
Abstract
Earlier observations that plant clonality, i.e., production of potentially independent offspring by vegetative growth, increase in importance in cold climates such as in arctic and alpine regions, have been recently questioned. However, lack of data obtained using a comparable methodology throughout different regions limit such comparisons. Here we present a classification of clonal growth forms for vascular plants from East Ladakh (an arid mountain range in NW Himalaya, India), and assess the relationship of these forms with multiple environmental gradients. Based on field assessment of clonality in 540 species we distinguished 20 growth forms, which were then grouped into four broader space occupancy strategies. Occurrence in communities and relationship with environmental characteristics and altitude were analyzed using multivariate methods. The most abundant growth form was represented by non-clonal perennial species with a pleiocorm having short branches, prevailing in steppes, Caragana shrubs and screes. The most abundant clonal species were those with very short epigeogenous rhizomes, such as turf graminoids prevailing in wet Kobresia grasslands. Two principal environmental gradients, together with several abiotic variables, affected space occupancy strategies: moisture and altitude. Non-spreading integrators prevailed on shaded rocky slopes, non-spreading splitters in wet grasslands and spreading splitters at the wettest sites. Spreading integrators were the least frequent strategy predominantly occurring at the most elevated sites. Because relevance of clonality decreased with altitude and different communities host different sets of clonal growth strategies, comparison with other cold climate regions should take multiple environmental gradients into account.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Plant nomenclature Klimeš and Dickoré (2006)
Introduction
The ability of plants to grow clonally, i.e., to produce potentially independent offspring, is considered to be important especially in stressful environments such as cold regions of high mountains and the Arctic (Bliss 1971; Billings 1974; Callaghan et al. 1992; Jónsdóttir et al. 1996; Klimeš et al. 1997; Körner 1999a; Evette et al. 2009). However, preliminary analyses show that this relationship is not as universal as expected (Klimeš 2003; Klimeš and Doležal, unpubl.). For further understanding of the relevance of clonality in cold regions, comparable data obtained with standardized methodologies are needed. This requires the use of a general classification scheme of clonal versus non-clonal growth forms.
The classification of plant species into easily distinguishable growth forms has a long tradition in botany, dating back to von Humboldt (1806) and his attempt to describe physiognomic plant types in taxonomically unknown vegetation of South America. The idea of a common ecological function of similar gross morphologies was proposed by Grisebach (1872), Drude (1887), Raunkiaer (1907), and Warming (1923) in their classifications focusing on different environmental factors, e.g., climate or soil. Although other growth form classifications have been developed (Du Rietz 1931; Gimingham 1951; Hejný 1960; Lukasiewicz 1962; Den Hartog and Segal 1964; Serebrjakov 1964; Parsons 1976; Hallé et al. 1978; Barkman 1988; Halloy 1990; Kästner and Karrer 1995; von Lampe 1999; Krumbiegel 1998, 1999), few are generally, or at least repeatedly, employed in ecological studies. Despite these there are indeed different ecomorphological plant traits that are widely used in ecological studies to understand the functionality of plants and ecosystems. These are, e.g., the life forms delimited by Raunkiaer according to the highest buds on plants surviving unfavourable seasons (Raunkiaer 1910), phalanx versus guerrilla growth forms distinguishing two contrasting space occupancy strategies of clonal plants (Lovett Doust 1981), and splitters versus integrators distinguishing strategies of clonal plants either reallocating or conserving resources (Jónsdóttir and Watson 1997). However, other ecomorphological traits have a potential to become similarly important, namely clonal growth organs (e.g., stolons, rhizomes or roots), the functionality of which is often studied in comparative experimental studies (see van Groenendael and de Kroon 1990; de Kroon and van Groenendael 1997), although their importance at the community level remains rarely investigated (but see Tamm et al. 2002; Halassy et al. 2005, Sosnová et al. 2010).
Although sometimes difficult to standardize across different taxa, a limited set of morphological characters relevant to clonality can be established and can overcome the contingency of the growth form approach of classifications in particular studies and ecosystems (Klimeš et al. 1997; Klimeš and Klimešová 1999, 2000, 2005; Klimešová and Klimeš 2008). In this approach it is first essential to identify which organ bears buds for shoot iteration (sensu Hallé et al. 1978), which organ provides connections between ramets, and where these organs are placed. Then it is necessary to assess whether adventitious roots are formed and whether specialized storage organs are present. These principles were used for the classification of clonal growth organs for plants of Central Europe with the notion that a similar classification is applicable with some minor ad-hoc modifications in other regions (Klimešová and de Bello 2009).
In this study we develop and apply such an ecomorphological classification for 540 species from the high mountain range in Eastern Ladakh, i.e., in the dry West Himalayas, to test the suitability of principles of the clonal growth organ classification in a floristic region of high altitude. We were aware of other growth form classifications aiming to describe clonal propagation in the alpine zone (Hess 1909; Hartmann 1957; Rauh 1987; Halloy 1990; Körner 1999a), however we considered the effort to include both the above- and below-ground architecture of plants proposed in most of them as too complicated for the purpose of describing clonal morphological traits and assessing the relevance of space occupancy strategies in cold regions. We are aware, however, that classifications resulting in 20 categories can limit generalizations across regions. Therefore, we grouped the morphological categories according to their capacity to spread (spreading and non-spreading) and the longevity of their connection (integrator or splitter). The resulting four basic clonal strategies in space occupancy are delimited according to the spectrum of traits found in the area.
Eastern Ladakh has a harsh climate, where plants are subjected to multiple stresses such as low precipitation (50–100 mm per year), extreme diurnal temperature fluctuations, strong winds, solifluction at higher and salinity at lower altitudes (Stewart 1916–1917). Habitat types and plant communities are relatively sharply delimited by differences in abiotic factors (mostly related to altitude, substrate and moisture availability) and plant species composition (Dvorský et al., 2011). Environmental selection pressure is strong and is presumably not masked by biotic interactions (Körner 1999b). Due to the remoteness of E Ladakh, the local flora is little affected by plant invasions even though human activities, such as grazing of domestic animals and collection of fire wood, have influenced the vegetation substantially despite low human population.
Ladakh in the NW Himalayas is often termed a “cold desert” (Chowdhery and Rao 1990). Although the flora is species-poor (Dickoré and Miehe 2002), it includes almost the entire spectrum of life and growth forms (Klimeš 2003) and most of the important families of the Holarctic flora, except for exclusively woody taxa. Growth form spectra and their altitudinal distribution in the study region were determined in four broad geographical sub-areas, between 750 and 3,100 km2 in size. The study area comprises one of the most prominent altitudinal gradients inhabited by vascular plants in the world, ranging from 3,550 to 6,030 m above sea level.
Our study had three aims: i) to classify clonal growth forms of the study area on a morphological basis and to combine them into four basic clonal strategies of space occupancy, ii) to assess clonal growth spectra of plant communities, iii) to evaluate their distribution along environmental gradients, in particular the elevation gradient, i.e., to test whether morphologically delimited clonal growth forms have a functional relevancy. To correlate clonal growth form spectra and environmental conditions, both at the regional flora and the plant community levels, we used two different approaches, i) cross-species analysis and ii) relative species abundance within a vegetation type. Studies of trait responses based on equal importance of species may yield results different from those considering species abundance. Because growth form traits are correlated with commonness/rarity within a stand (Pakeman and Quested 2007), trait–habitat relationships based only on species presence/absence data tend to overestimate rare forms and underestimate common forms (Grime et al. 1988; Grime 2006; Pakeman et al. 2008).
Material and Methods
Study Area
The study area comprises the eastern part of Ladakh (Jammu and Kashmir State, India), situated between 32°41.5′–33°59.7′ N and 77°47.0′–78°33.4′ E, and covering 6,912 km2. Eastern Ladakh extends between altitudes of 3,550 and 6,670 m (Lungser Kangri Peak), along both sides of the Upper Indus River between the Eastern Karakorum range in the north and the main range of the Western Himalayas to the south, and includes the south-western most extension of the Tibetan Plateau with three high-altitude internal drainage basins. The study area can be further divided into four districts: Pangong, Stot, Samad Rokchen, and Tso Morari (Klimeš 2008), 760 to 3,080 km2 in size (Fig. 1; Table 1). Due to the large differences in species composition these can be considered as distinct bioregions; only 197 species out of the 540 found in Eastern Ladakh were found in all four areas (Klimeš 2008). The whole area is generally arid with vascular plants growing from 3,550 m (bottom of Indus Valley) up to 6,060 m a.s.l. (snowline).

Map of Ladakh, NW India, with the four study regions (Pangong, Stot, Samad Rokchen, Tso Morari)
Eastern Ladakh is rarely affected by monsoonal precipitation, which fails to cross the main Himalayan range (Bhattacharyya 1989). Mean annual precipitation decreases eastward along the Indus Valley from 115 mm at Leh (3,514 m, 34°09′ N, 77°34′ E, about 50 km NW of the study region) to 54 mm at Gar in SW Tibet (4,232 m, 32°07′ N, 80°04′ E, ca. 160 km SE of the study region). Evaporation exceeds precipitation at lower and middle elevations. The few climatological data available for higher elevations of Ladakh suggest that the mean annual temperature is around −8.2°C; the mean monthly temperature rises above 0°C from June to August only (Miehe et al. 2001). Climatic measurements in the Tso Morari region in 2003 showed that the soil temperature at the upper distributional limit of vascular plants at 6,030 m a.s.l. remained above zero only between mid-July and the beginning of September (Klimeš and Doležal 2010). The substrate ranges from siliceous rocks (Precambrian granites, Tso Morari gneiss) to calcareous or saline sediments.
A general description of the vegetation of this area is provided by Hartmann (1983, 1984, 1987, 1990, 1995, 1997, 1999), Rawat and Adhikari (2005), Kala and Mathur (2002), and Klimeš (2003). Due to the low precipitation, steppe vegetation dominated by several Stipa and Artemisia species prevails in the whole region up to 5300 m in altitude (Fig. 2). Other frequent species include Oxytropis microphylla, Ajania fruticulosa and Ptilotrichum canescens. Only small areas at the relatively lowest elevations of the Indus Valley are occupied by semi-desert. A narrow belt of alpine grasslands occurs along water streams running from melting glaciers, where Kobresia pygmaea and Carex pseudofoetida usually dominate. Alpine and subnival vegetation, characterized by Astragalus confertus, Thylacospermum caespitosum and Potentilla pamirica has developed above 5,300 to 5,700 m. Vascular plants occur regularly up to 5,900 m, locally up to 6,060 m above sea level. Salt marshes and wetlands, which occupy relatively large areas of flat and broad valley bottoms in the whole region, represent extrazonal vegetation (Fig. 2). Synanthropic vegetation is developed on eutrophicated soil in the vicinity of stables of domestic animals and near villages up to 5,400 m. Steppes and alpine grasslands are grazed by sheep, goats and yaks up to 5,600 m (Jina 1995; Holzner and Kriechbaum 1998).

Eight major vegetation types in East Ladakh. 1 – salt marshes; 2 – animal resting places; 3 – water bodies; 4 – shrubs; 5 – steppes; 6 – wet meadows; 7 – screes; 8 – subnival zone
Growth Form Categorization
The methodology to categorize species into different growth forms is derived from the approach used to categorize the Central European flora as proposed by Klimeš et al. (1997). Although the basic idea of this classification had already been implicitly used to assess the altitudinal distribution of life forms and clonality in the region (Klimeš 2003) and to test the hypothesis whether split or integrated clones prevail in the harsh conditions of the region (Klimeš 2008), a general and more detailed description of this classification is necessary to enable comparative studies in other regions.
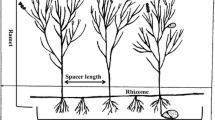
The approach is based on the assessment of a set of morphological plant traits related to clonal growth reproduction, i.e., persistence at a spot, lateral spread and clonal propagation (Fig. 3). These traits are hierarchically organized to form a hierarchical trait-based identification key (Appendix 1), which can be applied and compared in other regions of the world by experts and, after some basic training, also by non-experts. The advantage of producing many distinguishable groups with this approach is that i) this can be generalized across regions and ii) possible subsequent grouping into a more reduced number of groups can be based on the same hierarchical structure in different regions, and therefore allows for meaningful comparisons.

Classification scheme of 20 growth forms of vascular plants in East Ladakh (NW Himalaya, India). Growth forms are named after representative species
The classification is based on the following questions: which organ bears buds for iteration of shoots, which organ provides connections between offspring shoots, if this organ is short or long, if a plant is able to form adventitious roots, and if there are special storage organs. There are three exceptions: annuals and biennials, woody plants and cushion plants are assessed according to their whole morphology. Short-lived monocarpic plants are non-clonal, trees and big shrubs are very rare in the area and cushion plants are a traditionally distinguished growth form in the alpine regions. Traits not taken into account in the classification included, for example, plant size, number and spatial distribution of shoots, distribution of leaves along a shoot, and lifespan of leaves. We also avoided a delimitation of categories primarily according to taxonomy. Non-clonal plants are represented by a particularly heterogeneous group, including short-lived monocarpic species (annuals and biennials) and trees, cushion plants with a tap-root and perennial herbs lacking adventive roots and buds. Although their growth forms differ substantially, they form homogeneous group from point of view of clonality: they are not able to form offsprings by vegetative growth.
The resulting 20 categories (with the exclusion of annual plants) were further grouped into clonal strategies of space occupancy (Fig. 4) according to two characteristics: rate of lateral spread (spreading – more than 10 cm per year; non-spreading – less than 10 cm per year) and persistence of connections between ramets (splitters – plants producing adventitious roots with main root decaying; integrators – plants not producing adventitious roots and/or with perennial main root). Although the division into integrators and splitters was used in a preceding work from this area (Klimeš 2008), here we simplified the scheme by lumping together all plants belonging to a certain clonal growth form category and including non-clonal perennials with a perennial main root, as this category is the most plastic and potential clonality may occur in plants occurring in wetter habitats or after disturbance of below-ground plants.
Plants for morphological description, optimally several individuals of each species, were excavated with below-ground organs and dried between paper sheets for future comparison and identification. Plant material is stored in two herbaria, PRA (Institute of Botany, Academy of Sciences of the Czech Republic) and BSD (Botanical Survey of India, Northern Circle Herbarium, Dehradun).
Floristic and Vegetation Data
The field data were collected on eight expeditions lasting four to seven weeks each, from 1998 to 2003, in 2005 and in 2008. 540 species were recorded at a total of 1,282 localities (each approx. 1 ha in size). The localities were non-randomly distributed, as large areas are devoid of vascular plants (very unstable slopes, glaciers, lakes, extremely high elevations). Attempts were made to cover all elevations and habitats in all parts of the delimited areas. Depending on habitat diversity, accessibility, species richness and elevation, plants at each locality were searched for about 30 minutes to one day. The final number of records was 27,018 and these were used to calculate the mean and maximum elevation of each species in the four study regions of E Ladakh. The elevation (in meters above sea level) was estimated with an altimeter (Thommen, Switzerland). The number of vascular plant species recorded in the four study areas ranged from 279 to 486, with a total of 540 (Table 1).
Habitat preferences of individual growth form categories were analyzed using vegetation records from 369 plots (each 100 m2), sampled in a stratified design to cover visually assessed differences in vegetation types over the whole area (Fig. 2). The samples were non-randomly distributed, as large areas are devoid of vascular plants. In each plot, species composition and altitude were recorded and cover of individual species estimated on a semi-quantitative scale. Five environmental variables, considered to be the most important for habitat differentiation, were assessed in each plot using the following scales:
-
(1)
stability of the soil/substrate surface (the inverse of disturbance frequency): 1 – unstable (screes, dunes, solifluction soils), 2 – partly stable (grasslands, steppes), 3 – stable (rocky crevices, Kobresia pygmaea mats);
-
(2)
light availability: 1 – shaded (gorges, shaded rocky crevices, walls of stream banks), 2 – partially shaded (dense vegetation cover), 3 – full light (sparse vegetation cover);
-
(3)
soil moisture: 1 – dry (substrate usually without visible traces of water), 2 – mesic, 3 – wet (water level regularly but transiently above soil surface), 4 – permanent surface water;
-
(4)
nutrient availability: 1 – low (semi-deserts, steppes), 2 – medium, 3 – high (stables, animal resting places);
-
(5)
soil salinity: 1 – no salt deposits on soil surface, 2 – salt deposits scarce, 3 – salts forming a continuous crust.
Data Analysis
When a species was classified in several categories of clonal growth form its occurrence in these categories was expressed proportionally to the number of categories, so that the final value per species was 1.
Whether the mean and maximum elevations of individual growth form categories differ between the four study areas was tested using ANOVA followed by Tukey’s pairwise comparisons (Table 1).
The habitat preferences of individual growth forms were assessed using both unweighted data and data weighted by abundance. We first characterized the samples by the averages of trait values of the constituent species (as traits are categorical variables, the value represents the relative proportion of a species belonging to the given growth form, weighted or not by their cover), and then we compared the growth form spectra between major vegetation types derived from Two-way Indicator Species Analysis (TWINSPAN). This cluster analysis starts with primary ordination of sites along the first axis of correspondence analysis (CA; see Hill and Šmilauer 2005 for details) and the sites are then divided into two clusters by splitting the first CA axis near its middle. Site classification is refined using a discriminant function that emphasizes species preferential to one or the other half of the dichotomy. We repeated this process to classify the vegetation of the 369 plots into eight ecologically interpretable groups (animal resting places, salt marshes, semi-deserts and steppes, shrublands, alpine screes, wet grasslands, water bodies and the subnival zone, Fig. 2).
Principal Component Analysis (PCA) was used to assess relationships between the growth form categories. Their relations to the eight habitat types was also assessed by their passive projection to the PCA ordination plane using pie graphs depicting the weighted proportion of the four types of space occupancy strategies (and annuals) in an average community. We further used a direct ordination analysis, Redundancy Analysis (RDA, ter Braak and Šmilauer 1998), to relate growth form spectra to recorded environmental variables. The weighted proportion of individual growth forms in each relevé was used as response variables, and several recorded abiotic factors, altitude, substrate texture, soil stability, salinity, shading, soil moisture and fertility, as explanatory variables. Furthermore, an attempt was made to relate vegetation physiognomy (relative shrub, herb, and moss layer cover) and species composition characteristics to growth form spectra by their passive projection to the RDA ordination plane. Species composition was expressed by the first two DCA axes obtained from the indirect ordination of vegetation relevés. In RDA, we first reduced the number of predictors within the group of abiotic variables using a forward selection procedure, and then we performed several RDAs to assess their relative effects on growth form composition using a variance partitioning procedure, in which case the factors not used as predictors were defined as co-variables to remove their effects and obtain the net effect of an individual factor (using this approach we constructed tests analogous to the testing of particular terms in ANOVA models but for multivariate data). Monte Carlo permutation tests (999 permutations) were used to assess the significance of the relationships found in the multivariate analyses. The ordination methods and permutation tests were conducted using CANOCO (ter Braak and Šmilauer 1998). The distribution of abundant growth forms along the elevational gradient was visualized with response curves fitted using generalized additive models with a Poisson distribution and a stepwise selection of alternative models with increasing complexity up to 3 degree of freedom using the Akaike Information Criterion (see Lepš and Šmilauer 2003: 139).
Results
Implementation of the Classification Scheme
Based on a study of their morphology, 540 species of the East Ladakh flora were classified into 20 clonal growth forms (Fig. 3) and these categories were further grouped into four space occupancy strategies (Fig. 4). Some clonal growth forms, although morphologically distinct, were represented by only one or two species and several species were classified into more than one category (Figs. 5 and 6). With the exception of type 18 (“Potamogeton amblyphyllus”, possessing turions) absent from one of the subregions (Table 1), and type 19 (represented only by Potamogeton perfoliatus, which propagates by stem fragments) absent from all subregions, all other clonal growth forms were found in all studied subregions. Potamogeton perfoliatus was found only in river beds of the Indus valley and was outside of the altitudinal ranges considered in this study, however we keep the category in our classification for possible study of an expanded geographical area. Classification of space occupancy strategies revealed that a quarter of the species are non-spreading integrators (25.8%). Non-spreading splitters are slightly less numerous (22.3%), followed by spreading splitters (14.8%), and spreading integrators (9.5% of the flora) (Fig. 4).

Various forms of clonal structures categorized according to their capacity to spread and the longevity of their connections. The origin of the clonal structure and examples of species are given for each of the four categories (for more details, see Fig. 3)

Representative species of selected growth forms. 1 – Microula tibetica; 2 – Arnebia euchroma; 3 – Saussurea gnaphalodes; 4 – Thylacospermum caespitosum; 5 – Parrya nudicaulis; 6 – Ptilotrichum canescens; 7 – Halerpestes sarmentosa; 9 – Festuca kashmiriana; 10 – Cremanthodium ellisii; 11 – Biebersteinia odora; 12 – Leontopodium ochroleucum; 13 – Poa tibetica

Herbarium specimens of species representative of selected growth form categories. 1 – Microula tibetica; 2 – Arnebia euchroma; 3 – Saussurea gnaphalodes; 4 – Thylacospermum caespitosum; 5 – Parrya nudicaulis; 6 – Ptilotrichum canescens; 7 – Halerpestes sarmentosa; 8 – Thymus linearis; 9 – Festuca kashmiriana; 10 – Cremanthodium ellisii; 11 – Biebersteinia odora; 12 – Leontopodium ochroleucum; 13 – Poa tibetica; 15 – Lloydia serotina; 16 – Bistorta vivipara
Clonal Growth Spectra in the Flora and Plant Communities
The analysis of the proportions of the different growth forms in the four study regions showed that non-clonal plants generally prevail (58.7% of the species, attaining a weighted average vegetation cover of 76% per plot) over clonal plants, and their proportion increased with altitude (Table 1). The most abundant non-clonal perennial plants were those with a tap root, a pleiocorm having short branches (“Arnebia euchroma” type), representing 22.9% of the vascular plants, prevailing in steppes (37.9% weighted cover), Caragana shrubs (28.1%) and screes (31.4%) (Table 2). Non-clonal perennial plants with a pleiocorm having long branches (“Saussurea gnaphalodes” type, 8.3% of the species) were significantly more abundant in alpine screes (25.9% cover). Annuals and biennials (“Microula tibetica” type, 24.5% species) prevailed in animal resting places (higher soil N and disturbances, 58% cover), and were also abundant in salt marshes (15.9%). Compact cushion plants (“Thylacospermum caespitosum” type, 3%) occurred exclusively in the subnival zone (12.5% cover). The most abundant clonal plants were those with very short epigeogenous rhizomes (“Festuca kashmiriana” type, 13.4% species) prevailing in moist Kobresia grasslands (32.8%), but also in steppes (25.6%) and shrubland (23.8%). Clonal plants with long (>10 cm) below-ground hypogeogenous rhizomes (“Poa tibetica” type, 7.4% of the species) prevailed in salt marshes (36%), representing extrazonal vegetation in valley basins, and were the least abundant in screes (5%) and steppes (8.6%). All other types occurred in less than 6% of the E Ladakh vascular plants.
Growth form categories showed similar habitat preferences when unweighted data and data weighted by abundance were considered (Table 2). There was a close similarity between the ranking of growth types across species and across data weighted by cover. Differences in the percentage of species belonging to a particular growth type between the weighted and unweighted data were found for plants with very short epigeogenous rhizomes (“Festuca kashmiriana” type) and plants with long hypogeogenous rhizomes (“Poa tibetica” type), i.e., clonal dominants in steppes and salt marshes, respectively, which are overrepresented in the vegetation data. However, plants with a pleiocorm having short branches (“Arnebia euchroma” type) and annual and biennial plants (“Microula tibetica” type) (both non-clonal plants) are relatively underrepresented in the vegetation data, indicating their inability to occupy space and dominate vegetation.
The co-occurrence of individual growth forms is shown in the PCA diagram (Fig. 7), together with passively projected centroids of the eight habitat types. Among growth forms that showed a positive correlation were plants with a pleiocorm having short branches (“Arnebia euchroma” type), rootsprouters (“Ptilotrichum canescens” type) and woody plants (“Juniperus semiglobosa” type) and these were negatively correlated with plants possessing short rhizomes (“Cremanthodium ellisii” and “Leontopodium ochroleucum” types). A positive relationship was found between plants with a pleiocorm having long branches (“Saussurea gnaphalodes” type) and cushion plants (“Thylacospermum caespitosum” type).

Relations between the growth form categories, named after representative species, in a PCA plot with passively projected supplementary variables: eight major vegetation types in E Ladakh and additional information on weighted proportions of annuals and four clonal structures categorized according to their capacity to spread and the longevity of their connections (black – non-spreading integrators, white – non-spreading splitters, grey – spreading splitters, hatched – spreading integrators, dotted – annuals)
The PCA diagram also shows the habitat-specific proportion of clonal structures categorized by their capacity to spread and the longevity of connections (clonal strategies of space occupancy). Of the 540 species recorded in the East Ladakh, the most abundant were non-spreading integrators (25.8%), prevailing in steppes (41.2% weighted cover), alpine screes (40.7%), Caragana shrubs (45.4%), and subnival vegetation (38.4%). The second most abundant were non-spreading splitters (22.3% of the total species number) prevailing in wet grassland (50.2% weighted cover). Spreading splitters (14.8%) dominated in water bodies (79.3%) and salt marshes (43.5%). Least represented were spreading integrators (9.5%) being relatively more abundant in salt marshes (15.9%).
Relationships Among the Growth Form Categories and Environmental Variables
Variation in growth form composition were further explored in the context of the site (abiotic variables used as explanatory variables) and vegetation characteristics (vegetation physiognomy and species composition passively projected into the RDA ordination plane) (Fig. 8). From the pool of 14 abiotic variables, 10 were selected by forward selection in the following order: moisture, altitude, salinity, stability, fertility, silt, shading, 1 cm gravel, rocks, slope, and sand. These explained 28.4% of the total variation in growth form composition (Table 2). The variance partitioning procedure revealed that moisture is the most important determinant of growth form spectra, explaining the most data variation (Table 3), followed by altitude, salinity, stability, fertility and shading, with all other factors explaining less than 1% of variability. The first RDA axis corresponded to the salinity and gravel gradients tightly correlating with the “Poa tibetica” (positively) and “Arnebia euchroma” (negatively) types, i.e., clonal plants with long hypogeogenous rhizomes prevail in more saline sites, whereas non-clonal plants with a tap root, a pleiocorm having short branches, increase in importance on steeper sites with larger gravel proportion. The silt and moisture gradients (corresponding to DCA axis 1 describing variation in species composition from dry semi-deserts and steppes to wet grasslands and water bodies), correlated positively with the proportion of stoloniferous plants (“Halerpestes sarmentosa” type) and plants with short epi- and hypogeogenous rhizomes (“Cremanthodium ellisii” and “Leontopodium ochroleucum” types), and negatively with the proportion of root sprouters (“Ptilotrichum canescens” type) and non-clonal perennial plants with a pleiocorm having short below-ground branches (Arnebia euchroma type). The altitudinal gradient (corresponding to DCA axis 2 describing compositional changes from salt marshes in valley basins towards alpine screes and subnival vegetation at highest elevations) correlated positively with cushion plants (“Thylacospermum caespitosum” type), and plants with a pleiocorm having long branches (“Saussurea gnaphalodes” type), and negatively with annuals and biennials (Microula tibetica type) preferring sandy sites at lower elevations. The RDA diagram also shows the clonal strategies of space occupancy passively projected into the ordination plane (Fig. 8), with non-spreading integrators prevailing in shaded rocky slopes, spreading integrators at the highest elevation, non-spreading splitters in stable grassland along water streams, and spreading splitters in saline and nutrient-rich environments at lower elevations.

Relations between recorded environmental variables and growth form categories in an RDA plot with passively projected supplementary variables: vegetation physiognomy (relative shrub, herb, and moss layer cover), species composition (first two DCA axes), and four basic clonal growth strategies (centroids)
Clonal Growth Spectra Along Altitudinal Gradient
Modelling of both unweighted data and data weighted by abundance with altitude showed similar patterns, therefore only results from the latter approach are presented (Fig. 9). Non-clonal annual and biennial plants (“Microula tibetica” type) were more frequent at lower elevations than non-clonal perennial plants having a tap root and below-ground branches of varying length (“Arnebia euchroma” and “Saussurea gnaphalodes” types) (Fig. 9). At the cold end of the gradient, i.e., at higher elevations, non-clonal cushion plants (“Thylacospermum caespitosum” type) dominated. Most clonal growth forms occurred at middle elevations from 4,500 to 5,500 m a.s.l. (Fig. 9). Clonal plants with long hypogeogenous rhizomes (“Poa tibetica” type) or above-ground stolons (“Halerpestes sarmentosa” type), root sprouters (“Ptilotrichum canescens” type), and woody plants (“Juniperus semiglobosa” type) decline with altitude, while non-clonal perennial plants with a pleicorm and long below-ground branches (“Saussurea gnaphalodes” type), cushion plants (“Thylacospermum caespitosum” type), and clonal plants with short epigeogenous rhizomes (“Cremanthodium ellisii” type) increase with altitude. Annual and biennial plants (“Microula tibetica” type), non-clonal perennial plants with a pleiocorm having short branches (“Arnebia euchroma” type), and plants with very short epigeogenous rhizomes (“Festuca kashmiriana” type) peaked at middle elevations, and decreased towards both lower and higher elevations.

Response curves of selected growth forms for the altitudinal gradient named after typical representatives. Growth form proportions were weighted by species cover within each vegetation type
Discussion
We distinguished 20 clonal growth forms and four space occupancy strategies responding to multiple environmental gradients, mainly moisture and altitude together with several soil variables, in West Himalaya. Overall, in dry and the most elevated communities non-spreading integrators prevailed (i.e., plants with a pleiocorm having short branches, and cushion plants), in wet grasslands non-spreading splitters (plants with short rhizomes) and in saline wetlands spreading splitters prevailed (plants with long hypogeogenous rhizomes). Spreading integrators (plants with a pleiocorm having long branches, and root sprouters) were the least frequent strategies concentrated in places with an unstable substrate (screes and alluvial deposits). These overall results point out that future studies aiming to compare the relevance of clonality in different regions should take multiple environmental gradients into account.
Classification
Due to the high diversity of existing plant forms, inevitably some doubts arise during the process of attributing species to a given growth form. In our case, it was not clear whether below-ground stem branches emerging from the main root in some species (e.g., Ephedra gerardiana, Waldheimia tridactylites, and Thylacospermum caespitosum) could, when disconnected from the main root by disturbance, form adventitious roots and survive independently and therefore show some restricted capacity of clonal growth. Physochlaina praealta, a species forming tubers, seems to be potentially able to form rhizomes in sandy soil. Cushion growth forms, although they may be formed by species that produce adventitious roots and thus might be considered clonal in other regions of the world (Rauh 1939), are mostly represented by non-clonal plants in the region. Data from other regions are necessary to test whether the cushion growth form is more important for plant life in extreme environments than below-ground morphology. Phenotypic plasticity and variability at the population level and in relation to environmental conditions may require further study focusing on individual species and their ecology.
As the studied region was the first to which our methodology was applied outside of Central Europe, we can only make a general comparison with the flora of Central Europe (Klimešová and Klimeš 2008) and some preliminary comparisons with other regions. A remarkable difference when comparing the presented results with Central Europe, is the high proportion of non-clonal plants with a perennial main root in Ladakh, which is in contradiction with general statements about the predominance of clonal plants in cold alpine and arctic regions (Klimeš et al. 1997; Callaghan et al. 1997). Whereas alpine communities of the German Alps and the arctic tundra of central Spitsbergen host about 30% of non-clonal species (Klimešová, unpubl. data), the studied area of Ladakh includes about 70% of non-clonal taxa. Also in the “Carex-Elyna” plant community studied by Hartmann in the Alps (1957) clonal plants prevail, reaching 86.8% out of 228 species in the community. Although the regions differ in precipitation and solar irradiance, another possible reason for these differences can be found in the species pools of these regions, which show a higher proportion of dicot species in Ladakh.
In a previous study (see Klimeš 2003) the low proportion of taxa with long rhizomes was interpreted as caused by substrate instability due to cryoturbation. Cryopedogenic processes and shifting of screes would probably affect clonal plants with long rhizomes (“Poa tibetica” type) more than non-clonal plants with a pleiocorm having long branches (“Saussurea gnaphalodes” type). A similar non-clonal growth form with a perennial main root and long below-ground stems called “Schopftriebe” was described from unstable screes in the Alps by Hess (1909) and resemble those growing in Ladakh (see “Saussurea gnaphalodes” type). Its success is because scree has surface layers that are not very suitable for rooting due to mechanical stress and lack of fine soil particles and thus shoots broken off by shifting of scree are not able to form new roots and subsequently die. A main root, however, with attached perennial stem bases bearing a bud bank growing in deep and more stable strata of scree is able to rebuild new shoots. When the mechanical stress caused by frost and thaw cycles is too high, hardly any adaptation is possible (Jonasson and Callaghan 1992).
In comparison to other regions of the world where clonality is well known, e.g., Central Europe, where the two most common clonal growth forms with hypogeogenous and epigeogenous rhizomes are equally represented (Klimešová and Klimeš 2008), hypogeogenous rhizomes are much less common in Ladakh. Because hypogeogenous rhizomes are the most common at wet sites elsewhere (van Groenendael et al 1996), the prevailing dryness of the Trans-Himalayan environment may be a reason for this difference, and restriction of this type to extra-zonal wetlands confirms this idea.
In contrast to central Europe, Ladakh also has a very low number of species forming adventitious buds on horizontal roots (“Ptilotrichum canescens” type). However, the habitats that they occupy are similar to the habitats where root-sprouters grow in the Alps: glaciofluvial deposits (Stöcklin and Bäumler 1996). Deep homogenous substrates with occasional disturbance by water erosion seem to be very suitable for this clonal growth form.
The high number of annual species in the studied area resembles dry lowland areas (Baitulin 1979), however in Ladakh they are represented by weedy plants at low elevations affected by nutrient enrichment from domestic animals and not ephemeral species that profit from short periods with sufficient water supply. Contrary to lowlands, dry regions have a low proportion of bulbous and tuberous geophytes, which is, however, reported from other mountainous regions as well (Körner 1999a). The factors responsible for this might be frost heaving and cryoturbation typical of nival zones with repeating thaw and freezing cycles.
When grouping the clonal growth forms into four categories of space occupancy strategies we can support earlier analyses (van Groenendael et al. 1996; Jónsdóttir and Watson 1997; Halassy et al. 2005; Klimeš 2008; Sosnová et al. 2010), in which correlations between spreading splitters and moisture and non-spreading integrators and dryness were indicated. In our case moisture was correlated with nutrient and salinity as well, and dryness was correlated with poorly developed soils on slopes and rock. This tendency, observed in a whole range of habitats from wetlands (Sosnová et al. 2010) to semi-deserts (this study) is evolutionarily constrained, as monocots lacking a main root have inherited a splitting strategy and at the same time they generally prefer wetter habitats (van Groenendael et al. 1996). The importance of both elevation and moisture gradients in our analysis indicates that when comparing clonal growth spectra between regions the position on the moisture gradient of the regions should be taken into account.
According to Grace (1993), clonal growth organs differ in function and are therefore filtered by environmental factors into different communities. Our results support this idea, because certain types of clonal growth forms were preferred in certain habitats. Preliminarily, we can propose key functions of some clonal growth types, e.g., amelioration of the environment in cushion plants, searching for water in plants with a perennial main root; anchorage in unstable substrates in plants with a pleiocorm having long branches; fast colonization of sandy and gravel deposits along rivers by horizontal roots with adventitious buds, and space occupancy by long hypogeogenous rhizomes in wetlands. The proposed preferences remain to be tested in other alpine regions of the world. We believe that the clonal growth form identification key and the hierarchical nature of the introduced scheme will certainly allow non-experts to apply these plant functions and make meaningful comparisons between different mountain regions.
References
Baitulin IO (1979) Kornevaia sistema rastenii aridnoi zony Kazakhstana (Root systems of plants of the arid zone in Kazakhstan). Nauka, Alma-Ata
Barkman JJ (1988) New systems of plant growth forms and phenological plant types. In Werger MJA, van der Aart PJM, During HJ, Verhoeven JTA (eds) Plant form and vegetation structure. SPB Academic Publishing, The Hague, pp 9–44
Bhattacharyya A (1989) Vegetation and climate during the last 30000 years in Ladakh. Palaeogeogr Palaeoclimatol Palaeoecol 73:25–38
Billings WD (1974) Adaptations and origins of alpine plants. Arctic Alpine Res 6:129–142
Bliss LC (1971) Arctic and alpine life cycles. Annual Rev Ecol Syst 2:405–438
Callaghan TV, Carlsson BA, Jónsdóttir IS, Svenson BM, Jonasson S (1992) Clonal plants and environmental change – Introduction to the proceedings and summary. Oikos 63: 341–347
Callaghan TV, Jonasson S, Brooker RW (1997) Arctic clonal plants and global change. In de Kroon H, van Groenendael JM (eds) The ecology and evolution of clonal plants. Backhuys Publishers, Leiden, pp 381–403
Chowdhery HJ, Rao RR (1990) Plant life in the Himalayan cold deserts: some adaptive strategies. Bull Bot Surv India 32:43–56
de Kroon H, van Groenendael J (1997) The ecology and evolution of clonal plants. Backhuys Publishers, Leiden
den Hartog C, Segal S (1964) A new classification of water-plant communities. Acta Bot Neerl 13:367–393
Dickoré WB, Miehe G (2002) Cold spots in the highest mountains of the world – diversity patterns and gradients in the flora of the Karakorum. In Körner C, Spehn E (eds) Mountain biodiversity: a global assessment. Parthenon Publishers, Lancaster, pp 129–147
Drude O (1887) Deutschlands Pflanzengeographie. Handb Deutsch Landes- Volksk 4:1–502
Du Rietz GE (1931) Life-forms of terrestrial flowering plants. Acta Phytogeogr Suec 3:1–95
Dvorský M, Doležal J, de Bello F, Klimešová J, Klimeš L (2011) Vegetation types of East Ladakh: species and growth form composition along main environmental gradients. Appl Veg Sci (in press)
Evette A, Bédécarrats A, Bornette G (2009) Environmental constraints influence clonal traits of herbaceous plant communities in an Alpine Massif. Folia Geobot 44:95–108
Gimingham CH (1951) The use of life form and growth form in the analysis of community structure, as illustrated by a comparison of two dune communities. J Ecol 39:396–406
Grace JB (1993) The adaptive significance of clonal reproduction in angiosperms – an aquatic perspective. Aquatic Bot 44:159–180
Grime JP (2006) Trait convergence and trait divergence in herbaceous plant communities: mechanisms and consequences. J Veg Sci 17:255–260
Grime JP, Hodgson JG, Hunt R (1988) Comparative plant ecology. Unwin-Hyman, London
Grisebach A (1872) Die Vegetation der Erde nach ihrer klimatischen Anordnung I, II. W. Engelmann, Leipzig
Halassy M, Campetella G, Canullo R, Mucina R. (2005) Patterns of functional clonal traits and clonal growth modes in contrasting grasslands in the central Apennines, Italy. J Veg Sci 16:29–36
Hallé F, Oldeman RAA, Tomlinson PB (1978) Tropical trees and forests. An architectural analysis. Springer Verlag, New York
Halloy S (1990) A morphological classification of plants, with special reference to the New Zealand alpine flora. J Veg Sci 1:291–304
Hartmann H (1957) Studien über die vegetative Fortpflanzung in den Hochalpen. Bischofberger & Co., Buchdruckerei Untertor, Chur
Hartmann H (1983) Pflanzengesellschaften entlang der Kashmirroute in Ladakh. Jahrb Ver Schutz Bergwelt 48:131–173
Hartmann H (1984) Neue und wenig bekannte Blütenpflanzen aus Ladakh mit einem Nachtrag zur Flora des Karakorum. Candollea 39:507–537
Hartmann H (1987) Pflanzengesellschaften trockener Standorte aus der subalpinen und alpinen Stufe im südlichen und östlichen Ladakh. Candollea 42:277–326
Hartmann H (1990) Pflanzengesellschaften aus der alpinen Stufe des westlichen, südlichen und östlichen Ladakh mit besonderer Berücksichtigung der rasenbildenden Gesellschaften. Candollea 45:525–574
Hartmann H (1995) Beitrag zur Kenntnis der subalpinen Wüsten-Vegetation im Einzugsgebiet des Indus von Ladakh (Indien). Candollea 50:367–410
Hartmann H (1997) Zur Flora und Vegetation der Halbwüsten, Steppen und Rasengesellschaften im südöstlichen Ladakh (Indien). Jahrb Ver Schutz Bergwelt 62:129–188
Hartmann H (1999) Studien zur Flora und Vegetation im östlichen Transhimalaya von Ladakh (Indien). Candollea 45:171–230
Hejný S (1960) Ökologische Charakteristik der Wasser- und Sumpfpflanzen in der slowakischen Tiefebenen (Donau und Theissgebiet). Vydavateľstvo SAV, Bratislava
Hess E (1909) Über die Wuchsformen der alpinen Geröllpflanzen. Arbeit aus dem Botanischen Museum des eidg. Polytechnikum Zürich, Druck von C. Heinrich, Dresden
Hill MO, Šmilauer P (2005) TWINSPAN for Windows version 2.3. Centre for Ecology and Hydrology, Huntingdon and University of South Bohemia, České Budějovice
Holzner W, Kriechbaum M (1998) Man’s impact on the vegetation and landscape in the Inner Himalaya and Tibet. In Elvin M, Ts’ui-Jung L (eds) Sediments of time. Environment and society in Chinese history. Cambridge University Press, Cambridge, pp 53–106
Jina PS (1995) High pasturelands of Ladakh Himalaya. Indus Publishing Company, New Delhi
Jonasson S, Callaghan TV (1992) Root mechanical properties related to disturbed and stressed habitats in the Arctic. New Phytol 122:179–186
Jónsdóttir IS, Watson M (1997) Extensive physiological integration: an adaptive trait in resource-poor environments? In de Kroon H, van Groenendael J (eds) The ecology and evolution of clonal plants. Backhuys Publishers, Leiden, pp 109–136
Jónsdóttir IS, Callaghan TV, Headley AD (1996) Resource dynamics within arctic clonal plants. Ecol Bull 45:53–64
Kala CP, Mathur VB (2002) Patterns of plant species distribution in the Trans-Himalayan region of Ladakh, India. J Veg Sci 13:751–754
Kästner A, Karrer G (1995) Übersicht der Wuchsformtypen als Grundlage für deren Erfassung in der “Flora von Österreich”. Fl Austr Novit 3:1–51
Klimeš L (2003) Life-forms and clonality of vascular plants along an altitudinal gradient in E Ladakh (NW Himalayas). Basic Appl Ecol 4:317–328
Klimeš L (2008) Clonal splitters and integrators in harsh environments of the Trans-Himalaya. Evol Ecol 22:351–367
Klimeš L, Dickoré WB (2006) Flora of Ladakh (Jammu & Kashmir, India). A preliminary checklist. Available at: http://www.butbn.cas.cz/klimes
Klimeš L, Doležal J (2010) An experimental assessment of the upper elevational limit of flowering plants in the Western Himalayas. Ecography 32 (doi:10.1111/j.1600-0587.2009.05967.x)
Klimeš L, Klimešová J (1999) CLO-PLA2 – A database of clonal plants in central Europe. Pl Ecol 141:9–19
Klimeš L, Klimešová J (2000) Plant rarity and the type of clonal growth. Z Ökol Naturschutz 9:43–52
Klimeš L, Klimešová J (2005) Clonal traits. In Knevel IC, Bekker RM, Kunzmann D, Stadler M, Thompson K (eds) The Leda traitbase collecting and measuring standards of life-history traits of the Northwest European flora. EDA Traitbase project, University of Groningen, Community and Conservation Ecology Group, Groningen, pp 66–88
Klimeš L, Klimešová J, Hendriks R, van Groenendael J (1997) Clonal plant architectures: a comparative analysis of form and function. In de Kroon H, van Groenendael J (eds) The ecology and evolution of clonal plants. Backhuys Publishers, Leiden, pp 1–29
Klimešová J, de Bello F (2009) CLO-PLA: the database of clonal and bud bank traits of Central European flora. J Veg Sci 20:511–516
Klimešová J, Klimeš L (2008) Clonal growth diversity and bud banks in the Czech flora: an evaluation using the CLO-PLA3 database. Preslia 80:255–275
Körner Ch (1999a) Alpine plant life. Springer Verlag, Berlin
Körner Ch (1999b) Alpine plants: stressed or adapted? In Press MC, Scholes JD, Barker MG (eds) Physiological plant ecology. Blackwell Science, Oxford, pp 297–311
Krumbiegel A (1998) Growth forms of annual vascular plants in central Europe. Nordic J Bot 18:563–575
Krumbiegel A (1999) Growth forms of biennial and pluriennial vascular plants in central Europe. Nordic J Bot 19:217–226
Lepš J, Šmilauer P. (2003) Multivariate analysis of ecological data using CANOCO. Cambridge University Press, Cambridge
Lovett Doust L (1981) Population dynamics and local specialization in a clonal perennial (Ranunculus repens). 1. The dynamics of ramets in contrasting habitats. J Ecol 69:743–755
Lukasiewicz A (1962) Morfologiczno-pozvojove typy bylin (Morphologic development types of perennials). Poznaňskie Towarzystwo Przyjaciól Nauk, Wydz Mat-Przyr, Prace Komis Biol 27(1):1–398
Miehe G, Winiger M, Böhner J, Zhang YL (2001) The climatic diagram map of High Asia. Purpose and concepts. Erdkunde 55:94–97
Pakeman RJ, Quested HM (2007) Sampling plant functional traits: what proportion of the species need to be measured? Appl Veg Sci 10:91–96
Pakeman RJ, Garnier E, Lavorel S, Ansquer P, Castro H, Cruz P, Doležal J, Eriksson O, Freitas H, Golodets C, Kigel J, Kleyer M, Lepš J, Meier T, Papadimitriou M, Papanastasis VP., Quested H, Quétier F, Rusch G, Sternberg M, Theau J-P, Thébault A, Vile D (2008) Impact of abundance weighting on the response of seed traits to climate and land use. J Ecol 96:355–366
Parsons DJ (1976) Vegetation structure in the mediterranean scrub community of California and Chile. J Ecol 64:435–447
Rauh W (1939) Über polsterförmigen Wuchs. Nova Acta Leopold 7/49:267–508
Rauh W (1987) Tropische Hochgebirgpflanzen. Sitzungsber Heidelberger Akad Wiss Math-Natuwiss Kl 3:105–304
Raunkiaer C (1907) Planterigets Livsformer og deres Betydning for Geografien. Munskgaard, Copenhagen
Raunkiaer C (1910) Statistik der Lebensformen als Grundlage für die biologische Pflanzengeographie. Beih Bot Centralbl 27:171–206
Rawat GS, Adhikari BS (2005) Floristics and distribution of plant communities across moisture and topographic gradients in Tso Kar basin, Changthang plateau, eastern Ladakh. Arctic Antarct Alpine Res 37:539–544
Serebrjakov IG (1964) Zhiznennyje formy vysshikh rastenii ikh izuchenie (Life-forms of higher plants and their study). In Lavrenko EM, Korchagin AA (eds) Field geobotany. Nauka, Moskva, pp 146–205
Sosnová M, van Diggelen R, Klimešová J (2010) Which clonal growth organs are found in various wetland communities? Aquatic Bot (in press)
Stewart RR (1916–1917) The flora of Ladakh. Bull Torrey Bot Club 43:571–588 and 625–650
Stöcklin J, Bäumler E (1996) Seed rain, seedling establishment and clonal growth strategies on a glacier foreland. J Veg Sci 7:45–56
Tamm A, Kull K, Sammul M (2002) Classifying clonal growth forms based on vegetative mobility and ramet longevity: a whole community analysis. Evol Ecol 15:383–401
ter Braak CJF, Šmilauer P (1998) CANOCO reference manual and user’s guide to Canoco for Windows. Software for canonical community ordination (version 4). Centre for Biometry, Wageningen
van Groenendael JM, de Kroon H (1990) Clonal growth in plants: regulation and function. SPB Academic Publishing, The Hague
van Groenendael JM, Klimeš L, Klimešová J, Hendriks RJJ (1996) Comparative ecology of clonal plants. Philos Trans, Ser B 351:1331–1339
von Humboldt A (1806) Ideen zu einer Phisiognomik der Gewächse. Cotta, Stuttgart
von Lampe M (1999) Vorschlag zur Bezeichnung der Innovations- und Überdauerungsorgane bei den terrestrischen Stauden Zentraleuropas. Beitr Biol Pflanzen 71:335–367
Warming E (1923) Ökologiens Grundformer. Kongel Danske Vidensk Selsk Skr, Naturvidensk Math Afd 8(4):120–187
Acknowledgements
We would like to thank Jan W. Jongepier for linguistic improvements and Bernhardt W. Dickoré and two anonymous reviewers for comments on an earlier version of the manuscript. The study was supported by the Institute of Botany Academy of Sciences of the Czech Republic (AV0Z60050516), grant GAAV IAA600050802 and the CNRS PICs 4876 project.
Author information
Authors and Affiliations
Corresponding author
Appendices
Appendix 1
Identification key of clonal growth forms of vascular plants in eastern Ladakh, Western Himalayas
-
1a stems only above-ground................................................................................................................2
-
1b stems also below-ground................................................................................................................7
-
2a neither adventitious roots nor buds present on roots (non-clonal plants)...............................................3
-
2b horizontal rooting stem...................................................................................................................4
-
2c plant fragments...............................................................................................................................5
-
3a annual and biennial herbs (“Microula tibetica” type)
-
3b trees (“Juniperus semiglobosa” type)
-
3c cushion plants (“Thylacospermum caespitosum” type)
-
4a horizontal stems short-lived (up to two growing seasons)
-
Short-lived horizontal rooting stems on or above soil surface:
-
short-lived herbaceous clonal growth organ rooting in the soil and providing connection between offspring plants or formed by creeping axis; nodes on stem bearing leaves, internodes usually long, stem serving as a storage organ and bud bank; vegetative spreading usually fast and persistence low (“Halerpestes sarmentosa” type)
-
4b horizontal stems long-lived (more than two growing seasons)
-
Long-lived horizontal rooting stems on soil surface:
-
long-lived, usually woody, clonal growth organ formed by creeping axis, rooting in the soil; nodes of youngest parts bearing leaves, internodes shorter than in short-lived horizontal stems; stem serving as a storage organ and bud bank; vegetative spreading slow and persistence high (“Thymus linearis” type)
-
5a plant fragments specialized for overwintering (turions)
-
Turions:
-
detachable over-wintering buds (usually in water plants) composed by tightly arranged leaves filled with food reserves; turions, developing axially or apically and needing vernalisation to re-growth (“Potamogeton amblyphyllus” type)
-
5b unspecialized plant fragments
-
Plant fragments of stem origin:
-
detached parts of shoot with rooting ability (“Potamogeton perfoliatus” type)
-
5c detachable offspring........................................................................................................................6
-
6a offspring in inflorescence (pseudovivipary)
-
Plantlets (pseudovivipary):
-
meristem normally develops into a flower, but forms vegetative buds (plantlets, bulbils, root or stem tubercules), which are sometimes soon detached from the parent plant; alternatively the entire inflorescence falls off and the plantlets root at the soil surface; offspring size similar to seedlings (“Bistorta vivipara” type)
-
6b offspring in axils of leaves
-
Axillary buds:
-
small vegetative diaspores produced in axils of leaves on stems above-ground formed by axillary buds subtended by small storage organ of stem, root (tubercule) of leaf (bulbil) origin; buds soon shed from mother plant, beginning to grow immediately; resembling seedlings in size (“Saxifraga cernua” type)
-
7a below-ground stems lacking adventitious roots and roots lacking adventitious buds.............................................................................................................................................8
-
7b below-ground stems possessing adventitious roots and/or roots possessing adventitious buds.............................................................................................................................................9
-
8a long below-ground stems (pleiocorm)
-
Pleiocorm having long branches:
-
plant possessing a primary root system lacking adventitious roots and buds; tap root serving as storage organ; bud bank situated on perennial stems with long (more than 10 cm) branches serving as vascular link between shoots and primary root; non-clonal plants (“Saussurea gnaphalodes” type)
-
8b short below-ground stems (pleiocorm)
-
Pleiocorm having short branches:
-
plant possessing a primary root system lacking adventitious roots and buds; tap root serving as storage organ; bud bank situated on perennial stems with short (less than 10 cm) branches serving as vascular link between shoots and primary root; non-clonal plants (“Arnebia euchroma” type)
-
9a roots with adventitious buds.........................................................................................................10
-
9b roots without adventitious buds....................................................................................................11
-
10a adventitious buds on main root
-
Main root with adventitious buds:
-
main root (include hypocotyle) forming adventitious buds spontaneously or after injury; clonal growth usually only after fragmentation of main root (“Parrya nudicaulis” type)
-
10b adventitious buds on horizontal creeping roots
-
Horizontal roots with adventitious buds:
-
branches of main root and adventitious roots forming adventitious buds spontaneously or after injury; roots serving as bud bank and vascular connection between offspring shoots; lateral spread usually extensive; persistence differing among species (“Ptilotrichum canescens” type)
-
11a below-ground stems lacking specialized storage organs............................................................12
-
11b below-ground stems possessing specialized storage organs, stems sometimes reduced.............15
-
12a stems formed below-ground (hypogeogenous rhizomes)...........................................................13
-
12b stems formed at soil surface and older parts placed below-ground (epigeogenous rhizomes)...................................................................................................................................14
-
13a hypogeogenous rhizomes with short increments (less than 10 cm)
-
Short hypogeogenous rhizomes:
-
perennial organs of stem origin formed below-ground; rhizome usually growing at a species specific depth, periodically becoming orthotropic and forming above-ground shoots; horizontal part of the rhizome bearing bracts, some roots and possessing short internodes; vegetative spreading intermediate; persistence differing considerably among species (“Leontopodium ochroleucum” type)
-
13b hypogeogenous rhizomes with long increments (more than 10 cm)
-
Long hypogeogenous rhizomes:
-
perennial organs of stem origin formed below-ground; rhizome usually growing at a species-specific depth, periodically becoming orthotropic and forming above-ground shoots; horizontal part of the rhizome bearing bracts, some roots possessing long internodes; vegetative spreading fast; persistence differing considerably among species (“Poa tibetica” type)
-
14a Short epigeogenous rhizomes:
-
perennial organ of stem origin formed above-ground; its distal part covered by soil and litter or pulled into the soil by contraction of roots; nodes bearing green leaves; internodes usually short; rhizomes bearing roots and serving as a bud bank and storage organ; vegetative spread low (up to one cm per year); persistence usually low; typical of tussock grasses (“Festuca kashmiriana” type)
-
14b Medium long epigeogenous rhizomes:
-
perennial organ of stem origin formed above-ground; its distal part covered by soil and litter or pulled into the soil by contraction of roots; nodes bearing green leaves; internodes usually short; rhizomes bearing roots and serving as a bud bank and storage organ; vegetative spread low (up to a few cm per year); persistence usually low (“Cremanthodium ellisii” type)
-
14c Long epigeogenous rhizomes:
-
perennial organ of stem origin formed above-ground; its distal part covered by soil and litter or pulled into the soil by contraction of roots; nodes bearing green leaves; internodes usually short; rhizomes bearing roots and serving as a bud bank and storage organ; vegetative spread low (up to a few cm per year); persistence usually long, resulting in a large preserved rhizome system (“Biebersteinia odora” type)
-
15a storage in leaves
-
Bulbs:
-
storage and perennation organ consisting of storage leaves and shortened stem base; stem providing bud bank and connection between offspring shoots; lateral spread low (“Lloydia serotina” type)
-
15b storage in stem
-
Stem tubers:
-
below-ground, usually short-lived storage and regenerative organ of shoot origin; lateral spread high in the case of offspring tubers formed on hypogeogenous rhizome, persistence usually low (“Potamogeton filiformis” type)
Rights and permissions
About this article
Cite this article
Klimešová, J., Doležal, J., Dvorský, M. et al. Clonal Growth Forms in Eastern Ladakh, Western Himalayas: Classification and Habitat Preferences. Folia Geobot 46, 191–217 (2011). https://doi.org/10.1007/s12224-010-9076-3
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s12224-010-9076-3