
Abstract
Trace amounts of the carcinogenic ethyl carbamate can appear in wine as a result of a reaction between ethanol and citrulline, which is produced from arginine degradation by some bacteria used in winemaking. In this study, arginine deiminase (ADI) pathway genes were evaluated in 44 Oenococcus oeni strains from wines originating from several locations in order to establish the relationship between the ability of a strain to degrade arginine and the presence of related genes. To detect the presence of arc genes of the ADI pathway in O. oeni, pairs of primers were designed to amplify arcA, arcB, arcC and arcD1 sequences. All strains contained these four genes. The same primers were used to confirm the organization of these genes in an arcABCD1 operon. Nevertheless, considerable variability in the ability to degrade arginine among these O. oeni strains was observed. Therefore, despite the presence of the arc genes in all strains, the expression patterns of individual genes must be strain dependent and influenced by the different wine conditions. Additionally, the presence of arc genes was also determined in the 57 sequenced strains of O. oeni available in GenBank, and the complete operon was found in 83 % of strains derived from wine. The other strains were found to lack the arcB, arcC and arcD genes, but all contained sequences homologous to arcA, and some of them had also ADI activity.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
Oenococcus oeni is a species of lactic acid bacteria (LAB) that is mainly involved in malolactic fermentation (MLF), which usually occurs after alcoholic fermentation (AF) during the winemaking process (Wibowo et al. 1985). MLF involves the decarboxylation of l-malic acid into l-lactic acid and CO2, which decreases the acidity of the wine and produces a softer taste. O. oeni also produces other interesting compounds, such as diacetyl, which can improve the aromatic quality of wine (Henick-Kling 1993; Lonvaud-Funel 1999), and the microbiological stability of wine that has undergone MLF is also improved (Davis et al. 1985; Wibowo et al. 1985).
However, two groups of undesirable substances that are potentially harmful to human health may be released by LAB during and after winemaking: biogenic amines and precursors of ethyl carbamate (or urethane) (Lonvaud-Funel 1999). Wine, like most fermented foods and beverages, can contain trace amounts of ethyl carbamate (EC) (Ough 1976), which is an animal carcinogen (Schlatter and Lutz 1990). The recommended maximum level of EC in table wines is 15 μg/L, and 30 μg/L is the legal limit in some countries, such as Canada. EC is formed by a spontaneous chemical reaction between ethanol and carbamyl group containing compounds such as urea or citrulline (Ough et al. 1988). The urea produced by yeast arginine metabolism is the main precursor in EC production, but there is also evidence that EC levels can increase after MLF (Uthurry et al. 2004; Uthurry et al. 2006). EC-forming reactions are favoured by high temperature and an acidic pH (Stevens and Ough 1993).
Several studies have examined the role of wine LAB in EC formation (Liu et al. 1994; Liu and Pilone 1998; Arena et al. 1999). Many wine LAB strains are known to degrade l-arginine (Mira de Orduña et al. 2000), which is one of the main amino acids found in grapes and wine (Lehtonen 1996). It has been demonstrated that arginine catabolism by these LAB involves the arginine deiminase (ADI) pathway (Liu et al. 1995; Liu et al. 1996). This pathway is the most widespread anaerobic route for arginine degradation (Cunin et al. 1986), and its presence has been reported in bacteria, archaea, and some anaerobic eukaryotes (Zúñiga et al. 2002).
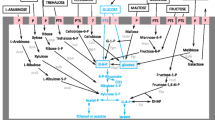
This pathway includes three enzymes—ADI (EC 3.5.3.6), ornithine transcarbamylase (EC 2.1.3.3, OTC), and carbamate kinase (EC 2.7.2.2, CK)―that catalyse the following reactions:
Theoretically, this pathway plays a positive role in the growth and viability of LAB because ATP is formed, and the production of ammonia decreases the acidity of the wine (Tonon and Lonvaud-Funel 2000), similar to LAB found in other acidic foods, as the apple surface (Savino et al. 2012). However, this reaction may differ among the different wine LAB strains, and it has been shown that arginine and citrulline do not stimulate the growth of some O. oeni strains (Terrade and Mira de Orduña 2009). However, if LAB degrades arginine by the ADI pathway, citrulline can be produced (Mira de Orduña et al. 2000), which can react spontaneously with ethanol to form EC.
It has been shown that O. oeni strains can catabolize arginine. Mira de Orduña et al. (2000) tested 23 O. oeni strains and found that all strains degraded arginine and excreted citrulline. Pilone et al. (1991) observed that 18 of 24 strains tested produced ammonia from arginine. Granchi et al. (1998) found 30 arginine-consuming strains among 32 that had been isolated from the Chianti region. Liu et al. (1995) found considerable variability among the arginine degradation pathways of 22 O. oeni strains (commercially available strains and those isolated in New Zealand), including five non-degradative strains. Finally, Tonon et al. (2001a) also found 13 different arginine-consuming strains among 17 in the Bordeaux Oenology collection.
Arginine degradation is influenced by different wine conditions. Malic acid inhibits this process because MLF, which consumes malic acid, seems to take priority over arginine consumption for LAB (Mira de Orduña et al. 2001; Terrade and Mira de Orduña 2006; Araque et al. 2011). The presence of ethanol has been shown to increase arginine utilization, and the expression of arc genes in strains of Lactobacillus brevis and Pediococcus pentosaceus may occur as a result of a stress response mechanism (Araque et al. 2013).
As with other bacteria (Maghnouj et al. 1998; Griswold et al. 2004), genes encoding the three enzymes in this pathway in LAB are clustered in an operon structure: arcA (ADI), arcB (OTC) and arcC (CK). These genes have been characterized in LAB species such as Lactobacillus sakei (Zúñiga et al. 1998), Lactobacillus hilgardii (Arena et al. 2002), Lactobacillus plantarum (Spano et al. 2004; Spano et al. 2006), and Lactobacillus fermentum (Vrancken et al. 2009). They have also been characterized in O. oeni (Tonon et al. 2001b).
Other protein-coding genes, such as transcriptional regulators (arcR, ArgR/AhrC) and arginine/ornithine transporters (arcD1, arcD2), have been described in some LAB arc operons (Kok et al. 2005; Vrancken et al. 2009), including that of O. oeni (Divol et al. 2003). Genes for other proteins that are not related to the ADI pathway have also been described in arc operons. For example, arcT for a putative transaminase and argS for arginyl-tRNA synthetase have both been described in Lactococcus lactis (Bolotin et al. 2001; Kok et al. 2005). An argS2 for a putative arginyl-tRNA synthetase has also been described at the 3′ end of the arc operon in O. oeni (Nehmé et al. 2006).
Although the arc genes have been characterized, few studies have analysed whether they are present in O. oeni strains (Tonon et al. 2001b; Divol et al. 2003), and no representative screening of O. oeni strains from different sources has been reported. The arc genes have been detected in several strains of different wine LAB species, including eight O. oeni, by means of degenerate primers (Araque et al. 2009). In this work, the arc genes were detected in arginine-degrading strains of all assayed heterofermentative lactobacilli, O. oeni and P. pentosaceus and in some Leuconostoc mesenteroides and Lb. plantarum strains.
Nevertheless, in the study of Araque et al. (2009), arc gene presence was evaluated in a few O. oeni strains. For this reason, and given the importance of this species in vinification, in the current study arc genes in several O. oeni strains have been detected by PCR amplification using species-specific primers. A wide range of strains from different sources has been analysed in order to establish a relationship between the strain’s ability to degrade arginine and the presence of arc genes.
Additionally, in order to corroborate the results and to study the presence of arc genes more globally in O. oeni, we have searched for these genes in silico in the sequenced strains available in GenBank, comparing them using BLAST with the published arc genes sequences from the type strain of O. oeni (Tonon et al. 2001b; Divol et al. 2003).
Materials and methods
Microorganisms and growth conditions
A total of 44 strains of O. oeni of different origins (Table 1) were used, including the type culture, 10 strains from CECT (the official Spanish culture collection), 15 strains from around the world (USA, South Africa, and mostly Italy), eight strains from different Catalan cellars, five commercial strains and five strains isolated from the university’s experimental cellar in Tarragona, Catalonia, Spain. All strains were grown anaerobically at 27 °C in MRS (De Man et al. 1960) at pH 5.0, supplemented with 4 g/L d,l-malic acid and 5 g/L d(−)-fructose.
Arginine degradation
Cells grown in MRS with malic acid and fructose (see above) were inoculated (1 % v/v) in 5 mL of a modified MRS (Tonon et al. 2001b) at pH 4.5, supplemented with 5 g/L of arginine and 5 g/L of glucose, and incubated anaerobically at 27 °C. Samples were taken at the end of the logarithmic phase, and all assays were performed in duplicate. Arginine and citrulline supernatant concentrations were quantified by HPLC, according to the method reported by Gómez-Alonso et al. (2007).
Determination of arginine deiminase activity
Cells grown as above were harvested by centrifugation at 10,000×g for 15 min, and the pellet was washed twice with 0.2 mol/L sodium phosphate buffer, pH 6.5 and then resuspended at 2.5 % (w/v) in the same buffer. Then, cellular extracts were obtained using a cell disrupter (BASIC Z; Constant Systems Ltd., Daventry, UK) at a pressure of 2.5 kbar. Unbroken cells and cellular debris were removed by centrifugation at 13,000×g for 6 min, and the supernatant extract was used. All operations were carried out at 4 °C. Arginine deiminase (ADI) activity of extracts was determined following Spano et al. (2007) by analysing citrulline produced from arginine.
Genomic DNA extraction
DNA was prepared by extraction with chloroform and previous incubation with lysozyme (2.5 mg/mL) with TE buffer, according to the method reported by Persing et al. (1993) with some modifications (see Reguant and Bordons 2003).
Detection of arc genes in O. oeni
Starting from the published sequences of the arcABC cluster in O. oeni ATCC 23279T (Tonon et al. 2001b) [GenBank accession number AF124851, National Center for Biotechnology Information (NCBI, http://www.ncbi.nlm.nih.gov)] and arcD genes (Divol et al. 2003) [GenBank accession number AF541253], a pair of primers was designed for each of the following genes: arcA, arcB, arcC and arcD1. General recommendations on primer design were followed (Ausubel et al. 1989; Iowa State University 2005). Fragments were verified using the Amplify 1.2 software (Engels 1993) to avoid secondary structure formation. The Basic Local Alignment Search Tool (BLAST) program (Altschul et al. 1990) at NCBI was also used to check that the primers did not hybridize with other fragments from the same microorganism.
Finally, PCR conditions were optimized for four pairs of primers synthesized by Ecogen (Barcelona). This approach yielded amplified fragments of different sizes that were easily discriminated by electrophoresis (Table 2).
Amplification was performed in a GeneAmp PCR System thermocycler (Perkin–Elmer, Applied Biosystems, Foster City, CA, USA). The PCR volume was 20 μL and contained 20 ng of genomic DNA, 2.5 mmol/L MgCl2, 0.2 mmol/L of each dNTP, 0.5 U Taq DNA polymerase and 0.2 μmol/L of each primer in 1× PCR buffer. The amplification programme was 94 °C for 1 min, 60 °C for 1 min and 72 °C for 1 min, which was repeated for 30 cycles. Also included were a preincubation step of 94 °C for 2 min and a final extension of 72 °C for 10 min.
The amplification products were resolved by electrophoresis in 1 % (w/v) agarose TBE (Tris–HCl 89 mmol/L, boric acid 89 mmol/L, Na2-EDTA 2 mmol/L, pH 8.0) gels stained with ethidium bromide and photographed. A 100-bp ladder (Invitrogen SA, Barcelona) was included as a DNA molecular size marker.
Genomic organization of the O. oeni arc operon
To confirm the organization of the arc operon containing the four genes arcA, arcB, arcC and arcD1, corresponding pairs of primers were combined to obtain six PCR-amplified fragments (Fig. 1). DNA was obtained from O. oeni 4772. The procedure was the same as above but the amplification products were resolved in 0.8 % (w/v) agarose TBE gels, and a 250-bp ladder (Invitrogen S.A.) was included as a DNA molecular size marker.

The arc operon of Oenococcus oeni. Its location in the chromosome (left upper circle) is approximate. Other elements of this gene cluster are not shown. The arrows (→forward, ← reverse) indicate the positions of the primers used (see Table 2). Below, the PCR amplified fragments of each gene and those that confirm the operon structure (size in bp) are represented. In the left-hand column, the forward (f) and reverse (r) pairs of primers used for each amplification are indicated: A, B, C and D correspond to arcA, arcB, arcC and arcD1, respectively
The presence of arc genes in the sequenced strains of O. oeni
The abovementioned published sequences of arc genes from O. oeni ATCC 23279T (=DSM 20252) (Tonon et al. 2001b; Divol et al. 2003) were compared using the BLASTN 2.2.30+ method to somewhat similar sequences (Altschul et al. 1997) deposited to the NCBI website as of January 2015 from the 57 sequenced O. oeni genomes available at the NCBI (Table 3). In the search set, “whole-genome shotgun contigs” and “refseq genomic” databases, both for O. oeni taxid 1247, were used.
Results
Arginine degradation by O. oeni strains
As Table 1 shows, considerable variability was observed in the ability of the different O. oeni strains to degrade arginine in a complex medium. More than half of the strains (25 out of 44, i.e., 57 %) degraded more than 50 % of 5 g/L (29 mmol/L) arginine in MRS medium at pH 4.5 at the end of logarithmic phase (approx. 7 days). These strains were of very different origins and included the type culture and some commercial strains. The other 19 strains only partly degraded arginine or exhibited hardly any signs of degradation. Once again, these strains were from different sources and two of them were commercially available. Therefore, there is considerable variability among strains with regard to arginine degradation, and it seems that there is no relationship between the origin of a strain and its ability to degrade arginine.
Table 1 also shows the growth of the strains during the arginine degradation assay, as measured by absorbance at 600 nm. There was no relationship between increased arginine consumption and greater growth because some strains, such as strain 4760, degraded arginine completely and grew slightly, while others consumed very little arginine and grew considerably (strain OSU).
Supernatant citrulline concentrations at the end of assays were slightly higher in some arginine-degrading strains, reaching values over 200 mg/L (see Table S1 in the supplementary data), while they were clearly lower, with values lower than 20 mg/L in most strains with lesser arginine degradation. As can be seen, the ratios of citrulline-produced versus arginine consumed were between 0 and 9 % for the different strains.
Arginine deiminase activity
Enzymatic activity of ADI, corresponding to arcA gene, was preliminary studied in some strains of O. oeni. Variable values of activity were found in them from 5 U/mg protein (strains Viniflora oenos, 4727, MB42b) to 20 U/mg protein (217T strain). Additionally, PSU-1 strain, which has only arcA according its sequenced genome (see below), was analysed and it had an ADI activity of 9.5 U/mg protein.
Detection of arc genes in O. oeni
Four pairs of primers were designed for each arc gene cluster from the known sequences in O. oeni, and PCR conditions were optimized so that different fragments were obtained. As Table 2 shows, amplified fragments of different sizes were easily discriminated by electrophoresis.
The four pairs of primers were assayed with DNA isolated from 44 strains of different origins (Table 1). All of these strains contained the four genes of the ADI pathway. As an example, the electrophoretic bands obtained for two strains of O. oeni are shown in Fig. 2. These bands presented the expected sizes of the four amplified fragments and were in agreement with those described in Table 2.

The arcABCD genes in two strains of Oenococcus oeni (217T and Mf15). Genes are arranged (lanes 1 to 4, strain 217T; lanes 5 to 8, strain Mf2): arcA, arcB, arcC and arcD1 and sizes are as expected: 477, 892, 802 and 670 bp, respectively. An Invitrogen DNA 100-bp ladder was used
Genomic organization of the arc operon in O. oeni
When the four pairs of primers used to detect the arc genes of O. oeni (Table 2) were combined to obtain six PCR-amplified fragments including two or more genes of the same operon (Fig. 1), the sizes obtained by electrophoresis were as expected (Fig. 3). The sizes of these fragments were the same as the theoretical values obtained with the Amplify 1.2 software (Engels 1993). As shown, there were no secondary products except for some amplification between the arcB forward and arcD reverse primers, where a secondary band of approx. 750 bp appeared and was most likely due to unspecific hybridization.

PCR-amplified fragments of the arc operon of Oenococcus oeni, which confirm the operon structure by combining pairs of primers of the four genes. For each pair, the first letter corresponds to the forward primer and the second letter corresponds to the reverse one, where A, B, C and D are the genes arcA, arcB, arcC and arcD1, respectively. Lane M is an Invitrogen DNA 250-bp ladder
Presence of arc genes in the sequenced O. oeni strains
As seen in Table 3, there were published genome sequences available for 57 strains of O. oeni that had been deposited in GenBank from 2006 until 2014. Only one strain had a complete sequence reported (PSU-1), and as this was the first sequenced it was used as the representative genome. The sequence of another strain (ATCC BAA-1163) was at the scaffold level, and all other strains were at the contig draft level. Sequenced strains in Table 3 are grouped by the presence of the different arc genes and by the submitter organization, which promoted the corresponding Genome Annotation Project. As expected, the sequence identity found for type strain DSM 20252 (=AWRIB129 = ATCC 23279) in all genes was 100 % because the arc gene sequences reported were derived from this strain.
The arcA gene (Table 3) was present in all of the sequenced strains, with a query coverage and identity of 99–100 %, but in 14 strains (25 % of all evaluated), including the representative genome of PSU-1 strain, the identity observed was just 68 %. The other arc genes (arcB, arcC, arcD1 and arcD2) were found to be present in 44 strains (77 % of the total strains evaluated). The query coverage and identity was 99–100 % in all of these strains for the four genes examined, but a slightly lower coverage of 88–95 % for the arcD2 gene was observed in three strains.
The query coverage for these four genes in the other 13 strains (23 % of total) was lower than 50 %, and thus, these genes are likely absent in these strains. These are the same strains for which the arcA gene identity was 68 %, with the exception of strain IOEB_0501, which contains all the other genes. As shown, the four sequenced strains isolated from cider and included in this group lack the genes arcB, arcC, arcD1 and arcD2. All other sequenced O. oeni strains were isolated from wine.
Discussion
This work shows that there is considerable variability in the ability of different O. oeni strains to degrade arginine. The arginine consumption of different strains was studied in a rich media (MRS medium), as other authors have done (Liu et al. 1995; Tonon et al. 2001b). Relatively high levels of arginine (5 g/L) were used because the cell’s ability to consume is more clearly revealed at this concentration, as other authors have reported (Pilone et al. 1991). Similarly, a pH of 4.5 was used in spite of being too high for wine conditions because arginine metabolism occurs more readily at this pH and is inhibited at pH values lower than 3.5, as shown by Terrade and Mira de Orduña (2006). Previous studies have also found variability in the arginine consumption of different O. oeni strains, but in this respect, the main contribution of this work is that several strains from very different sources have been used.
Citrulline levels have been considered as good indicators of the final EC concentrations found in wine. The possible production of citrulline from arginine during MLF therefore strongly depends on the strain (Romero et al. 2009). This reinforces the idea that the ability of each strain used in MLF to degrade arginine needs to be assessed, and this information should be used to select the starter cultures. Here, we have seen some relationship between the ability of arginine degradation and the production of citrulline, which varies depending on the strain. The ratios of citrulline produced versus arginine consumed were lower than 10 % for all strains, and there was no clear relationship of these ratios to the level of arginine consumption or the origin or characteristics of strains. Similar ratios have been found in previous works for strains of O. oeni, such as 4 % (Terrade and Mira de Orduña 2009) or 6 % (Mira de Orduña et al. 2001), but higher ratios have been found for other LAB species: 10 % for Lactobacillus buchneri (Liu and Pilone 1998) or near 20 % for Lb. hilgardii (Araque et al. 2011).
This is the first time that a large number of O. oeni strains have been screened for the presence of the arc genes. It has been shown that all 44 strains tested contained the four genes of the arc operon. Because these strains were from very different sources, and included some commercial MLF starters, we can suggest that all O. oeni strains would have these genes. As a positive control, there were included strains such as the type strain in which the presence of these genes has already been demonstrated (Tonon et al. 2001b).
The presence of the arc operon in all of the tested strains is consistent with the fact that all strains consume arginine, although some to a very small extent. Other authors have reported that some O. oeni strains with no arc genes are unable to degrade arginine (Tonon et al. 2001b; Divol et al. 2003). Nevertheless, it has been shown that some of these non-arginine-degrading strains can sometimes have a faint amplification of the arcA gene (Tonon et al. 2001b). Therefore, the use of arginine can vary, including some cases of absent or very minor arginine degradation regardless the presence of the arc genes. This variability supports the idea that environmental factors are essential for the expression of these genes, depending on the strain.
Using all of the combinations of forward and reverse primers for the four genes arcA, arcB, arcC and arcD1 in O. oeni strains, amplified fragments of the expected sizes (Fig. 3) from the theoretical values were obtained (Fig. 1). As revealed by Divol et al. (2003), there is another arcD2 gene, but it was not included in the current study because arcD1 and arcD2 have been shown to be very similar, both codifying for the arg/orn antiport, and because arcD1 is the one closely located to arcABC. These data confirm that the four genes are organized in an arcABCD1 operon in the tested strains and correlate well with previously reported information (Divol et al. 2003). Other LAB that have been studied, such as Lb. sakei (Zúñiga et al. 1998), Lb. plantarum (Spano et al. 2004) and Lc. lactis (Bolotin et al. 2001; Kok et al. 2005), have slightly different operon organizations.
In other wine LAB, such as Lb. plantarum, a great variability in ADI activity was observed among different strains (Spano et al. 2007). Those strains with higher ADI activity and lower OTC activity accumulate greater amounts of citrulline and thus produce more EC. In the current work, we observed a variable ADI activity among tested strains of O. oeni. Interestingly, the representative sequenced strain PSU-1, which has only the arcA gene (see Table 3), presented similar ADI activity to other strains having all the arc operon, demonstrating that this separate arcA gene codifies also for an active enzyme.
To determine whether arc genes are present in other strains of O. oeni besides those tested in this study, we searched for their presence in the sequenced strains available in GenBank. There was found that all 57 strains (Table 3) have sequences that are homologous to arcA; in spite of this information, 14 strains (25 % of them) have an identity of only 68 % for this gene. With the exception of strain IOEB_0501, all of these strains (23 % of total) that have a partial identity to arcA lack the other four genes of the arc operon. The only four sequenced strains of O. oeni isolated from cider are included in this group and were shown to be phylogenetically clustered and separated from other strains upon SNP analysis (El Khoury 2014).
As seen in the GenBank data, the sequenced strains containing all arc genes have localized them to the same contig with one gene after another structured as an operon, which is the same as that seen in the strains of this work. Nevertheless, the strain IOEB_0501 presents the gene arcA (with an identity of 68 %) in a different genome location of genome separated from arcB, arcC, arcD1 and arcD2, as reported previously (Mills et al. 2005; Bon et al. 2009). On the other hand, these four genes are placed together, and upstream of that cluster appears a sequence with certain homology to arcA (38 % query coverage) which could be a nonfunctional evolutionary remnant of this gene.
Therefore, it can be concluded that all strains of O. oeni, those sequenced and deposited in GenBank and those studied in this work, contain the arcA gene. Regarding the other genes of arc operon, despite the fact that all strains tested here contained all of the operon genes, it has been observed that some of the sequenced strains are lacking these genes. In this way, we can suggest that the presence of the arcB, arcC and arcD genes is not universal in O. oeni because they can be found in approximately 77 % of sequenced strains. Excluding the cider-derived strains, the absence of those genes is limited to nine sequenced strains and the presence of all arc genes in sequenced strains of O. oeni isolated from wine increases to 83 %.
The results of the current study leads to a better understanding of how the different O. oeni strains can degrade arginine and in this way influence the formation of ethyl carbamate precursors. Having observed that the arcA gene is present in all studied strains of this species, a more in-depth study of the expression of this gene in different conditions is now required. Screening for safer strains should not only analyse arginine degradation but also seek strains having the complete operon or those with maximum specific activity for the final enzyme in the pathway, CK, so that citrulline does not accumulate.
References
Altschul SF, Gish W, Miller W, Myers EW, Lipman DJ (1990) Basic local alignment search tool. J Mol Biol 215:403–410
Altschul SF, Madden TL, Schäffer AA, Zhang J, Zhang Z, Miller W, Lipman DJ (1997) Gapped BLAST and PSI-BLAST: a new generation of protein database search programs. Nucl Acids Res 25:3389–3402
Araque I, Gil J, Carreté R, Bordons A, Reguant C (2009) Detection of arc genes related with the ethyl carbamate precursors in wine lactic acid bacteria. J Agric Food Chem 57:1841–1847
Araque I, Reguant C, Rozès N, Bordons A (2011) Influence of wine-like conditions on arginine utilization by lactic acid bacteria. Int Microbiol 14:225–233
Araque I, Bordons A, Reguant C (2013) Effect of ethanol and low pH on citrulline and ornithine excretion and arc gene expression by strains of Lactobacillus brevis and Pediococcus pentosaceus. Food Microbiol 33:107–113
Arena ME, Saguir FM, Manca de Nadra MC (1999) Arginine, citrulline and ornithine metabolism by lactic acid bacteria from wine. Int J Food Microbiol 52:155–161
Arena ME, Manca de Nadra MC, Muñoz R (2002) The arginine deiminase pathway in the wine lactic acid bacterium Lactobacillus hilgardii X1B: structural and functional study of the arcABC genes. Gene 301:61–66
Ausubel FM, Brent R, Kingston RE, Moore DD, Seidmann JG, Smith JA, Struhl K (1989) Current protocols in molecular biology. Wiley, New York, vols 1–3
Bolotin A, Wincker P, Mauger S, Jaillon O, Malarme K, Weissenbach J, Ehrlich SD, Sorokin A (2001) The complete genome sequence of the lactic acid bacterium Lactococcus lactis ssp. lactis IL1403. Genome Res 11:731–753
Bon E, Delaherche A, Bilhère E, De Daruvar A, Lonvaud-Funel A, Le Marrec C (2009) Oenococcus oeni genome plasticity is associated with fitness. Appl Environ Microbiol 75:2079–2090
Cunin R, Glansdorff N, Piérard A, Stalon V (1986) Biosynthesis and metabolism of arginine in bacteria. Microbiol Rev 50:314–352
Davis CR, Wibowo D, Eschenbruch R, Lee TH, Fleet GH (1985) Practical implications of malolactic fermentation: a review. Am J Enol Vitic 36:290–301
De Man JC, Rogosa M, Sharpe ME (1960) A medium for the cultivation of lactobacilli. J Appl Bacteriol 23:130–135
Divol B, Tonon T, Morichon S, Gindreau E, Lonvaud-Funel A (2003) Molecular characterization of Oenococcus oeni genes encoding proteins involved in arginine transport. J Appl Microbiol 94:738–746
El Khoury M (2014) Étude de la diversité des souches d’Oenococcus oeni responsables de la fermentation malolactique des vins dans différentes régions vitivinicoles. PhD thesis. Univ Bordeaux, Talence, France.
Engels WR (1993) Contributing software to the Internet: the Amplify program. Trends Biochem Sci 18:448–450
Gómez-Alonso S, Hermosín-Gutiérrez I, García-Romero E (2007) Simultaneous HPLC analysis of biogenic amines, amino acids and ammonium ion as aminoenone derivatives in wine and beer samples. J Agric Food Chem 55:608–613
Granchi L, Paperi R, Rosellini D, Vincenzini M (1998) Strain variation of arginine catabolism among malolactic Oenococcus oeni strains of wine origin. Ital J Food Sci 10:351–357
Griswold A, Chen YYM, Snyder JA, Burne RA (2004) Characterization of the arginine deiminase operon of Streptococcus rattus FA-1. Appl Environ Microbiol 70:1321–1327
Henick-Kling T (1993) Malolactic fermentation. In: Fleet GH (ed) Wine microbiology and biotechnology. Harwood Academic, Chur, pp 289–326
Iowa State University (2005) The DNA Facility of the Iowa State University Office of Biotechnology. http://www.dna.iastate.edu.
Kok J, Buist G, Zomer AL, van Hijum AFT, Kuipers OP (2005) Comparative and functional genomics of lactococci. FEMS Microbiol Rev 29:411–433
Lehtonen P (1996) Determination of amines and amino acids in wine—a review. Am J Enol Vitic 47:127–133
Liu SQ, Pilone GJ (1998) A review: arginine metabolism in wine lactic acid bacteria and its practical significance. J Appl Microbiol 84:315–327
Liu SQ, Pritchard GG, Hardman MJ, Pilone GJ (1994) Citrulline production and ethyl carbamate (urethane) precursor formation from arginine degradation by wine lactic acid bacteria Leuconostoc oenos and Lactobacillus buchneri. Am J Enol Vitic 45:235–242
Liu SQ, Pritchard GG, Hardman MJ, Pilone GJ (1995) Occurrence of arginine deiminase pathway enzymes in arginine catabolism by wine lactic acid bacteria. Appl Environ Microbiol 61:310–316
Liu SQ, Pritchard GG, Hardman MJ, Pilone GJ (1996) Arginine catabolism in wine lactic acid bacteria: is it via the arginine deiminase pathway or the arginase-urease pathway? J Appl Bacteriol 81:486–492
Lonvaud-Funel A (1999) Lactic acid bacteria in the quality improvement and depreciation of wine. A Van Leeuw 76:317–331
Maghnouj A, Cabral TFS, Stalon V, Wauven CV (1998) The arcABDC gene cluster, encoding the arginine deiminase pathway of Bacillus licheniformis, and its activation by the arginine repressor ArgR. J Bacteriol 180:6468–6475
Mills DA, Rawsthorne H, Parker C, Tamir S, Makarova K (2005) Genomic analysis of Oenococcus oeni PSU-1 and its relevance to winemaking. FEMS Microbiol Rev 29:465–475
Mira de Orduña R, Liu SQ, Patchett ML, Pilone GJ (2000) Ethyl carbamate precursor citrulline formation from arginine degradation by malolactic wine lactic acid bacteria. FEMS Microbiol Lett 183:31–35
Mira de Orduña R, Patchett ML, Liu SQ, Pilone GJ (2001) Growth and arginine metabolism of the wine lactic acid bacteria Lactobacillus buchneri and Oenococcus oeni at different pH values and arginine concentrations. Appl Environ Microbiol 67:1657–1662
Nehmé B, Ganga MA, Lonvaud-Funel A (2006) The arginine deiminase locus of Oenococcus oeni includes a putative arginyl-tRNA synthetase ArgS2 at its 3′-end. Appl Microbiol Biotechnol 70:590–597
Ough CS (1976) Ethyl carbamate in fermented beverages and foods. I. Naturally occuring ethyl carbamate. J Agric Food Chem 24:323–328
Ough CS, Crowell EA, Gutlove BR (1988) Carbamyl compound reactions with ethanol. Am J Enol Vitic 39:239–242
Persing DH, Smith TF, White TJ, Tenover FC (1993) Diagnostic molecular biology, principles and application. ASM, Washington DC
Pilone GJ, Clayton MG, van Duivenboden RJ (1991) Characterization of wine lactic acid bacteria: single broth culture for tests of heterofermentation, mannitol from fructose and ammonia form arginine. Am J Enol Vitic 42:153–157
Reguant C, Bordons A (2003) Typification of Oenococcus oeni strains by multiplex RAPD-PCR and study of population dynamics during malolactic fermentation. J Appl Microbiol 95:344–353
Romero SV, Reguant C, Bordons A, Masqué MC (2009) Potential formation of ethyl carbamate in simulated wine inoculated with Oenococcus oeni and Lactobacillus plantarum. Int J Food Sci Technol 44:1206–1213
Savino MJ, Sánchez LA, Saguir FM, Manca de Nadra MC (2012) Lactic acid bacteria isolated from apples are able to catabolise arginine. World J Microbiol Biotechnol 28:1003–1012
Schlatter J, Lutz WK (1990) The carcinogenic potential of ethyl carbamate (urethane): risk assessment at human dietary exposure levels. Food Chem Toxicol 28:205–211
Spano G, Chieppa G, Beneduce L, Massa S (2004) Expression analysis of putative arcA, arcB and arcC genes partially cloned from Lactobacillus plantarum isolated from wine. J Appl Microbiol 96:185–193
Spano G, Beneduce L, de Palma L, Quinto M, Vernile A, Massa S (2006) Characterization of wine Lactobacillus plantarum by PCR-DGGE and RAPD-PCR analysis and identification of Lactobacillus plantarum strains able to degrade arginine. World J Microbiol Biotechnol 22:769–773
Spano G, Massa S, Arena ME, Manca de Nadra MC (2007) Arginine metabolism in wine Lactobacillus plantarum: in vitro activities of the enzymes arginine deiminase (ADI) and ornithine transcarbamilase (OTCase). Ann Microbiol 57:67–70
Stevens DF, Ough CS (1993) Ethyl carbamate formation: reaction of urea and citrulline with ethanol in wine under low to normal temperature conditions. Am J Enol Vitic 44:309–312
Terrade N, Mira de Orduña R (2006) Impact of winemaking practices on arginine and citrulline metabolism during and after malolactic fermentation. J Appl Microbiol 101:406–411
Terrade N, Mira de Orduña R (2009) Arginine and citrulline do not stimulate growth of two Oenococcus oeni strains in wine. FEMS Microbiol Lett 290:98–104
Tonon T, Lonvaud-Funel A (2000) Metabolism of arginine and its positive effect on growth and revival of Oenococcus oeni. J Appl Microbiol 89:526–531
Tonon T, Bourdineaud JP, Lonvaud-Funel A (2001a) Catabolisme de l’arginine par Oenococcus oeni: aspects energetiques et genetiques. Lait 81:139–159
Tonon T, Bourdineaud JP, Lonvaud-Funel A (2001b) The arcABC cluster encoding the arginine deiminase pathway of Oenococcus oeni, and arginine induction of a CRP-like gene. Res Microbiol 152:653–661
Uthurry CA, Varela F, Colomo B, Suárez Lepe JA, Lombardero J, García del Hierro JR (2004) Ethyl carbamate concentrations of typical Spanish red wines. Food Chem 88:329–336
Uthurry CA, Suárez Lepe JA, Lombardero J, García del Hierro JR (2006) Ethyl carbamate production by selected yeasts and lactic acid bacteria in red wine. Food Chem 94:262–270
Vrancken G, Rimaux T, Weckx S, De Vuyst L, Leroy F (2009) Environmental pH determines citrulline and ornithine release through the arginine deiminase pathway in Lactobacillus fermentum IMDO 130101. Int J Food Microbiol 135:216–222
Wibowo D, Eschenbruch R, Davis CR, Fleet GH, Lee TH (1985) Occurrence and growth of lactic acid bacteria in wine: a review. Am J Enol Vitic 36:302–313
Zúñiga M, Champomier-Verges M, Zagorec M, Pérez-Martínez G (1998) Structural and functional analysis of the gene cluster encoding the enzymes of the arginine deiminase pathway of Lactobacillus sakei. J Bacteriol 180:4154–4159
Zúñiga M, Pérez G, González-Candelas F (2002) Evolution of arginine deiminase (ADI) pathway genes. Mol Phylogen Evol 25:429–444
Acknowledgments
This work was supported by grant AGL-2000-0827-C02-02 from the Spanish Ministry of Science and Technology.
Compliance with Ethical Standards
This work complies with ethical standards. The authors declare that they have no conflict of interest. This article does not contain any studies with human participants or animals performed by any of the authors.
Author information
Authors and Affiliations
Corresponding author
Electronic supplementary material
Below is the link to the electronic supplementary material.
ESM 1
Table S1 Production of citrulline from arginine (5 g/L) in MRS medium at pH 4.5 by strains of Oenococcus oeni. Values are the averages of duplicate cultures. (DOCX 30 kb)
Rights and permissions
About this article

Cite this article
Araque, I., Gil, J., Carreté, R. et al. Arginine deiminase pathway genes and arginine degradation variability in Oenococcus oeni strains. Folia Microbiol 61, 109–118 (2016). https://doi.org/10.1007/s12223-015-0416-9
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s12223-015-0416-9