28.5 × 2.8–12 cm, persistent, glabrescent, usually with white velvety trichomes, stramineous abaxially, shiny copper-red abaxially, not mottled, the margins glabrous to ciliate; auricles 5–14 × 1.5–4 mm; fimbriae 3–6 mm long; internal ligule membranous-ciliolate, 0.4–2 mm; external ligule inconspicuous; blades narrowly triangular, erect to reflexed with age, constricted at the base, not pseudopetiolate, tardily deciduous, 3.5–20 × 1.5–3 cm, adaxially pubescent, abaxially sparsely pilose to pubescent, the apex setose. Branch complement with 1 branch, often rebranching at its lower nodes to form branchlets. Foliage leaves 2–18(–23) per complement; sheaths 7.1–13.3 cm long, usually glabrous, sometimes with sparse white velvety trichomes, keeled; margins glabrous, auricles present; fimbriae present or not, 0.3–1.7 cm long, erect, free, rounded, slightly wavy, stramineous; internal ligule membranous to membranous-ciliolate, 1.5–5 mm long, asymmetrical; external ligule membranous, ciliolate, ca. 0.5 mm long; pseudopetioles 1.4–5 mm long, glabrous to sparsely pubescent on both surfaces; blades lanceolate, 6.2–22.5 × 0.8–3.9 cm, glabrous to sparsely pilose on the adaxial surface, sparsely pilose to pubescent, densely papillose abaxially, glaucous with a green stripe along one abaxial margin; the base asymmetrical, rounded; the apex acuminate to setose; margins antrorsely scabrous. Synflorescences 22–72 × 0.3–1.1(–1.6) cm, a spicate panicle; peduncle glabrous; the branches appressed to the main axis. Spikelets 1.5–2.6(–3.2) × 0.1–0.2 cm, linear, terete, consisting of 2–3 glumes, 2–5 fertile florets and 1 apical rudimentary floret; glume I 2.3–4 mm long, 1–3-nerved, glabrous, ciliolate towards the apex, stramineous, awned, the awn 2.0–3.2 mm long; glume II 4.7–5.1 mm long, 5–6-nerved, glabrous, ciliolate towards the apex, stramineous, awned, the awn 3.5 mm; glume III 6.1–7.0 mm long, 7–8-nerved, papillose, ciliate towards the apex, stramineus, rarely mottled, awned, the awn 3.3–3.8 mm long; fertile lemma 8 mm long, 11-nerved, papillose, mottled, awned, the awn 3.5 mm; palea 9 mm long, 8-nerved, awnless, papillose, not mottled. Lodicules 3, disimilar, lanceolate to oval-lanceolate, the apex ciliate. Stamens 3; anthers 5–5.5 mm long, purple to greenish. Ovary ca. 1.3 mm long, linear to ellipsoid, glabrous; stigmas 2, pale. Caryopsis narrowly elliptic to oblong, ca. 4.8 mm, pale to brownish; hilum linear, as long as the caryopsis; embryo pale to brownish, ca. 1/7 the length of the caryopsis.
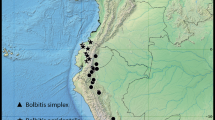
Distribution and ecology.—Aulonemia setosa was a poorly known species with only one population documented in the literature (Londoño & Clark, 2002). An inventory of Brazilian herbaria and additional fieldwork revealed a wider range of geographic distribution for this species. Like most Brazilian Aulonemia species, A. setosa is endemic to the Atlantic Rain Forest biome. It is now known from Espirito Santo, Rio de Janeiro, São Paulo and Minas Gerais states in evergreen and semideciduous forests (Veloso et al., 1991) at elevations of (100–)800–1300 m.
Life cycle .—As suggested by existing specimens, four populations of Aulonemia setosa were sampled in mass flowering events. Two were documented in 1985, one in Espirito Santo (F.O. Zuloaga et al. 2399) and the other in Rio de Janeiro state (G. Martinelli et al. 10635). A single flowering collection was made in 2000 in São Paulo (J. P. Souza et al. 3463) and a large profusely flowering population was recorded by the senior author and collaborators in Minas Gerais from 2004 to 2006 (P. L. Viana et al. 1358, 2229, 3762).
Based on vegetative specimens, which appear to be young individuals collected by the senior author and collaborators in the Quadrilátero Ferrífero region of Minas Gerais state, in 2008 (P. L. Viana et al. 3461, 3760), and on the flowering specimens of 2000, 2004, and 2006, we suggest another probable gregarious flowering event for this species during this period. These data indicate that this species may present a flowering cycle of 15 to 21 years. No other species of Aulonemia has a flowering cycle documented in the literature.
Additional specimens examined. BRAZIL. Espírito Santo: Domingos Martins, selva en cerro del Dr. Kautsky, 600–850 m, 11 May 1985 (fl), F. O. Zuloaga et al. 2399 (BHCB, MO, RB, US, WIS). Minas Gerais: Felício dos Santos, APA Felício, 1000–1400 m, 11 Jun 2006 (fl), P. L. Viana et al. 3762 (BHCB, ISC, RB); same locality, 8 Aug 2004 (fl), P. L. Viana & N. F. O. Mota 1358 (BHCB); Barão de Cocais, RPPN Santuário do Caraça, próx. à RPPN Cachoeira da Capivara, 20°06′49′′S, 43°34′08′′W, 1462 m, 28 May 2008 (st), P. L. Viana & F. S. F. Leite 3760 (BHCB, ISC, RB); Nova Lima, próximo a córrego do Boiadeiro, 20°06′44′′, 43°53′26′′W, 789 m, 23 Aug 2008 (st), P. L. Viana et al. 3461 (BHCB). Rio de Janeiro: Macaé, Pico do Frade de Macaé, floresta higrófila de encosta, 1150–1250 m, 7 Feb 1985 (fl), G. Martinelli et al. 10635 (BHCB, F, RB, US). São Paulo: Caraguatatuba, Parque Estadual da Serra do Mar, Núcleo Caraguatatuba, 23°38′44′′S, 45°40′21′′W, 1100 m. 25 Apr 2000 (fl), J. P. Souza et al. 3463 (CEN); Santos, Área da Santa Casa de Misericórdia de Santos, Trilha 3, 100 m, 7 May 2007 (st), R. J. F. Garcia et al. 3038 (BHCB, PMSP).
Leaf anatomy
Leaf blade in transverse section (Fig. 3D, E)
Leaf blade more or less undulated, the midrib slightly prominent towards the abaxial side. Epidermal cells slightly larger on the adaxial side, protected by a thick cuticle; bulliform cells in groups of two to five oval to elliptic cells, in an adaxial groove or at the same plane of other epidermal cells, reaching about half of the mesophyll width; papillae inconspicuous on the abaxial and adaxial surfaces. Abaxial and adaxial intercostal sclerenchyma fibers present; two adaxial clusters parallel to the bulliform cells, not extending laterally under the adaxial epidermis, the abaxial clusters uni-or biseriate, as wide as the bulliform cells group. Vascular bundles of first order alternated with two to nine of second order ones; first order bundles circular, positioned toward to the epidermis and located abaxially in the mesophyll, surrounded by double sheath, the outer parenchymatic with voluminous globe-shaped cells, the inner one (mestome), composed of fibers with thick and lignified walls, with reduced lumina; second order vascular bundles elliptical, no more than half as wide as those of first order, positioned abaxially in the mesophyll, toward to the epidermis, sheaths incomplete, the parenchyma sheath restricted to two lateral arches, the mestome composed of a few cells. Midrib with a complex vascular system, including a first order vascular bundle surrounded by an expressive group of fibers, which can project laterally under the epidermis on the adaxial surface, and two small vascular bundles disposed on both sides of the midrib; some globose to oblong parenchyma cells surround the bundles (corresponding to the outer sheath). Photosynthetic parenchyma composed of compact arm-cells, forming a biseriate layer located adaxially to the fusoid cells and a uniseriate layer on the abaxial side; the row directly beneath the epidermis (at adaxial side of the leaf) composed of more elongated cells (when compared to the remaining arm-cells) that are more or less isodiametric; fusoid cells 1/3 to 1/2 the width of the mesophyll, elliptic to trapezoid.

Leaf anatomy of Aulonemia setosa. A–C. Epidermis in surface view. A, B. Details of the adaxial leaf surface, outside and inside the marginal stripe region, respectively. C. Abaxial surface with stomata. D–E. Leaf in transverse section, midrib region shown in D. F–G. Deep paradermal section showing vascular system and fusoid cells. Abbreviations: bc=bulliform cell; cz=costal zone; fc=fusoid cells; if=intercostal fibers; iz=intercostal zone; pk=prickle; st=stoma; 1st=first order vascular bundle; 2nd=second order vascular bundle. (From Viana & Leite 3760, BHCB.)
Leaf epidermis in surface view (Figs. 3A–C and 4)
Adaxial surface.—Adaxial surface slightly undulated, ridged on the costal zones and faintly grooved on the bulliform cell region. Long cells with slightly sinuous anticlinal walls alternating with solitary short corky cells. Short barbed to unarmed prickles occasional, present on the entire adaxial surface, except on the bulliform cell rows; bulliform cells hexagonal to quadrangular in the central rows and slightly elongated in the lateral rows; bicellular microhairs occasional, with basal and distal cells approximately equal in length, with the basal one more inflated and apparently thicker-walled (Fig. 4c, d); unicellular macrohairs absent; papillae usually absent, but small globose papillae can be rarely found in a single row at long cells, and a group of four to six elongated appressed papillae overarches the stomata, when they are present; stomata rarely present, restricted to the marginal stripe region.

Leaf surface of Aulonemia setosa leaf under SEM. A–B. Abaxial surface at marginal stripe region. C. Adaxial surface at marginal stripe region. D–E. Abaxial surface outside marginal stripe region. F. Adaxial surface outside the marginal stripe region. Abbreviations: bc=bulliform cells; bm=bicellular microhair; cz=costal zone; iz=intercostal zone; pk=prickle; st=stoma, um=unicellular macrohair. (From Viana & Leite 3760, BHCB.)
Abaxial surface.—Abaxial surface more or less undulated, slightly ridged on the costal zones and on the region opposite to the bulliform cells, on the intercostal zone. Long and short cells in a pattern similar to the adaxial surface, except the interstomatic long cells, which have slightly sinuous anticlinal walls and are alternated with solitary stoma. Short barbed to unarmed prickles present on the entire abaxial surface, except on the interstomatic cell rows, as well as bicellular microhairs which are similar to those of abaxial surface, and unicellular macrohairs with apparently unspecialized cells associated with their bases; papillae abundant, found as a single row of globose papillae on the long cells at the costal zone and at the region opposite to the bulliform cells, one or two rows of larger globose or compound papillae on the interstomatic cells, and a dense group of compound papillae overarching the stomata; stomata abundant, alternate with long interstomatic cells in groups of three to five adjacent rows. Marginal stripe not clearly differentiated from the rest of abaxial leaf surface, except by the presence of a small amount of epicuticular wax.
Discussion
Aulonemia setosa is closely related to A. setigera, known from Rio de Janeiro and Bahia states of Brazil, with which it shares a set of features not found in the core species of Aulonemia. Both species have culm and foliage leaf blades with setose apices, lending a graceful aspect to the plant. Both species also have strong thick-walled culms up to 3 cm thick that are usually covered by white velvety appressed trichomes near the nodal region; leathery culm leaves with shiny copper-red adaxial sheath surfaces and erect blades sometimes becoming reflexed with age; prominent auricles on the culm leaves; and terete, awned spikelets. Aulonemia setosa can be differentiated from A. setigera by its shorter leaves (6.2–23 cm long, vs. 21–38 cm long in A. setigera), shorter fimbriae on foliage leaves (0.3–1.7 cm long in A. setosa and (2.0–)4.0–5.2 cm long in A. setigera), pilose abaxial leaf surface (glabrous in A. setigera) and hollow, thick walled culms (solid or with an inconspicuous lumen up to 1 mm wide in A. setigera). This species is cited as Eremocaulon sp. in Clark (2001). It is evident that the floral characteristics and anatomical features of A. setosa fit within Aulonemia, according to the circumscription adopted here (McClure, 1973; Judziewicz et al., 1999; Table I).
Most anatomical features of A. setosa agree with those found in the Arthrostylidiinae (Judziewicz et al., 1999; Soderstrom & Ellis, 1987). The presence of intercostal sclerenchyma fibers, the conspicuous green stripe on the abaxial margin, and the absence of stomata on the entire adaxial surface is a pattern found in almost all of the known Brazilian Aulonemia species (P. L. Viana, unpubl. data), but not in the core Guaduinae genera, including Eremocaulon. On the other hand, the complex vascular system in the midrib of foliage leaf blades in A. setosa is a character not previously recorded in the Arthrostylidiinae. It was considered a typical Guaduineae feature (Soderstrom & Ellis 1987) but does not appear to be a consistent distinction between the two subtribes.
The importance of foliage leaf anatomical features in bamboo taxonomy is exemplified in a recent phylogeny of the Guaduinae genus Otatea (Ruiz-Sanchez et al., 2008), where two Mexican species included in the analysis described in Aulonemia (A. clarkiae Davidse & R.W. Pohl and A. fulgor Soderstr.) were found nested within the Guaduinae clade. Although these species possess most diagnostic morphological features of Aulonemia (McClure, 1973; Soderstrom, 1988; Davidse & Pohl, 1992; Judziewicz et al., 1999), their anatomical characteristics are typically Guaduinae-like, without the abaxial marginal green stripe on the foliage leaves, with stomata present on both foliage leaf blades over the entire surface, and an absence of intercostal sclerenchyma.
The number of second order vascular bundles between any two first order bundles, which is often used as a character of taxonomic value, is not uniform along the leaf blade of A. setosa. The pattern of the ratio of first and second order vascular bundles in different portions of the leaf should be better studied and illustrated to assess the taxonomic value of this character.
Although there is enough morphological and anatomical data to accept the combination Aulonemia setosa (Londoño & Clark) P.L. Viana & Filg., phylogenetic analyses including more species will hopefully elucidate the relationships between the Guaduinae and Arthrostylidiinae clades and also the circumscription of some critical genera, such as Aulonemia. Intensive fieldwork is still a critical need because bamboos are poorly represented in the collections of most herbaria in Brazil (and elsewhere). This results in an inaccurate representation of the distributional ranges of the species, provides incomplete morphological characterizations, and underestimates local plant diversity.
Literature Cited
Bukatsch, F. 1972. Bemerkungen zur doppelfärbung Astrablau-Safranin. Mikrokosmos 61: 255.
Clark, L. G. 2001. Eremocaulon. Pp. 37–38. In: H. M. Longhi-Wagner, V. Bittrich, M. G. L. Wanderley, G. J. Shephered (eds.), Flora Fanerogâmica de São Paulo, vol 1: Poaceae. Editora Hucitec. São Paulo.
Davidse, G. & W. P. Pohl. 1992. New taxa and nomenclatural combinations of Mesoamerican grasses (Poaceae). Novon 2: 81–110.
Ellis, R. P. 1979. A procedure for standartizing comparative leaf anatomy in the Poaceae. II. The epidermis as seen in surface view. Bothalia 12: 641–672.
———. 1976. A procedure for standardizing comparative leaf anatomy in the Poaceae. I. The leaf blade as viewed in transverse section. Bothalia 12: 65–109.
Fuchs, C. H. 1963. Fuchsin staining with NaOH clearing for lignified elements of whole plants or plant organs. Stain Technology, 38: 141–144.
Judziewicz, E. J. & R. Riina. 2005. Aulonemia dinirensis (Poaceae: Bambusoideae: Bambuseae), a new dwarf Venezuelan species from the easternmost Andean páramos. Bamboo Science and Culture 19: 11–15.
———, L. G. Clark, X. Londono & M. J. Stern. 1999. American bamboos. Smithsonian Institution, Washington.
Jensen, W. A. 1962. Botanical histochemistry: principles and practice. W.H. Freeman, San Francisco.
Johansen, D. A. 1940. Plant microtechnique. McGraw Book Co., New York.
Londoño, X. & L. G. Clark. 2002. A revision of the Brazilian bamboo genus Eremocaulon (Poaceae: Bambuseae: Guaduinae). Systematic Botany 27: 703–721.
McClure, F. A. 1973. Genera of bamboos native to the New World (Gramineae: Bambusoideae). Smithsonian Contributions to Botany 9: 1–148.
Ruiz-Sanchez, E., V. Sosa & M. T. Mejía-Saules. 2008. Phylogenetics of Otatea inferred from morphology and chloroplast DNA sequence data, and recircumscription of Guaduinae (Poaceae: Bambusoideae). Systematic Botany 33(2): 277–283.
Soderstrom, T. R. 1988. Aulonemia fulgor (Poaceae: Bambusoideae), a new species from Mexico. Brittonia 40: 22–31.
——— & R. P. Ellis. 1987. The position of bamboo genera and allies in a system of grass classification. Pp. 21–30 In: T. R. Soderstrom, M. E. Barkworth, C. S. Campbell, and K.W. Hilu (eds.), Grass Systematics and Evolution. Smithsonian Institution Press, Washington.
Veloso, H. P., A. L. R. Rangel Filho & J. C. A. Lima. 1991. Classificação da vegetação brasileira, adaptada a um sistema universal. IBGE, Rio de Janeiro.
Zhang, W. & L. G. Clark. 2000. Phylogeny and classification of the Bambusoideae (Poaceae). Pp. 53–42. In: S. W. L. Jacobs & J. E. Everett (eds.), Grasses: systematics and evolution. CSIRO Publishing, Collingwood.
Acknowledgments
P. L. Viana thanks FAPEMIG and CAPES for support in the Doctoral program. E. Paiva is supported by a research grant from CNPq. We thank Dr. Emmet Judziewicz, Dr. Dalva G. Ribeiro, Rosy M. Isaías, Ana P. S. Gonçalves and João R. Stehmann for valuable suggestions on the manuscript. We also thank Myrian M. Duarte for the line drawing. The reviewers Dr. Robert Soreng and Dr. Lynn Clark improved substantially the final version of this paper. Heitor Bispo, Nara F. O. Mota and Felipe S. F. Leite (Miguelão Biodiversity Insights) for their help during fieldwork.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article
Cite this article
Viana, P.L., Filgueiras, T.S. & Paiva, E.A.S. A new combination in Aulonemia (Poaceae: Bambusoideae: Bambuseae) based on floral analysis, anatomical features, and distribution. Brittonia 63, 102–112 (2011). https://doi.org/10.1007/s12228-010-9138-0
Published:
Issue Date:
DOI: https://doi.org/10.1007/s12228-010-9138-0