
Abstract
Interleukin-6 receptor-alpha (IL6R) interacts with IL6 and forms a ligand–receptor complex, which can stimulate various cellular responses, such as cell proliferation, cell differentiation, and activation of inflammatory processes. Both genetic mutation and epigenetic modification regulate gene transcription. We identified a novel splice variant of bovine IL6R, designated as IL6R-TV, which is characterized by the skipping of exon 2 of the NCBI-referenced IL6R gene (IL6R-reference). The expression levels of IL6R-TV and IL6R-reference transcripts were lower in normal mammary gland tissues. These transcripts play a potential role during inflammatory infection. We also detected two putative functional SNPs (g.19711 T > C and g.19731 G > C) located within the upstream 100 bp of exon 2. These SNPs formed two haplotypes (T-G and C-C). Two mutant pSPL3 exon-trapping plasmids (pSPL3-T-G and pSPL3-C-C) were transferred into the bovine mammary epithelial cells (MAC-T) and human embryonic kidney 293 T cells (HEK293T) to investigate the relationship between the two SNPs and the aberrant splicing of IL6R. DNA methylation levels of the alternatively spliced exon in normal and mastitis-infected mammary gland tissues were quantified through nested bisulfate sequencing PCR (BSP) and cloning sequencing. We found that DNA methylation regulated IL6R transcription. The DNA methylation level was high in mastitis-infected mammary gland tissues and stimulated IL6R expression, thereby promoting the inclusion of the alternatively spliced exon. The upregulated expression of the two transcripts was due to DNA methylation modification rather than genetic mutations.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
Clinical mastitis is a visible inflammatory disease that infects the mammary gland of dairy cows and the most frequent disease of dairy cattle which can be potentially fatal. Bovine mastitis is an economically important pathology associated with reduced milk production and changes in milk composition and quality (Gomes and Henriques 2016), and is considered the most costly infectious disease in dairy cows (Rainard et al. 2016). Many programs have been developed to improve bovine resistance and reduce the occurrence of mastitis. However, conventional strategies based on phenotypic selection are inefficient because of their low heritability (Hinchs et al. 2011). By contrast, genomic selection programs are an effective tool against diseases and important for cattle breeding (Wiggans et al. 2016; García-Ruiz et al. 2016).
Inflammation is a biological process commonly caused by bacterial infection and initiated upon injury to remove harmful stimuli and induce healing. However, prolonged inflammation may be detrimental to the host and promote a permanent disease state, resulting in gene expression dysregulation and tissue damage (Huang et al. 2014). Bovine mastitis leads to the overwhelming release of pro-inflammatory cytokines, which play pivotal roles in regulation of inflammatory responses, including increasing macrophage and neutrophil bactericidal capacity, promoting the recruitment of neutrophils towards the site of infection in the pathophysiology of bovine mastitis, inducing the maturation of dendritic cells, and controlling the acquired immune response (Oviedo-Boyso et al. 2007). IL6 is the keystone cytokine and modulates almost every aspect of the innate immune system; it promotes an alternatively activated macrophage phenotype associated with wound healing and inhibits the microbicidal activities of macrophages and the production of pro-inflammatory cytokines (Hunter and Jones 2015a).
Escherichia coli and Staphylococcus aureus pathogens can rapidly enhance IL6 expression (Günther et al. 2011). Interleukin-6 receptor-alpha (IL6R), a receptor of IL6, was upregulated in mastitic udder tissues challenged by bovine E. coli 24 h postinfection (Buitenhuis et al. 2011). The expression level of IL6R increased by 17-fold in lipopolysaccharide-challenged human mammary epithelial cells (Wang et al. 2013a). By contrast, IL6R expression was downregulated by 3.53-fold in bovine monocyte-derived macrophages infected with S. aureus in vitro 6 h postinfection (Lewandowska-Sabat et al. 2013). Thus far, the molecular mechanism underlying IL6R expression in dairy cattle remains unknown.
Alternative gene splicing is a universal phenomenon that enables a single coding gene to generate many products with similar or different functions (Brett et al. 2001). Several important diseases have been linked with mutations or variations in either cis-acting elements or trans-acting factors that lead to aberrant splicing and abnormal protein production (Garcia-Blanco et al. 2004). These variants, such as SNPs, affect gene function and induce a disease phenotype because the splice mutation shifts the protein reading frame. Commonly observed alternative splicing events preserve the reading frame and minimally affect the protein function (Resch et al. 2004). Our previous studies showed that several immune-related genes undergo alternative splicing events, and SNPs located in splicing-related domains influence susceptibility to bovine mastitis (Wang et al. 2014; Ju et al. 2015; Zhang et al. 2015). However, individuals with the same genetic background may develop different levels of disease severity; this variation cannot be explained by genetics alone; one of the many potential nongenetic sources of variation is epigenetic modification (Runyon et al. 2012; Tryndyak et al. 2016).
Epigenetic modification includes acetylation and methylation of histone and methylation of long noncoding RNAs and DNA (Breiling and Lyko 2015; Larriba and del Mazo 2016; Messier et al. 2016). Of these processes, DNA methylation is the most widely studied because it plays an important role in genome-wide pre-transcriptional regulation (Song et al. 2016), such as alternative splicing (Yearim et al. 2015; Lev Maor et al. 2015). DNA methylation is altered by bacteria (Benakanakere et al. 2015) or inflammation caused by infection (Niwa et al. 2010). Researchers determined that bovine mastitis increased the methylation levels in the promoter region of CD4 and decreased the expression level of CD4 by inhibiting the binding of transcription factors (Wang et al. 2013b). Moreover, mastitis caused by E. coli induced the loss of promoter methylation and increased the expression level of the TLR4 gene (Chang et al. 2015). These studies confirmed the role of DNA methylation in innate immune responses in the process of mastitis. Perturbation of DNA methylation leads to the aberrant splicing of alternatively spliced exons (Yearim et al. 2015; Maunakea et al. 2013). Thus, the generation of new splice variants can be influenced by both genetic and epigenetic mechanisms.
This study aims to determine (1) whether various splice variants of IL6R are commonly observed in the mammary glands of normal and mastitic cows, (2) whether genetic variants that affect splicing patterns are located within the splicing sites, and (3) whether epigenetic modification regulates the expression and alternative splicing patterns of IL6R.
Materials and methods
Animal and tissue samples
Mammary tissue samples were collected from three normal and three mastitis-infected first-lactating Holstein cows obtained from a commercial bovine slaughterhouse in Jinan, Shandong Province, China. The cows were classified into normal and mastitic groups based on clinical symptoms and pathogenic bacteria from the milk culture test. The normal cows did not present any clinical symptoms, such as heat, pain, redness, swelling of the udder, or milk clotting, and were not infected by pathogens. Clinically diagnosed mastitis cows were identified based on these clinical signs. The milk isolates of bovine mastitis mammary areas and the biopsies of mammary gland tissues were tested for pathogenic bacteria through cell culture (Wang et al. 2014; Ju et al. 2015). A total of 18 tissue samples (three for each cow) were immediately frozen in liquid nitrogen. DNA was extracted from six tissue samples by using a TIANamp Genomic DNA Kit (Tiangen, Cat. #DP304) and stored at − 20 °C. Total RNA was extracted from six mammary tissues by using an RNAsimple Total RNA Kit (Tiangen, Cat. #DP419) according to the manufacturer’s instructions. RNA concentration was determined with a Nanophotometer (P330; IMPLEN). RNA and DNA quality was monitored by visualization of ethidium bromide-stained bands in 1% agarose gels after electrophoresis. The RNA samples were stored at − 80 °C.
Sixty-eight Holstein bulls (aged between 1 and 3 years old) with genomic estimated breeding values (gEBVs) were selected from Shandong OX Livestock Breeding Co., Ltd. These bulls were unrelated individuals, which were specially chosen according to the pedigrees. The bulls were obtained by embryo transfer of frozen embryo from North America and genotyped by the bovine GH2 SNP chips (Illumina, GeneSeek 77k chip, version 2), and their genetic evaluations are completed by the Holstein Association USA (http://www.holsteinusa.com/). A gEBV includes mastitis-related traits, such as somatic cell score (SCS) and udder conformation, and may be used for association analysis. Semen samples were collected from the 68 bulls, and DNA was extracted according to the protocol in our previous study (Gao et al. 2014). All DNA samples were stored at − 20 °C for subsequent analysis.
Identification of IL6R novel splice variant
RNA samples were reverse transcribed into cDNA with the PrimeScript™ II First-strand cDNA Synthesis Kit (TaKaRa, Cat. #6210A). RT-PCR analysis was performed with specific primers of IL6R-cDNA (Additional file: Table S1) to amplify the coding region of the IL6R gene by using 2× Taq Master mix (Generay, Cat. #GK8006) according to the manufacturer’s instructions. After electrophoresis, the products were purified with a TIANgel Midi Purification Kit (Tiangen, Cat. #DP209), cloned into the pEASY-T3 vector (Transgene, Cat. #CT301), and transformed into Trans5α (Transgene, Cat. #CD201). Positive clones were directly sequenced by a commercial sequencing company (BGI). The results were compared with the reference IL6R mRNA sequence (GenBank, NM_001110785.1) by using DNAMAN software. Clustal OMEGA (http://www.ebi.ac.uk/Tools/msa/clustalo/) was applied to align the sequences.
Expression analysis of splice variants
Quantitative real-time PCR (qRT-PCR) analysis was conducted to investigate the relative expression of IL6R transcripts in normal and mastitis-infected bovine mammary gland tissues. Analysis was performed using SYBR® Premix Ex Taq™ (TliRNaseH Plus) (TaKaRa, Cat. #RR420A) on a Roche LightCycler® 480II machine (Roche Applied Science). The qRT-PCR protocol and calculation of relative expression were described in our previous study (Ju et al. 2015). Two pairs of specific primers IL6R-G and IL6R-TV (Additional file: Table S1) were used to quantify the expression of IL6R-reference mRNA and the novel splice variant IL6R-TV mRNA. The housekeeping gene β-actin (Additional file: Table S1) was used as reference to normalize data (Hou et al. 2012). The template that did not undergo the reverse transcription reaction served as negative control. All qRT-PCRs were performed in triplicate. Inclusion levels of exon 2 were calculated by dividing the amount of IL6R-reference by the amount of IL6R-TV (Yearim et al. 2015). The relative expression of the two splice variants (IL6R-reference and IL6R-TV) and the relative exon inclusion level were compared between the normal and mastitis-infected cows by using two-tailed, unpaired Student’s T test. Differences with P < 0.05 were considered statistically significant.
Screening, genotyping of SNPs, and statistical analysis
The fragments encompassing the exon 2 of IL6R were amplified using the specific primer EXON2 (Additional file: Table S1). The PCR products of 68 semen samples and six cows were sequenced. Two SNPs located upstream of exon2 were in complete linkage disequilibrium. The two SNPs formed two haplotypes (T-G and C-C) in the studied bulls. SHEsis software (http://analysis.bio-x.cn/myAnalysis.php) was used to analyze the pairwise linkage disequilibrium and haplotypic frequency (Gao et al. 2014). The association between genotypes and gEBVs was tested by Student’s T test. Values with P < 0.05 were considered significant.
Splicing mini-gene reporter assay
Construction of vectors
The 804-bp genomic fragments spanning 369 bp of intron 1, 249 bp of exon 2, and 186 bp of intron 2, which harbored the two SNPs, were amplified using the specific primer E2 (Additional file: Table S1). The mini-gene reporter assays were constructed by cloning the 804-bp fragments with the wild type (g.19711 T > C and g.19731 G > C; wild haplotype: T-G) or the mutant type (g.19711 T > C and g.19731 G > C; mutant haplotype: C-C) into the empty pSPL3 vector after digestion with EcoRI and XhoI. The constructed vectors and the empty vector were transformed into Trans5α. The clones were cultured on agar containing 100 mg/mL ampicillin and incubated overnight at 37 °C. Positive clones were chosen and incubated overnight at 37 °C in 10 mL of lysogeny broth medium containing 100 mg/mL ampicillin. Plasmids were isolated with the EZNA Endo-Free Plasmid DNA Midi Kit (OMEGA, Cat. #D6915) and then measured by a NanoPhotometer (P330; IMPLEN). The plasmids were sequenced after isolation to guarantee the authenticity and orientation of the vectors.
Cell culture
Bovine mammary epithelial cells (MAC-T) and human embryonic kidney 293 T cells (HEK293T) were used in our experiments. Dulbecco’s modified Eagle medium/Ham’s F-12 medium (DMEM-F12) (GIBCO, Cat. #1699748) was used as the basic medium for MAC-T, and Dulbecco’s modified Eagle medium (DMEM) (GIBCO, Cat. #1744353) was used as the basic medium for HEK293T. The growth medium comprised the basic medium supplemented with 500 U/L penicillin, 500 mg/L streptomycin (GIBCO, Cat. #15140163), and 10% (v/v) fetal bovine serum (GIBCO, Cat. #16000044). Both cells were maintained at 37 °C with 5% CO2 and subcultured every other day.
Transfection
MAC-T and HEK293T were used for transfection. The cells were cultured in six-well culture plates. One day before the transfection, the medium was replaced by removing the antibiotics. When the cells reached 80% confluence, 4 μg of the mini-gene reporters or pSPL3 vector was added to the Lipofectamine 2000 Transfection Reagent (Invitrogen, Cat. #11668030) in Opti-MEM medium (Gibco, Cat. #51985042). A growth medium without antibiotics was substituted for the Opti-MEM medium after 5 h. The cells were harvested 24 h after transfection, and total RNA was extracted with the RNAsimple Total RNA Kit (Tiangen, Cat. #DP419) to determine the expression of mini-gene reporters.
Expression analysis of splice variants
RNA extracted from the transfected cells was reverse transcribed into cDNA by using the pSPL3 vector-specific primer SA2 with the PrimeScript™ II First-strand cDNA Synthesis Kit (TaKaRa, Cat. #6210A). Specific primers for pSPL3 vector SD6 and SA2 were used to perform RT-PCR as previously described. The products were separated through electrophoresis on 1% agarose gels. Each DNA band was further analyzed by direct sequencing after extraction with the TIANgel Midi Purification Kit (Tiangen, Cat. #DP209).
Analysis and prediction of CpG island
The online software Methprimer (http://www.urogene.org/cgi-bin/methprimer/ methprimer.cgi) was used to detect and analyze the CpG island located within exon 2 of the IL6R gene.
Bisulfite treatment
A total of 200 ng of genomic DNA from the six mammary tissue samples collected from three normal and three mastitis-infected cows was used for bisulfate modification with a BisulFlash DNA Modification Kit (Epigentek, Cat. #P-1026) according to the manufacturer’s instructions. DNA was stored at − 20 °C until use as templates for bisulfate sequencing PCR (BSP).
Nested BSP
Two special pairs of primer (BSP-F2/R2 for the first step and BSP-F1/R1 for the second step) were designed using Methyl Primer Express Software v1.0. PCR analysis was conducted using 5 μL of 2× GC buffer (5 mM Mg2+ Plus, TaKaRa, Cat. #9154), 1.6 μL of 2.5 mM dNTP mixture (TaKaRa, Cat. #4030), 0.1 μL of TaKaRa LA Taq (5 U/μL, TaKaRa, Cat. #RR52A), 0.5 μL of each primer (10 mM), and 0.5 μL of template DNA in a total volume of 10 μL. The first step of PCR was performed as follows: (1) 94 °C denaturation for 5 min, (2) 16 thermal cycles of denaturation at 94 °C for 30 s, (3) annealing starting at 56 °C and reducing 1 °C every cycle for 30 s, (4) extension at 72 °C for 1 min, and (5) another 29 cycles of denaturation at 94 °C for 30 s, 41 °C for 30 s, and 72 °C for 1 min. Final extension was conducted for 10 min at 72 °C. The products of the first step were used as templates for the second step, which was conducted as follows: (1) 94 °C denaturation for 5 min, (2) followed by 16 thermal cycles of denaturation at 94 °C for 30 s, (3) annealing starting at 55 °C and reducing 1 °C every cycle for 30 s, (4) extension at 72 °C for 1 min, and (5) another 29 cycles of denaturation at 94 °C for 30 s, 40 °C for 30 s, and 72 °C for 1 min. Final extension was performed for 10 min at 72 °C. The PCR products were purified, cloned into pEASY-T3 vectors, and transformed into Trans5α for sequencing. Sequences with > 95% cytosine conversion were analyzed through BiQ Analyzer software with the reference IL6R gene sequence.
Results
Identification of a novel bovine IL6R splice variant
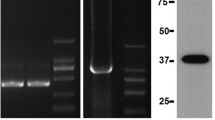
Six mammary tissues of dairy cows (three healthy cows and three mastitic ones) were used to amplify the entire coding region of the reference IL6R gene. In addition to the predicted 664-bp PCR product, a smaller band (415 bp) was detected (Fig. 1a). After the cloning and sequencing of the smaller band, a new transcript, named IL6R-TV, was found in both normal and mastitic mammary gland tissues. The novel IL6R-TV (415 bp) was characterized by the skipping of the entire exon 2 (Fig. 1b); therefore, the exon 2 of the IL6R gene was considered an alternatively spliced exon.

RT-PCR products from the IL6R gene expressed in bovine mammary tissues and schematic representation of the splicing patterns of IL6R transcripts and SNPs (g.19711 T > C and g.19731 G > C): a RT-PCR products from the IL6R gene expressed in bovine mammary tissues; lane 1–lane 6: RT-PCR products of different cows. TT-GG and TC-GC indicated the genotype combination of the cows. H-MG, healthy mammary glands. M-MG, mastitis mammary glands. b Genomic structure of the bovine IL6R gene and IL6R-TV. The IL6R-TV splice variant lacks the sequence of exon 2. Primers E2F and E2R span exon 2 and are used to amplify fragments to screen the existence of SNPs. The position of the A nucleotide in the start codon (ATG) is defined as +1
Bioinformatic analysis suggested that the IL6R-reference and IL6R-TV proteins shared the same signal peptide and transmembrane domain. In contrast to the IL6R-reference protein, the putative IL6R-TV protein lost 83 amino acids encoding an immunoglobulin-like domain (Additional file: Fig. S1).
Analysis of the expression of the two transcripts in mammary tissues
Quantitative real-time PCR (qRT-PCR) results showed that the two transcripts were expressed in the mammary tissues of normal and mastitic cows; the IL6R-reference was found to be the primary transcript in the mammary gland tissues (Fig. 2a).The expression levels of IL6R-reference and IL6R-TV in the normal mammary tissues were significantly lower than those in the mastitic tissues (P < 0.05). The inclusion levels of exon 2 were significantly higher in the mammary tissues of mastitis-infected cows than those in normal cows (P < 0.05) (Fig. 2b).

Expression levels of different transcripts of IL6R and the relative exon inclusion levels between normal and mastitis-infected groups. a qRT-PCR of IL6R-reference and IL6R-TV of normal and mastitis-infected cows. One asterisk, P < 0.05, significantly different from normal controls. Two asterisks, P < 0.01, significantly different from normal controls. b The relative exon inclusion level was calculated by dividing the amount of the inclusion isoform with the amount of the skipping isoform. Asterisk, P < 0.05, significantly different from normal controls
Putative effect of SNPs on splicing in exonic splicing enhancer motif
We amplified the genomic region from exon 1 to exon 3 of the bovine IL6R-reference gene by using specific primers EXON2. We directly sequenced the PCR products with specific primer E2F and E2R in 68 bulls and six cows and found two SNPs (g.19711 T > C and g.19731 G > C), which were located 85 and 65 bp upstream of exon 2 of IL6R, respectively (Fig. 1a), and within the exonic splicing enhancer (ESE) motif region. ESEfinder 3.0 (http://www.mybiosoftware.com/esefinder-3-0-exon-splicing-enhancer-finder.html) was applied to identify exonic splicing enhancer motifs for wild and mutant sequences. Two binding sites for the splicing factors SF2/ASF and SRp40 diminished, and one binding site for the splicing factor SC35 was added to the mutant (Additional file: Figs. S2 and S3). Genetic mutations can putatively affect the binding of the splicing factors. However, the effect of changes in the binding of splicing factors on the splicing patterns of IL6R was not determined in this study.
Cell culture and transfection
The SV-40 T transformed bovine MAC-T cell line was an easy-to-handle model for the analysis of immune functions of mammary epithelial cells (Günther et al. 2016). So MAC-T was applied in our experiments; HEK293T was chosen as the reference. We constructed two IL6R mini-genes which comprised the wild and mutant haplotypes of the two SNPs and transfected them into two cell lines, namely MAC-T and HEK293T, respectively, to investigate their effects at the splicing level and investigate whether SNP genotypes directly define the patterns of the spliced isoforms (Fig. 3).The RT-PCR results of the splicing assay showed that both wild haplotype (T-G) and mutant haplotype (C-C) mini-gene vectors produced a 512-bp fragment, which included the 249 bp of exon 2 and 263 bp of pSPL3. The empty pSPL3 vector generated a 263-bp fragment and a nonspecific binding fragment in MAC-T (Fig. 3d) and HEK293T (Fig. 3e). In both cells, the small RT-PCR product corresponding to the mini-genes of the two haplotypes was directly sequenced and showed nonspecific binding. Both the wild and mutant haplotype reporters did not generate the predicted 263-bp fragment (Fig. 3d, e). This finding indicated that the two haplotypes did not produce the novel splice variant IL6R-TV characterized by deleting the entire exon 2.

Two haplotypes generated the same splicing pattern of IL6R in MAC-T and HEK293T: a protocol of splicing mini-gene reporter assay. b Microscopic images of MAC-T before and after cell transfection with two IL6R mini-genes (amplification, 10 × 20). c Microscopic images of HEK293T before and after cell transfection with two IL6R mini-genes (amplification, 10 × 20). d Splicing mini-gene reporter assay in MAC-T. e Splicing mini-gene reporter assay in HEK293T. The wild haplotype (g.19711 T > C-T and g.19731 G > C-G; haplotype: T-G) and the mutant type (g.19711 T > C-C and g.19731 G > C-C; haplotype: C-C) mini-gene reporters give rise to a 512-bp fragment, which consists of exon 2 and the 263-bp fragment of pSPL3 vector in the MAC-T and HEK293T. Nonspecific amplification products were visible and sequenced
Association between two SNPs and the gEBV of the SCS as well as udder conformation
Somatic cell count (SCC) and SCS are used as indicators of dairy mastitis (Wang et al. 2014). Lund et al. (Lund et al. 1994) showed that improved udder conformation reduced the SCC and the occurrence of clinical mastitis. SCC, SCS, and udder conformation can be considered as indicators of dairy mastitis. Sixty-eight bull samples were used to amplify the fragments containing the two SNPs, and the bulls were genotyped through sequencing to determine the association of the SNPs with mastitis in dairy cows. The linkage disequilibrium between the two SNPs in the population was complete: there were only two haplotypes, namely T-G and C-C. The two haplotypes resulted in three two-locus genotypes, namely TT-GG, TC-GC, and CC-CC, with frequencies 60.3, 38.2, and 1.5%, respectively. The relationship between the three genotypes and mastitis indicators, including gEBVs of SCS and seven udder traits, was analyzed. Since only one sample of genotype CC-CC was detected, this genotype was not included in the association analysis. The results indicated that the two SNPs were not significantly associated with SCS (P > 0.05); hence, the two SNPs might not be used as the markers for resistance to mastitis (Table 1). More samples are needed to further investigate their correlation.
Analysis of DNA methylation status
DNA methylation affects pre-mRNA splicing by influencing the recognition of an alternatively spliced exon (Yearim et al. 2015; Maunakea et al. 2013). In the present study, the second exon and intron of IL6R which spanned one CpG island was identified by Methprimer (http://www.urogene.org/cgi-bin/methprimer/methprimer.cgi) (Fig. 4a). The result indicated the presence of epigenetic regulation by ASE methylation. Nested BSP and touchdown PCR analyses were used to amplify the 233 bp of the CpG island with 13 CpG sites. Sixty clones (10 clones for each cow) were selected for analysis. The methylation rate significantly differed and was found to be higher in the mammary gland tissues of mastitis-infected cows than in normal cows (Fig. 4b, c). Further analysis showed that the methylation levels of the 3rd, 6th, 8th, 9th, and 10th CpG sites were significantly higher in mastitic cows than in normal cows (Fig. 4d).

Methylation profiles of exon 2 and intron 2 of IL6R in mammary tissues. a Schematic of the predicted CpG island and the location of the BSP primers. A strong CpG island maps within exon 2 of IL6R. The red lines represent 13 CpG sites. b Percentage of methylation of mammary gland tissues in normal (1–3) and mastitis-infected (4–6) cows. c The frequency of DNA methylation of the predicted CpG island was lower in normal cows (55.40%, n = 30 clones) than in mastitis-infected cows (84.83%); asterisk, P < 0.05, significantly different from normal controls. Results of BSP of normal (animal #1, 2, and 3) and mastitis-infected (animal #4, 5, and 6) groups. A 233-bp fragment with 13 CpG sites was amplified by using nested BSP. A total of 60 clones (10 for each cow) that had sequences with > 95% cytosine conversion were analyzed through BiQ Analyzer software with the reference IL6R gene sequence. A black circle represents methylated CpG sites; a blank circle represents unmethylated CpG sites. d Percentage of methylation of the 13 CpG sites of normal and mastitis-infected groups. The methylation levels were higher in the mastitis-infected cows than in normal cows at all the 13 CpG sites, particularly at CpG_3, CpG_6, CpG_8, CpG_9, and CpG_10. One asterisk, P < 0.05, significantly different from normal controls. Two asterisks, P < 0.01, significantly different from normal controls
Discussion
IL6 has long been recognized as a prototypic pro-inflammatory cytokine that is involved in the pathogenesis of all inflammatory diseases (Garbers et al. 2015). IL6 acts on the immune system by regulating the development and activation status of innate and adaptive immune cells (Fontes et al. 2015). As the receptor of IL6, IL6R stimulates various cellular responses, including inflammatory processes, to initiate intracellular signaling pathways by participating in the formation of the signal transducing complex IL-6/IL6R/gp130 (Hunter and Jones 2015b; Kumar et al. 2015). IL6 first binds to IL6R and forms a complex, which binds to gp130 to initiate different pathways, such as those of MAPK-C/EBPβ and JAK/STAT (Kumar et al. 2015). Human IL6R, which exists in membrane-bound (mIL6R) and soluble (sIL6R) forms, plays critical roles in eliciting immune responses (Wang et al. 2013a; Garbers et al. 2015; Lopez-Lasanta et al. 2015). This protein is expressed on normal hepatocytes, CD4+ or CD8+ T cells (Jones et al. 2011), inflammatory cells, and cancer cells (Zhang et al. 2013; Fenton et al. 2000). Human sIL6R is one of the IL6R transcripts partly generated by the alternative mRNA splicing of the IL6R gene by skipping exon 9 (Li et al. 2015). The IL6R gene, also known as IL6Rα, CD126, and gp80 gene, possesses other splice variants generated through alternative or aberrant splicing in other species, such as mice and pigs (http://ensemblgenomes.org/). Thus, IL6R expression and its regulation should be investigated to elucidate its potential role in bovine mastitis.
In this study, a novel IL6R splice variant without the entire exon2 of the IL6R-reference gene, named IL6R-TV, was identified. This study is the first to report a distinct alternative splicing pattern of the bovine IL6R gene. Furthermore, two transcripts, IL6R-reference and IL6R-TV, were high-regulated in the mastitic tissues. IL6R was upregulated in E. coli-challenged bovine udder tissues 24 h postinfection (Buitenhuis et al. 2011) and in lipopolysaccharide-challenged human mammary epithelial cells at the early stage of infection (Wang et al. 2013a). IL6R was downregulated by 3.53-fold in bovine monocyte-derived macrophages infected with S. aureus in vitro 6 h postinfection (Lewandowska-Sabat et al. 2013). In the present study, the expression levels of IL6R-reference and IL6-TV increased in the mammary tissues of mastitic cows, suggesting their potential role against pathogenic infection.
Inflammation is a normal and essential immunological process to defend the host against pathogens. Host inflammatory responses eliminate the source of tissue injury, restore immune homeostasis, and return affected mammary glands to normal function. Imbalance between pro-inflammatory and pro-resolving mechanisms is needed to ensure optimal bacterial clearance and induce return to immune homeostasis (Aitken et al. 2011). Therefore, changes of the IL6R expression level are necessary in the process of inflammation. The IL6R gene may be considered as a candidate gene for mastitis susceptibility.
Numerous SNPs are located in splicing-relevant sequences, such as canonical splice-site consensus sequences, including ESE, intronic splice enhancer, exonic splicing silencer, and intronic splicing silencer (ElSharawy et al. 2006). An increasing body of evidence demonstrated the functional importance of alternative splice variations and splicing-related SNPs within intronic and exonic motif sequences in mastitis (Wang et al. 2014; Ju et al. 2015; Zhang et al. 2015; Li et al. 2013). Thus, we searched genetic mutation within the close gene sequences around IL6R splicing sites. The results revealed two SNPs (g.19711 T > C and g.19731 G > C) in the upstream of the missed exon 2. Analysis of bioinformatic prediction suggested that the two SNPs were located in the ESE motif and influenced the target region-binding capability of splicing-related factors. The mutant alleles abolished two binding sites for the splicing factors SF2/ASF and SRp40 and increased one binding site for the splicing factor SC35. The serine/arginine family of proteins constitutes an essential class of splicing regulators through splice-site selection. Mammalian SF2/ASF binds to a purine-rich motif and coordinates with SRSF2 to regulate exon inclusion and skipping (Bradley et al. 2015). SRp40 and SC35 splicing factors are associated with the formation of gene splice variants characterized by variable exons (Bradley et al. 2015; Huang et al. 2007). Through in silico prediction, we initially speculated that the two SNPs might be the source of aberrant splice variant IL6R-TV. Therefore, a pSPL3 exon-trapping system was used to test this hypothesis. We constructed two mini-gene vectors with different haplotypes and transfected them separately into MAC-T and HEK293T. Our findings indicated that changes in ESE binding sequences did not enhance the splicing pattern of IL6R; hence, the two SNPs did not induce the production of IL6R-TV. Moreover, the statistical analysis of sperm samples showed that the two SNPs were not associated with gEBVs of SCS and udder conformation traits. By combining the results of correlation analysis and the exon-trapping system, we concluded that production of aberrantly spliced IL6R-TV was not by the two SNPs but through other means.
DNA methylation is an important epigenetic marker that significantly affects natural variation (Heyn et al. 2013). The DNA methylation level in the gene promoter is a powerful epigenetic marker for clinical mastitis (Buitenhuis et al. 2011) and acute mastitis induced by E. coli (Vanselow et al. 2006). Similar to that in gene bodies, DNA methylation is positively correlated with gene expression (Yang et al. 2014) and inclusion levels of alternative exons (Maunakea et al. 2013) by affecting the kinetics of transcriptional elongation or by recruiting special splicing factors (Yearim et al. 2015; Fenton et al. 2000; Iannone and Valcárcel 2013). Previous research showed that DNA methylation regulates gene expression, and the present findings indicated that the CpG island was located within alternative exon 2; hence, we speculate that the expression levels of the transcripts of IL6R and the inclusion levels of the alternatively spliced exon 2 were regulated by DNA methylation. Our BSP and qRT-PCR results confirmed the hypothesis. Mastitis enriched DNA methylation on the exon 2 of IL6R. The high DNA methylation levels upregulated the expression of IL6R-reference and IL6R-TV and the inclusion levels of exon 2. These results proved that DNA methylation was in correlation with alternative splicing patterns of IL6R during inflammatory infection.
Conclusions
We identified one novel IL6R transcript that skipped exon 2, and it is upregulated in the mammary gland tissue of clinical mastitis cows. Bovine mastitis increased the DNA methylation level, particularly on some distinct CpG sites, and influenced the expression levels of both inclusion and skipping isoforms and their ratios, resulting in the increased inclusion level of exon 2. DNA methylation rather than SNP regulates the production of an aberrant splice variant of IL6R in mastitic cows. The DNA methylation level in exon 2 could be a potential biomarker for monitoring bovine mastitis.
Abbreviations
- IL6R:
-
Interleukin-6 receptor-alpha
- SNPs:
-
Single nucleotide polymorphisms
- MAC-T:
-
Bovine mammary epithelial cells
- HEK293T:
-
Human embryonic kidney 293 T cells
- BSP:
-
Bisulfate sequencing PCR
- E. coli :
-
Escherichia coli
- S. aureus :
-
Staphylococcus aureus
- ASEs:
-
Alternatively spliced exons
- TPI:
-
Total performance index
- SCS:
-
Somatic cell score
- Q-PCR:
-
Quantitative real-time PCR
- DMEM-F12:
-
Dulbecco’s modified Eagle medium/Ham’s F-12 medium
- DMEM:
-
Dulbecco’s modified Eagle medium
- TFBS:
-
Transcriptional factor binding sites
- ESE:
-
Exonic splicing enhancer
- DMRs:
-
Differentially methylated regions
- ETS family:
-
E26 transformation-specific family
- mIL6R:
-
Membrane-bound interleukin-6 receptor-alpha
- sIL6R:
-
Soluble interleukin-6 receptor-alpha
- SR family:
-
Serine/arginine (SR) family
References
Aitken SL, Corl CM, Sordillo LM (2011) Immunopathology of mastitis: insights into disease recognition and resolution. J Mammary Gland Biol Neoplasia 16(4):291–304. https://doi.org/10.1007/s10911-011-9230-4
Benakanakere M, Adbolhosseini M, Hosur K, Finoti LS, Kinane DF (2015) TLR2 promoter hypermethylation creates innate immune dysbiosis. J Dent Res 94(1):183–191. https://doi.org/10.1177/0022034514557545
Bradley T, Cook ME, Blanchette M (2015) SR proteins control a complex network of RNA-processing events. RNA 21(1):75–92. https://doi.org/10.1261/rna.043893.113
Breiling A, Lyko F (2015) Epigenetic regulatory functions of DNA modifications: 5-methylcytosine and beyond. Epigenetics Chromatin 8(24). https://doi.org/10.1186/s13072-015-0016-6
Brett D, Pospisil H, Valcárcel J, Reich J, Bork P (2001) Alternative splicing and genome complexity. Nat Genet 30:29–30
Buitenhuis B, Røntved CM, Edwards SM, Ingvartsen KL, Sørensen P (2011) In depth analysis of genes and pathways of the mammary gland involved in the pathogenesis of bovine Escherichia coli-mastitis. BMC Genomics 12(130). https://doi.org/10.1186/1471-2164-12-130
Chang G, Petzl W, Vanselow J, Günther J, Shen X, Seyfert HM (2015) Epigenetic mechanisms contribute to enhanced expression of immune response genes in the liver of cows after experimentally induced Escherichia coli mastitis. Vet J 203(3):339–341. https://doi.org/10.1016/j.tvjl.2014.12.023
ElSharawy A, Manaster C, Teuber M, Rosenstiel P, Kwiatkowski R, Huse K, Platzer M, Becker A, Nürnberg P, Schreiber S, Hampe J (2006) SNPSplicer: systematic analysis of SNP-dependent splicing in genotyped cDNAs. Hum Mutat 27(11):1129–1134. https://doi.org/10.1002/humu.20377
Fontes JA, Rose NR, Čiháková D (2015) The varying faces of IL-6: from cardiac protection to cardiac failure. Cytokine 74(1):62–68. https://doi.org/10.1016/j.cyto.2014.12.024
Gao Q, Ju Z, Zhang Y, Huang J, Zhang X, Qi C, Li J, Zhong J, Li G, Wang C (2014) Association of TNP2 gene polymorphisms of the bta-miR-154 target site with the semen quality traits of Chinese Holstein bulls. PLoS One 9(1):e84355. https://doi.org/10.1371/journal.pone.0084355
Garbers C, Aparicio-Siegmund S, Rose-John S (2015) The IL-6/gp130/STAT3 signaling axis: recent advances towards specific inhibition. Curr Opin Immunol 34:75–82. https://doi.org/10.1016/j.coi.2015.02.008
Garcia-Blanco MA, Baraniak AP, Lasda EL (2004) Alternative splicing in disease and therapy. Nat Biotechnol 22(5):535–546. https://doi.org/10.1038/nbt964
García-Ruiz A, Cole JB, VanRaden PM, Wiggans GR, Ruiz-López FJ, Van Tassell CP (2016) Changes in genetic selection differentials and generation intervals in US Holstein dairy cattle as a result of genomic selection. Proc Natl Acad Sci U S A 113(28):E3995–E4004. https://doi.org/10.1073/pnas.1519061113
Gomes F, Henriques M (2016) Control of bovine mastitis: old and recent therapeutic approaches. Curr Microbiol 72(4):377–382. https://doi.org/10.1007/s00284-015-0958-8
Günther J, Esch K, Poschadel N, Petzl W, Zerbe H, Mitterhuemer (2011) Comparative kinetics of Escherichia coli- and Staphylococcus aureus-specific activation of key immune pathways in mammary epithelial cells demonstrates that S. aureus elicits a delayed response dominated by interleukin-6 (IL-6) but not by IL-1A or tumor necrosis factor alpha. Infect Immun 79(2):695–707. https://doi.org/10.1128/IAI.01071-10
Günther J, Koy M, Berthold A, Schuberth HJ, Seyfert HM (2016) Comparison of the pathogen species-specific immune response in udder derived cell types and their models. Vet Res 47(1, 22)
Heyn H, Moran S, Hernando-Herraez I, Sayols S, Gomez A, Sandoval J, Monk D, Hata K, Marques-Bonet T, Wang L, Esteller M (2013) DNA methylation contributes to natural human variation. Genome Res 23(9):1363–1372. https://doi.org/10.1101/gr.154187.112
Hinchs D, Bennewitz J, Stamer E, Junge W, Kalm E, Thaller G (2011) Genetic analysis of mastitis data with different models. J Dairy Sci 94(1):471–478. https://doi.org/10.3168/jds.2010-3374
Hou Q, Huang J, Ju Z, Li Q, Li L, Wang L et al (2012) Identification of splice variants, targeted microRNAs and functional SNPs of the BOLA-DQA2 gene in dairy cattle. DNA Cell Biol 31(5):739–744. https://doi.org/10.1089/dna.2011.1402
Huang CS, Shen CY, Wang HW, Wu PE, Cheng CW (2007) Increased expression of SRp40 affecting CD44 splicing is associated with the clinical outcome of lymph node metastasis in human breast cancer. Clin Chim Acta 384(1–2):69–74. https://doi.org/10.1016/j.cca.2007.06.001
Huang J, Luo G, Zhang Z, Wang X, Ju Z, Qi C, Zhang Y, Wang C, Li R, Li J, Yin W, Xu Y, Moisá SJ, Loor JJ, Zhong J (2014) iTRAQ—proteomics and bioinformatics analyses of mammary tissue from cows with clinical mastitis due to natural infection with Staphylococci aureus. BMC Genomics 15(1):839. https://doi.org/10.1186/1471-2164-15-839
Hunter CA, Jones SA (2015a) IL-6 as a keystone cytokine in health and disease. Nat Immunol 16(5):448–457. https://doi.org/10.1038/ni.3153
Hunter CA, Jones SA (2015b) IL-6 as a keystone cytokine in health and disease. Nat Immunol 16(5):448–457. https://doi.org/10.1038/ni.3153
Iannone C, Valcárcel J (2013) Chromatin’s thread to alternative splicing regulation. Chromosoma 122(6):465–474. https://doi.org/10.1007/s00412-013-0425-x
Jones SA, Scheller J, Rose-John S (2011) Therapeutic strategies for the clinical blockade of IL-6/gp130 signaling. J Clin Invest 121(9):3375–3383. https://doi.org/10.1172/JCI57158
Ju Z, Wang C, Wang X, Yang C, Sun Y, Jiang Q, Wang F, Li M, Zhong J, Huang J (2015) Role of an SNP in alternative splicing of bovine NCF4 and mastitis susceptibility. PLoS One 10(11):e0143705. https://doi.org/10.1371/journal.pone.0143705
Kumar H, Raj U, Gupta S, Tripathi R, Varadwaj PK (2015) Systemic review on chronic myeloid leukemia: therapeutic targets, pathways and inhibitors. J Nucl Med Radiat Ther 6(6)
Larriba E, del Mazo J (2016) Role of non-coding RNAs in the transgenerational epigenetic transmission of the effects of reprotoxicants. Int J Mol Sci 17(4):452. https://doi.org/10.3390/ijms17040452
Lev Maor G, Yearim A, Ast G (2015) The alternative role of DNA methylation in splicing regulation. Trends Genet 31(5):274–280
Lewandowska-Sabat AM, Boman GM, Downing A, Talbot R, Storset AK, Olsaker I (2013) The early phase transcriptome of bovine monocyte-derived macrophages infected with Staphylococcus aureus in vitro. BMC Genomics 14(1):891. https://doi.org/10.1186/1471-2164-14-891
Li L, Huang J, Ju Z, Li Q, Wang C, Qi C, Zhang Y, Hou Q, Hang S, Zhong J (2013) Multiple promoters and targeted microRNAs direct the expressions of HMGB3 gene transcripts in dairy cattle. Anim Genet 44(3):241–250. https://doi.org/10.1111/age.12007
Li H, Rokavec M, Hermeking H (2015) Soluble IL6R represents a miR-34a target: potential implications for the recently identified IL-6R/STAT3/miR-34afeed-back loop. Oncotarget 6(16):14026–14032. https://doi.org/10.18632/oncotarget.4334
Lopez-Lasanta M, Julià A, Maymó J, Fernández-Gutierrez B, Ureña-Garnica I, Blanco FJ, Cañete JD, Alperi-López M, Olivè A, Corominas H, Tornero J, Erra A, Almirall M, Palau N, Ortiz A, Avila G, Rodriguez-Rodriguez L, Alonso A, Tortosa R, Gonzalez-Alvaro I, Marsal S (2015) Variation at interleukin-6 receptor gene is associated to joint damage in rheumatoid arthritis. Arthritis Res Ther 17(242). https://doi.org/10.1186/s13075-015-0737-8
Lund T, Miglior F, Dekkers JCM, Burnside EB (1994) Genetic-relationships between clinical mastitis, somatic-cell count, and udder conformation in Danish Holsteins. Livest Prod Sci 39(3):243–251. https://doi.org/10.1016/0301-6226(94)90203-8
Maunakea AK, Chepelev I, Cui K, Zhao K (2013) Intragenic DNA methylation modulates alternative splicing by recruiting MeCP2 to promote exon recognition. Cell Res 23(11):1256–1269. https://doi.org/10.1038/cr.2013.110
Messier TL, Gordon JA, Boyd JR, Tye CE, Browne G, Stein JL et al (2016) Histoen H3 lysine 4 acetylation and methylation dynamics define breast cancer subtypes. Oncotarget 7(5):5094–5109. https://doi.org/10.18632/oncotarget.6922
Niwa T, Tsukamoto T, Toyoda T, Mori A, Tanaka H, Maekita T et al (2010) Inflammatory processes triggered by Helicobacter pylori infection cause aberrant DNA methylation in gastric epithelial cells. Cancer Res 70(4):11430–11440
Oviedo-Boyso J, Valdez-Alarcón JJ, Cajero-Juárez M, Ochoa-Zarzosa A, López-Meza JE, Bravo-Patiño A et al (2007) Innate immune response of bovine mammary gland to pathogenic bacteria responsible for mastitis. J Inf Secur 54(4):399–409
Rainard P, Cunha P, Gilbert FB (2016) Innate and adaptive immunity synergize to trigger inflammation in the mammary gland. PLoS One 11(4):e0154172. https://doi.org/10.1371/journal.pone.0154172
Rawstron AC, Fenton JA, Ashcroft J, English A, Jones RA, Richards SJ et al (2000) The interleukin-6 receptor alpha-chain (CD126) is expressed by neoplastic but not normal plasma cells. Blood 96(12):3880–3886
Resch A, Xing Y, Alekseyenko A, Modrek B, Lee C (2004) Evidence for a subpopulation of conserved alternative splicing events under selection pressure for protein reading frame preservation. Nucleic Acids Res 32(4):1261–1269. https://doi.org/10.1093/nar/gkh284
Runyon RS, Cachola LM, Rajeshuni N, Hunter T, Garcia M, Ahn R, Lurmann F, Krasnow R, Jack LM, Miller RL, Swan GE, Kohli A, Jacobson AC, Nadeau KC (2012) Asthma discordance in twins is linked to epigenetic modifications of T cells. PLoS One 7(11):e48796. https://doi.org/10.1371/journal.pone.0048796
Song M, He Y, Zhou H, Zhang Y, Li X, Yu Y (2016) Combined analysis of DNA methylome and transcriptome reveal novel candidate genes with susceptibility to bovine Staphylococcus aureus subclinical mastitis. Sci Rep 6(29390)
Tryndyak VP, Han T, Fuscoe JC, Ross SA, Beland FA, Pogribny IP (2016) Status of hepatic DNA methylome predetermines and modulates the severity of non-alcoholic fatty liver injury in mice. BMC Genomics 17(298). https://doi.org/10.1186/s12864-016-2617-2
Vanselow J, Yang W, Herrmann J, Zerbe H, Schuberth HJ, Petzl W, Tomek W, Seyfert HM (2006) DNA-remethylation around a STAT5-binding enhancer in the alphaS1-casein promoter is associated with abrupt shutdown of alphaS1-casein synthesis during acute mastitis. J Mol Endocrinol 37(3):463–477. https://doi.org/10.1677/jme.1.02131
Wang XX, Wu Z, Huang HF, Han C, Zou W, Liu J (2013a) Caveolin-1, through its ability to negatively regulate TLR4, is a crucial determinant of MAPK activation in LPS-challenged mammary epithelial cells. Asian Pac J Cancer Prev 14(4):2295–2299. https://doi.org/10.7314/APJCP.2013.14.4.2295
Wang XS, Zhang Y, He YH, Ma PP, Fan LJ, Wang YC, Zhang YI, Sun DX, Zhang SL, Wang CD, Song JZ, Yu Y (2013b) Aberrant promoter methylation of the CD4 gene in peripheral blood cells of mastitic dairy cows. Genet Mol Res 12(4):6228–6239. https://doi.org/10.4238/2013.December.4.10
Wang X, Zhong J, Gao Y, Ju Z, Huang J (2014) A SNP in intron 8 of CD46 causes a novel transcript associated with mastitis in Holsteins. BMC Genomics 15(1):630. https://doi.org/10.1186/1471-2164-15-630
Wiggans GR, Cole JB, Hubbard SM, Sonstegard TS 2016 Genomic selection in dairy cattle: the USDA experience. Annu Rev Anim Biosci
Yang X, Han H, De Carvelho DD, Lay FD, Jones PA, Liang G (2014) Gene body methylation can alter gene expression and is a therapeutic target in cancer. Cancer Cell 26(4):577–590. https://doi.org/10.1016/j.ccr.2014.07.028
Yearim A, Gelfman S, Shayevitch R, Melcer S, Glaich O, Mallm JP, Nissim-Rafinia M, Cohen AHS, Rippe K, Meshorer E, Ast G (2015) HP1 is involved in regulating the global impact of DNA methylation on alternative splicing. Cell Rep 10(7):1122–1134. https://doi.org/10.1016/j.celrep.2015.01.038
Zhang G, Tsang CM, Deng W, Yip YL, Lui VW, Wong SC, al e (2013) Enhanced IL-6/IL6R signaling promotes growth and malignant properties in EBV-infected premalignant and cancerous nasopharyngeal epithelial cells. PLoS One 8(5):e62284. https://doi.org/10.1371/journal.pone.0062284
Zhang Z, Wang X, Li R, Ju Z, Qi C, Zhang Y et al (2015) Genetic mutations potentially cause two novel NCF1 splice variants up-regulated in the mammary gland, blood and neutrophil of cows infected by Escherichia coli. Microbiol Res 174:24–32
Acknowledgements
We gratefully acknowledge Professor Kerong Shi from Shandong Agricultural University for kindly providing the MAC-T cell line. We thank Dr. Guili Song of the Institute of Hydrobiology, Chinese Academy of Sciences, for providing the pSPL3 vector.
Funding
This work was supported by grants from the National Natural Science Foundation of China (31401049, 31671286 and 31672397), the Agricultural Science and Technology Innovation Program (ASTIP-IA S06), and the Key Research and Development Program of Shandong (2015GNC110002).
Author information
Authors and Affiliations
Corresponding authors
Ethics declarations
All experiments were performed in accordance with the Regulations for the Administration of Affairs Concerning Experimental Animals published by the Ministry of Science and Technology, China, in 2004. The procedures were approved by the Animal Care and Use Committee of the Dairy Cattle Research Center, Shandong Academy of Agricultural Sciences, Shandong, People’s Republic of China.
Electronic supplementary material
ESM 1
(PDF 575 kb)
Rights and permissions
About this article

Cite this article
Zhang, Y., Wang, X., Jiang, Q. et al. DNA methylation rather than single nucleotide polymorphisms regulates the production of an aberrant splice variant of IL6R in mastitic cows. Cell Stress and Chaperones 23, 617–628 (2018). https://doi.org/10.1007/s12192-017-0871-0
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s12192-017-0871-0