
Abstract
Conditioning protocols involving mechanical stress independently or with chemical cues such as growth factors (GFs) possess significant potential to enhance bone regeneration. However, utilization of thermal stress conditioning alone or with GFs for bone therapy has been under-investigated. In this study, a preosteoblast cell line (MC3T3-E1) was exposed to treatment with water bath heating (44°C, 4 and 8 min) and osteoinductive GFs (bone morphogenetic protein-2 and transforming growth factor-β1) individually or in combination to investigate whether these stimuli could promote induction of bone-related markers, an angiogenic factor, and heat shock proteins (HSPs). Cells remained viable when heating durations were less than 20 min at 40ºC, 16 min at 42ºC, and 10 min at 44ºC. Increasing heating duration at 44°C, promoted gene expression of HSPs, osteocalcin (OCN), and osteopontin (OPN) at 8 h post-heating (PH). Heating in combination with GFs caused the greatest gene induction of osteoprotegerin (OPG; 6.9- and 1.6-fold induction compared to sham-treated and GF only treated groups, respectively) and vascular endothelial growth factor (VEGF; 16.0- and 1.6-fold compared to sham and GF-only treated groups, respectively) at 8 h PH. Both heating and GFs independently suppressed the matrix metalloproteinase-9 (MMP-9) gene. GF treatment caused a more significant decrease in MMP-9 protein secretion to non-detectable levels compared to heating alone at 72 h PH. Secretion of OCN, OPN, and OPG increased with the addition of GFs but diminished with heating as measured by ELISA at 72 h PH. These results suggest that conditioning protocols utilizing heating and GFs individually or in combination can induce HSPs, bone-related proteins, and VEGF while also causing downregulation of osteoclastic activity, potentially providing a promising bone therapeutic strategy.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
Bone diseases such as osteoporosis primarily result from abnormal activities of bone cells such as osteoclasts or osteoblasts that cause an imbalance between bone formation and resorption (Teitelbaum 2000). Numerous research groups have investigated the cellular response and regenerative potential of stress conditioning (e.g., tensile and shear stress) to develop novel therapeutic strategies for bone repair (Singh et al. 2007; Sharp et al. 2009). In addition to mechanical stress, in situ delivery of exogenous bone-related growth factors (GFs) such as transforming growth factors (TGFs; Choi et al. 1996; Kandziora et al. 2002), bone morphogenetic proteins (BMPs; Wei et al. 2007; Jeong et al. 2004), and vascular endothelial growth factor (VEGF; Zhang et al. 2009) enhanced osteogenesis and bone healing. In particular, recombinant human BMP-2, an FDA-approved GF, induced osteogenesis of mesenchymal cells into osteoblasts by various transfection factors such as Runx-2 and Osterix (Ryoo et al. 2006). MC3T3-E1 cells exhibited enhanced alkaline phosphatase (ALP) and osteocalcin (OCN) following exogenous delivery of BMP-2 (Spinella-Jaegle et al. 2001) and their proliferation increases following introduction of TGF-β1 (Peterson et al. 2004). VEGF is produced by several cell types (e.g., fibroblasts and osteoblasts) and can induce new blood vessel formation and bone healing when administered exogenously, suggesting that VEGF plays a critical role in bone regeneration (Carano and Filvaroff 2003; Dai and Rabie 2007; Street et al. 2002).
Despite the extensive use of conditioning with mechanical stress and chemical cues in tissue engineering and bone-related research, few studies have investigated thermal stress conditioning as a method for promoting regeneration. Low-level heating in the range of 40–42°C using various heating devices (e.g., water bath, humidified incubator, heating blanket, and temperature stimulator) induced positive in vitro and in vivo responses for tissue regeneration in skin (Harder et al. 2004), muscle (Riederer et al. 2008), cartilage (Hojo et al. 2003), teeth (Lee et al. 2008), heart (Wang et al. 2008; Rylander et al. 2005), and bone (Shui and Scutt 2001; Yoshida et al. 2009). Even though it is difficult to accurately compare the results of each study due to the variation in heating protocols employed and type of cells considered, utilization of low-level heating is preferred for skeletal tissue studies due to its ability to increase metabolic activity and cellular proliferation without cytotoxicity (Hojo et al. 2003; Shui and Scutt 2001; Ye et al. 2007). Heating with a water bath at 39–41°C for 1 h increased cell proliferation and ALP production (Shui and Scutt 2001) in bone-related cells and heating at 41°C or lower for 15–30 min improved proteoglycan metabolism with higher viability than sham-treated chondrocytes (Hojo et al. 2003). Furthermore, the use of conditioned culture media acquired from heat-treated osteoblasts (incubator heating at 42°C for 1 h) enhanced OCN secretion and mineralization in bone marrow mesenchymal cells (Ye et al. 2007). In addition to water bath or incubator heating, the exposure to elevated temperatures in the range of 42.5–44°C following treatment with 915 MHz microwave for 45 min generated new osseous tissue (Leon et al. 1993). Taken together, thermal stress conditioning can potentially serve as a positive cue for bone regeneration, which could be employed in combination with other tissue engineering exogenous stimuli such as GFs. However, the synergistic benefit of employing thermal stress and GFs for bone tissue engineering has not been explored.
The heat-stimulating effects causing bone formation may be mediated by elevated expression of heat shock proteins (HSPs). HSPs are induced by stresses that perturb the cellular environment, such as hypoxia (Mestril et al. 1994) and hyperthermia (Wang et al. 2008; Rylander et al. 2005; Wang et al. 2003). HSPs are multifunctional proteins involved in a variety of cellular activities such as anti-apoptotic self-protection in response to cytotoxic environments (Lee et al. 2009; Cooper et al. 2000), tissue regeneration (Hebb et al. 2006; Laplante et al. 1998), actin stabilization (Russotti et al. 1997), and differentiation by regulating caspase activity and stabilizing proteins associated with differentiation (Lanneau et al. 2007). In particular, HSP27 is involved in bone physiology through upregulation of TGF-β (Hatakeyama et al. 2002) and estrogen (Cooper and Uoshima 1994) which can increase bone mass, endothelin-1 (Tokuda et al. 2003), and prostaglandins (Kozawa et al. 2001). HSP47 is a procollagen-binding protein expressed in the endoplasmic reticulum (Nagata 1998) and involved in the biosynthesis of type I collagen (Dafforn et al. 2001), a major bone extracellular matrix protein. Treatment of MC3T3-E1 cells with TGF-β1 elevated HSP47 and type I collagen mRNA expression (Yamamura et al. 1998). Therefore, control of HSP27 and HSP47 may be important in modulating cellular events in skeletal tissue such as bone and cartilage (Tiffee et al. 2000). HSP70 has a potential role in enhancing anti-apoptotic effects following thermal stress (Riederer et al. 2008), but has exhibited variation in induction level depending on post-heating duration in response to elevated temperatures (44–50°C; Rylander et al. 2005). In bone-related cells, thermal stress caused HSP70 upregulation (Shui and Scutt 2001) and enhanced cell proliferation. Furthermore, HSP27 and HSP70 are involved in the apoptosis pathway and operate as anti-apoptotic molecules (Garrido et al. 2006) in response to diverse stresses (Terauchi et al. 2003; Dressel et al. 2000). Although HSPs have roles in bone physiology and can be induced by heat, limited understanding exists regarding the response of bone cells to thermal stress and the potential benefit of thermal stress conditioning for bone tissue engineering.
In vitro differentiation and the metabolic response of bone cells can be investigated by measuring induction of bone matrix proteins such as osteopontin (OPN; Klein-Nulend et al. 1997) and OCN (Shea et al. 2000), cytokines such as osteoprotegerin (OPG; Tang et al. 2006), and enzymes such as matrix metalloproteinases (MMPs; Mizutani et al. 2001). Matrix proteins such as OPN, OCN, bone sialoprotein (BSP), and type I collagen have been used as indicators to confirm successful osteogenic processes (Sharp et al. 2009; Klein-Nulend et al. 1997; Shea et al. 2000). VEGF is a key growth factor controlling bone formation as well as angiogenesis by mediating communication between endothelial cells and bone cells (Dai and Rabie 2007). Due to VEGF’s role as an angiogenic inducer, numerous tissue engineering experts have investigated enhancing VEGF levels in an osteo-scaffolding system with VEGF gene or protein delivery (Santos and Reis 2009). OPG has a critical role in bone physiology by diminishing osteoclast differentiation through mediation between the receptor activator nuclear factor κB (RANK) and RANK-ligand (Aubin and Bonnelye 2000). In addition, MMP-9 is used to identify the phenotype and differentiation status of osteoclast precursor cells into mature osteoclasts (Kaneshita et al. 2007) and is considered a therapeutic target to treat bone diseases such as osteoporosis (Rodan and Martin 2000).
To develop thermal stress conditioning protocols capable of enhancing bone formation, our study investigated whether in vitro thermal stress in combination with GFs could promote bone-related proteins which are potential indicators for osteogenic, anti-osteoclastic, and angiogenic activity as well as the stress-protective capacity of preosteoblasts. Therefore, we measured the induction of HSPs (HSP27, HSP47, and HSP70), an angiogenic factor (i.e., VEGF), and critical bone proteins (e.g., OPN, OCN, OPG, type I collagen, BSP, ALP, and MMP-9) by MC3T3-E1 cells following thermal conditioning utilizing a range of thermal stimulation protocols (44°C, 4 and 8 min) and two osteoinductive GFs (BMP-2 and TGF-β1) individually or in combination for varying post-heating (PH) timepoints (8, 24, and 72 h).
Materials and methods
Cell culture
A murine preosteoblastic cell line, MC3T3-E1 (ATCC), was cultured as a monolayer with growth media that was composed of alpha Minimum Essential Medium (αMEM; Mediatech) supplemented with 10% fetal bovine serum (FBS) and 1% penicillin–streptomycin (PS) in an incubator at 5% CO2 and 37°C. For cytotoxicity measurements, cells were seeded into 12 well plates at a concentration of 5x104/well and cultured for 24 h to allow adhesion before heating. For western blot and PCR analysis, cells were seeded into 25-cm2 T-flasks at 5×105/flask for measurement at 8–24 h PH and 3×105/flask for measurement at 72 h PH to avoid over-confluency of cells. Each group was cultured for 48 h prior to heating.
Water bath heating
A constant temperature water bath (ISOTEMP 210, Fisher Scientific) was employed as the mode of thermal conditioning to heat experimental specimens similar to methods described in our prior work (Rylander et al. 2005; Rylander et al. 2010). Media composed of MEM without l-glutamine was used during the heating process to prevent cell damage caused by the degradation of l-glutamine at high temperatures. Heating media was pre-warmed to the desired temperature and added to a monolayer of MC3T3-E1 cells. Subsequently, the flask was submerged in the water bath that was set at constant temperatures of 40–44°C for heating durations of 2–20 min. Following thermal stress, osteogenic media was added to promote osteogenesis during the PH period. Osteogenic media was composed of αMEM including 50 μg/ml l-ascorbic acid, 10 mM β-glycerol phosphate, 1% FBS, and 1% PS with or without the addition of BMP-2 (50 ng/ml) and TGF-β1 (2 ng/ml). The selected concentrations of GFs were previously shown by Chung et al. to cause differentiation of MC3T3-E1 cells (Chung et al. 1999). The osteogenic media for cytotoxicity measurements was supplemented with 10% FBS rather than 1% to permit ideal media conditions for cell proliferation. To permit induction of HSP and manifestation of cellular injury, cells were returned to a 5% CO2 incubator at 37°C after heating and post-incubated for varying durations depending on measurement type: cytotoxicity (24 h), western blot (24 h), PCR (8, 24, and 72 h), and enzyme-linked immunosorbent assay (ELISA; 72 h). We selected these short post-heating periods to investigate and compare the induction of HSPs and other bone-related proteins following stress treatment because previous studies described the transient induction of HSP occurring within several hours up to 72 h (Rylander et al. 2005; Wang et al. 2003).
Analysis of cytotoxicity
After MC3T3-E1 cells were heated at 40–44°C for 2–20 min and post-incubated for 24 h, cell viability was measured by 3-(4,5-dimethylthiazol-2-yl)-5-(3-carboxymethoxyphenyl)-2-(4-sulfophenyl)-2H-tetrazolium (MTS) assay using CellTiter96® Aqueous One Solution Cell Proliferation Assay (Promega). MTS solution mixed with basal αMEM media at a volume ratio of 1:5 was added to cultured cells. After 4-h incubation at 37°C, the solution was transferred to a 96-well plate and optical density was measured at 490 nm by a microplate reader (SpectraMax M2e, Molecular Devices).
Western blot
Western blot analysis was used to investigate the correlation between HSP expression and thermal stress conditioning (44°C for 0–8 min) at 24 h PH. Cells were lysed with RIPA buffer (Santa Cruz Biotechnology). For electrophoresis, isolated protein was loaded in a 10% Criterion Tris–HCl Gel (Bio-Rad Laboratories). Proteins transferred onto the membranes were immunoblotted using primary antibodies for HSP27, HSP47, and HSP70 and secondary antibodies diluted in 5% non-fat dry milk solution in Tween-20/Tris buffered saline (TBS) for blocking. All primary and secondary antibodies were purchased from Santa Cruz (SC) Biotechnology except for the HSP70 primary antibody (Stressgen). HSP27 (SC-51956, dilution ratio = 1:200), HSP47 (SC-13150, 1:500), and HSP70 (SPA-810, 1:1,000) were mouse monoclonal primary antibodies. HRP-conjugated secondary antibodies (SC-2005 for HSP27 and HSP47, and SC-2969 for HSP70, 1:2,000) corresponding with the specific species of each primary antibody were used. After adding a chemiluminescent solution, protein bands were visualized by LAS3000 Image Analyzer (Fujifilm). Quantification of each protein band on western blot images was accomplished by calculating the intensity difference between a band of interest and background per unit area using Multi Gauge V3.0 program (Fujifilm). After imaging, the membranes were stripped for actin normalization by incubating for 30 min at 50°C in the stripping solution.
Quantitative real time RT-PCR
Cells were heated at 44°C for 0, 4, and 8 min and gene expression for all markers shown in Table 1 was measured at 8, 24, and 72 h PH. RNA was isolated by spin protocol using RNeasy Mini kit (Qiagen) and QIAshredder (Qiagen). Isolation procedures were conducted according to manufacturer’s protocols. Isolated RNA was converted to cDNA using a Reverse Transcription System (Promega). After reverse transcription, cDNA samples were mixed with Taqman PCR Master Mix (Applied Biosystems) and each specific primer, and then polymerized in 7300 Real-Time PCR Systems (Applied Biosystems). The PCR reaction was initiated at 50°C for 2 min and 95°C for 10 min. For each polymerization (total PCR reaction cycles = 45), temperature was set at 95°C for 15 s and 60°C for 1 min. Taqman® Gene Expression Assays (Applied Biosystems) used for specific gene detection are listed as a primer and probe in Table 1. Relative fold induction (RFI) of each mRNA expression was calculated according to the 2-△△CT method used in Lee et al.’s study (Lee et al. 2004). Threshold cycle (C T), derived using SDS v1.2× system software of 7300 Real-Time PCR System, denotes the fractional cycle number at threshold polymerized gene and ∆∆C T was derived from (C T of target gene − C T of GAPDH)treated group − (C T of target gene − C T of GAPDH)control group (Lee et al. 2004). Treated groups refer to heated and GF-added only groups or groups treated with heat and GFs. For all tests, control groups indicated sham-treated cells with neither GF addition nor heating. Since all samples were normalized with the control groups and housekeeping GAPDH gene, the basal level of expression for the control groups was close to 1.
ELISA
Protein secretion was analyzed using ELISA. Conditioned media from cell monolayers was isolated at 72 h following thermal stress (44°C, 0–8 min). ELISA was conducted using the manufacturer’s protocol. OPG (MOP00), VEGF (MMV00), OPN (MOST00), and MMP-9 (MMPT90) were measured using Quantikine® ELISA (R&D Systems). OCN secretion data were acquired by Mouse Osteocalcin EIA kit (Biomedical Technologies). Culture supernatant was added to a 96-well microplate coated with specific antibodies for each secreted protein. The optical absorbance of the solution was measured at 450 nm by a microplate reader and compared to a standard curve. The concentration of secreted proteins in the culture supernatant was quantified from standard curve estimation.
Statistical analysis
Experimental groups with a minimum of three repetitions were utilized and analyzed independently. All graphs are presented as mean ± standard deviation (SD). One-way ANOVA with a Tukey multiple comparison test was used for analyzing the significant difference of cytotoxicity in heated samples compared to sham-treated control (p < 0.05). Two-way ANOVA with Tukey multiple comparison test was used to analyze experiments involving thermal stress conditioning in combination with GFs to demonstrate the statistical significant mean differences (p < 0.05).
Results
Cytotoxicity in response to thermal stress conditioning
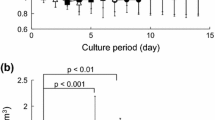
As a prerequisite for identifying feasible heating protocols for bone regeneration, we first characterized the threshold heating temperature and duration necessary to cause cytotoxicity. MTS data showed cytotoxicity after heating for durations longer than 20 min at 40ºC, 16 min at 42ºC, and 10 min at 44ºC (Fig. 1). Preosteoblast viability was not significantly altered compared to control groups provided a heating duration shorter than 10 min was chosen for all temperatures (40ºC, 42ºC, and 44ºC). In an effort to maximize response to heating, all future measurements were acquired using the highest water bath temperature of 44ºC with corresponding heating durations shorter than 10 min to maintain viability.

Cytotoxicity of MC3T3-E1 cells following of water bath heating for varying heating durations and temperatures. MTS assay was performed at 24 h PH at 40°C (a), 42°C (b), and 44°C (c). Absorbance at 490 nm was used as relative quantification data for cytotoxicity (mean ± SD). Asterisk represents statistical significant difference between control group (unheated) and heated groups (p < 0.05)
HSP expression in response to thermal stress conditioning and GF treatment
The effect of heating duration (0–8 min) at 44°C on protein levels of HSPs by MC3T3-E1 cells was measured using western blot analysis at 24 h PH (Fig. 2a, b). In addition, we investigated the impact of thermal stress (44°C, duration = 0, 4, 8 min) and GFs (BMP-2 and TGF-β1) independently and in combination on HSP gene expression by measuring mRNA levels of HSP27, HSP47, and HSP70 using RT-PCR (Fig. 2c–h) at 8, 24, and 72 h PH. Our results demonstrate that a single dose of thermal stress for less than 10 min can cause substantial induction in HSP expression. However, the expression of each protein was transient and the expression level varied for each HSP following thermal stress. Thermal stress increased gene expression of all HSPs for all protocols at 8 h PH (Fig. 2c, d; HSP27, 2.1 and 2.4 RFI; HSP47, 1.8 and 4.4 RFI; HSP70, 10.2 and 63 RFI following 4 and 8 min of heating, respectively). Statistically significant induction in HSP47 and HSP70 mRNA occurred following 8 min of heating. HSP70 exhibited the greatest increase in mRNA (63 RFI) following thermal stress compared to HSP27 (2.4 RFI) and HSP47 (4.4 RFI) at 8 h PH. Similarly, western blot analysis demonstrated HSP70 increased dramatically compared to the other HSPs at 24 h PH. The protein expression of HSP27 was observed only in the groups exposed to 6 and 8 min of heating whereas non-heated groups or groups exposed to heating durations shorter than 6 min exhibited no visible HSP27 protein bands. HSP47 exhibited a high level of basal expression which was unaffected by thermal stress in spite of the induction of HSP47 mRNA.

Expression of HSP27, HSP47, and HSP70 following heating (44°C for 0–8 min) alone or in combination with GFs (BMP-2 and TGF-β1) for varying PH duration (8, 24, and 72 h). Western blot analysis of HSP expression following heating alone with varying heating duration (0, 2, 4, 6, and 8 min; PH = 24 h; a) and corresponding western blot quantification of HSP expression (n = 3; b). Gene expression following thermal conditioning (44°C, 0, 4, and 8 min) and post-culturing with GFs (BMP-2 and TGF-β1) for 8, 24, and 72 h (c–h). HSP27, HSP47, and HSP70 mRNA expression at PH durations of 8 (c, d), 24 (e, f), and 72 h (g, h). Asterisk, double asterisks, and number sign represent statistical significance between control (non-heated without GFs) and stressed groups, between groups heated for 4 and 8 min, and between each heated group in the absence or presence of GFs (e.g., 4 min heating with or without GFs), respectively (p < 0.05). N.D. denotes no detection in western blot analysis
Thermal stress independently and in combination with GFs increased gene expression of all HSPs for all protocols at 8 h PH, while the addition of GFs following 8 min of heating diminished HSP expression (no statistical significance); HSP27 (2.4 RFI without GFs; 1.6 RFI with GFs), HSP47 (4.4 RFI without GFs; 3.0 RFI with GFs), and HSP70 (63.0 RFI without GFs; 35.6 RFI with GFs). At 24 h PH (Fig. 2e, f), the fold inductions observed following treatment with heating and GFs decreased compared to those observed at 8 h PH. HSP70 gene expression increased 5.0- and 24.1-fold following 4 and 8 min of heating without GFs, respectively. However, the addition of GFs diminished the benefit achieved by heating with corresponding HSP70 RFI values of 0.7, 1.9, and 5.5 following 0, 4, and 8 min of heating with GFs, respectively. HSP27 did not show any significant change in expression following both individual and combinatorial heating and GF addition. However, HSP47 mRNA was induced more significantly with GF addition with 7.5 RFI in non-heated groups and 2.3 and 1.1 RFI following 4 and 8 min of heating with GFs, respectively. Heating in combination with GFs diminished the benefit of GFs. At 72 h PH (Fig. 2g, h), GFs significantly suppressed HSP70 expression (lower than 0.4 RFI) and exhibited similar HSP27 and HSP47 mRNA expression levels compared to those without GF addition.
Gene expression and protein release of bone-related proteins in response to thermal stress conditioning and GF treatment
The response of preosteoblasts to combined thermal stress conditioning and GFs was determined by measuring several bone-related proteins: matrix proteins (OCN, OPN, BSP, and type I collagen), GFs (VEGF), cyclooxygenase-2 (COX-2), the cytokine OPG, and enzymes (MMP-9 and ALP). Figure 3 shows that numerous bone-related genes exhibited increased induction in response to heating alone and in combination with GFs (TGF-β1 and BMP-2) for longer heating durations at 8 h PH: OPN (without GFs: 1.1 and 1.8 RFI; with GFs: 0.5 and 0.8 RFI after 4 and 8 min of heating, respectively), OCN (without GFs: 1.9 and 3.8 RFI; with GFs: 1.3 and 2.0 RFI after 4 and 8 min of heating, respectively), and OPG (without GFs: 1.2 and 2.1 RFI; with GFs: 5.6 and 6.9 RFI after 4 and 8 min of heating, respectively). The addition of GFs caused a significant decrease in gene levels of OCN and OPN, however, a significantly greater induction of OPG compared to heating alone (8 min) was observed. Specifically, OPN expression showed a significant increase of 1.8 RFI following 8 min of heating without GFs compared to control. However, GF inclusion following 4 and 8 min of heating significantly suppressed OPN with 0.5 and 0.8 RFI, respectively, compared to cells experiencing heating only. OCN expression increased 1.9 and 3.8 RFI following 4 and 8 min of heating without GFs, respectively (only 8 min, p < 0.05). However, the addition of GFs to samples heated for 4 and 8 min suppressed gene expression with 1.3 and 2.0 RFI (only 8 min, p < 0.05). OPG expression significantly increased 5.6 and 6.9 RFI for 4 and 8 min of heating, respectively in combination with GFs as compared to 1.2 and 2.1 RFI for heating alone for 4 and 8 min, respectively.

Expression of bone-related mRNA following thermal conditioning (44°C, 0, 4, and 8 min) and post-culturing with GFs (BMP-2 and TGF-β1) for PH durations of 8, 24, and 72 h. OPN, OCN, OPG, and MMP-9 mRNA at 8 h PH (a); ALP, BSP, and type I collagen (COL 1) mRNA at 8 (b), 24 (d), and 72 h PH (f); OPN, OCN, OPG, and MMP-9 mRNA at 24 (c) and 72 h PH (e). Asterisk, double asterisks, and number sign represent statistical significance between control (non-heated without GFs) and stressed groups, between groups heated for 4 and 8 min, and between each heated group in the absence or presence of GFs (e.g., 4 min heating with or without GFs), respectively (p < 0.05)
At 24 h PH, OPN and OPG showed similar induction trends in response to heating or GF addition compared to 8 h PH data. However, OCN did not show any significant response to heating or GFs. At 72 h PH, OPN did not show any apparent induction in response to heating and GFs. OPG showed significant increase (1.5 RFI) only in the group experiencing 8 min of heating with GFs compared to the control. MMP-9 significantly increased at 8 h following GF addition for both heated (2.8 and 2.1 RFI for 4 and 8 min of heating) and sham-treated cells (2.8 RFI). However, without GF addition, MMP-9 decreased significantly following 8 min of heating for PH durations of both 24 (0.4 RFI) and 72 h (0.2 RFI). GF treatment alone and in combination with heating inhibited MMP-9 (0.1 RFI) expression more significantly compared to heating alone (0.4 and 0.2 RFI for 4 and 8 min of heating) and there was no significant change in MMP-9 expression following heating with GFs after 72 h.
The thermal conditioning protocols utilized in this study hindered the induction of ALP and BSP genes (Fig. 3). ALP decreased following heating alone and in combination with GFs for all PH durations and significantly decreased after 8 min of heating in combination with GFs compared to groups treated with only GFs following periods of 8 and 24 h: 1.1 RFI after 8 min of heating with GFs and 1.6 RFI in only GF groups following 8 h; 0.5 RFI after 8 min of heating with GFs and 2.0 RFI for treatment with GFs at 24 h. Interestingly, BSP increased significantly in response to GF addition alone (4.6 RFI) at 72 h but heating significantly diminished its induction (∼ 1 RFI).
ELISA was performed to investigate the secretion of bone proteins from MC3T3-E1 cells into the culture media following heating (44°C, 0, 4, and 8 min) and GF addition individually and in combination at 72 h (Fig. 4). Without heating, GF addition alone significantly promoted secretion of OPN (5.4-fold) and OPG (10.7-fold) while inhibiting MMP-9 expression so significantly that it was not detectable by ELISA. In response to heating alone, OPN and OPG showed a slight increase (not statistically significant) in OPN and OPG levels which was much lower than those of GF-treated groups. OPG and OPN showed significantly decreased secretion in response to heating (4 min) and GFs. Moreover, OCN secretion in cells without GFs was too low to be detected by ELISA. Sham-treated and groups heated for 4 and 8 min of heating without GFs decreased in MMP-9 release with longer heating duration corresponding to concentrations of 621.2, 545.4, and 441.3 pg/ml, respectively.

Secretion of bone-related proteins following thermal conditioning (44°C, 0, 4, and 8 min) and post-culturing with GFs (BMP-2 and TGF-β1) for 72 h. The concentrations of secreted OPN (a), OCN (b), OPG (c), and MMP-9 (d) were quantified using ELISA. Asterisk, double asterisks, and number sign represent statistical significance between control (non-heated without GFs) and stressed groups, between groups heated for 4 and 8 min, and between each heated group in the absence or presence of GFs (e.g., 4 min heating with or without GFs), respectively (p < 0.05)
Our study revealed that COX-2 mRNA was significantly upregulated by GF addition for all PH durations (Fig. 5). Furthermore, COX-2 expression increased with longer heating duration in response to thermal stress: 2.7 and 9.8 RFI following 4 and 8 min of heating, respectively at 8 h PH. However, for 8 min of heating and increasing PH duration, the induction effect of heating on COX-2 RFI diminished with increasing PH heating duration (9.8, 3.3, and 1.6 RFI at PH durations of 8, 24, and 72 h, respectively).

COX-2 mRNA expression following thermal conditioning (44°C, 0, 4, and 8 min) and post-culturing with GFs (BMP-2 and TGF-β1) for 8, 24, and 72 h. Asterisk, double asterisks, and number sign represent statistical significance between control (non-heated without GFs) and stressed groups, between groups heated for 4 and 8 min, and between each heated group in the absence or presence of GFs (e.g., 4 min heating with or without GFs), respectively (p < 0.05)
Induction of VEGF in response to thermal stress conditioning and GF treatment
As shown in Fig. 6, thermal stress (8 min) at 44°C in combination with GFs caused significantly greater expression of the VEGF gene (at 8 and 24 h PH) and protein (72 h PH) compared to heating or GF addition independently, implying these methods may be conducive to enhancing angiogenesis in bone healing. GF addition induced VEGF mRNA more significantly than heating for all PH durations. Eight minutes of heating with GFs caused the greatest VEGF mRNA induction at 8 h PH (16.0 RFI) whereas heating alone induced only a 5.3 RFI in VEGF mRNA expression. At 24 h PH, VEGF exhibited similar trends at 8 h PH, but the RFI level of VEGF declined for samples heated alone (1.5 RFI after 8 min of heating) and in combination with GFs (10.7 RFI). At 72 h PH, heated samples returned to the basal level, but samples treated with GFs still maintained significantly elevated expression (∼5 RFI) compared to the control. Without GF addition, secreted VEGF into the culture medium was significantly decreased to a nondetectable level. Heating for durations of 4 and 8 min with GFs significantly enhanced the release of VEGF concentration by 9.0- and 6.2-fold, respectively, compared to non-heated groups with GFs.

VEGF gene and protein expression following thermal conditioning (44°C, 0, 4, and 8 min) and post-culturing with GFs (BMP-2 and TGF-β1). VEGF mRNA expression (a) at different PH durations (8, 24, and 72 h). Released VEGF in the culture media during 72 h PH with GF addition analyzed by ELISA (b). Asterisk, double asterisks, and number sign represent statistical significance between control (non-heated without GFs) and stressed groups, between groups heated for 4 and 8 min, and between each heated group in the absence or presence of GFs (e.g., 4 min alone and 4 min with GFs), respectively (p < 0.05)
Discussion
In this study, we demonstrated a single application of thermal stress conditioning can cause rapid induction of heat shock proteins and expression of bone-related proteins by MC3T3-E1 preosteoblastic cells. Furthermore, a novel combinatorial stress conditioning approach utilizing heating and growth factors (i.e., TGF and BMP-2) was also explored to determine whether this combination has a synergistic potential for bone healing.
We concluded that heating temperature of 40–44°C with durations shorter than 10 min was conducive to sustained viability (Fig. 1). These results are comparable to studies by Rylander et al. in which cancerous (PC3) and normal (RWPE-1) prostate cells were heated with a water bath at 44°C to 60°C for 1 to 30 min (Rylander et al. 2010). Heating protocol parameters such as heating duration and temperature can affect the thermal stress experienced by the cells. The thermal response time for our water bath heating protocol is much shorter compared to incubator-based heating (Lee et al. 2008; Ye et al. 2007; Russotti et al. 1996), causing the onset of injury and protein induction to occur earlier. Prior studies using water bath heating have utilized lower temperatures (39–41°C) with longer heating durations (15 min–1 h), demonstrating beneficial cellular effects (Hojo et al. 2003; Shui and Scutt 2001). In order to maximize HSP gene and protein induction, which occurs more significantly at higher temperatures (Rylander et al. 2005) while maintaining high cell viability, we utilized the highest temperature (44°C) for heating durations of 0–8 min for measurements of thermal induction of HSPs and bone-related proteins.
The measurement of HSPs (Fig. 2) was a critical aspect of this study due to their importance in bone physiology (Tiffee et al. 2000), matrix protein production (Dafforn et al. 2001), and their thermally induced expression kinetics (Wang et al. 2008). According to an immunostaining study using rat tibia, HSPs are believed to have important roles in the bone formation process since HSP27, HSP47, and HSP70 are highly expressed in osteoblasts of new bone-generating areas with type I collagen (Tiffee et al. 2000; Rylander et al. 2010). To our knowledge, this is the first study to demonstrate induction of HSP27, HSP47, and HSP70 by preosteoblasts following identical heating conditions in vitro. Prior studies using cyclic tension (6 h, 15% magnitude, 1 cycle/s) showed only an 8.1-fold induction in HSP70 and a decrease in HSP27 mRNA in trabecular meshwork cells (Luna et al. 2009). Therefore, thermal stress may provide a more powerful conditioning method for inducing HSPs requiring only a single short thermal dose rather than repeated conditioning strategies, such as stretching (Luna et al. 2009) or hydrostatic pressure (Kaarniranta et al. 2003).
Comparable HSP70 expression levels and trends were observed in studies by Rylander et al. in which endothelial cells were heated with a water bath (44–46°C for 0–10 min) and HSP expression was measured at 16–18 h PH (Rylander et al. 2005). Similarly, Wang et al. reported heating using an incubator set at 42°C for 1.6 h caused transient induction of HSP70 with maximum expression for a PH of 16 h (Wang et al. 2008). Other studies have also shown increased HSP70 levels (∼1.7-fold) compared to control by chondrocytes heated at 43°C for 1 h with an incubator (Sawatzky et al. 2005). Thermally induced HSP27 and HSP70 expression kinetics observed in this study are similar to prior studies by Rylander et al. in which the temporal HSP expression profiles in cancerous (PC3) and normal (RWPE-1) prostate cells were measured following thermal stress (Rylander et al. 2010). In particular, HSP27 upregulation in MC3T3-E1 cells may offer protection to external stresses while also modulating several important molecules relevant in bone physiology. MC3T3-E1 cells have been previously shown to regulate HSP27-related transcripts by estrogen pretreatment in response to thermal stress (Cooper and Uoshima 1994) and promote HSP27 by prostaglandin D2, whose action is mediated by protein kinase C and mitogen-activated protein kinase (MAPK) signaling pathway (e.g., p44/42 and p38; Kozawa et al. 2001).
Although the effect of GFs on HSP induction is poorly understood and can vary depending on target cells and treatment protocols, previous literature has shown induction of HSPs by TGF-β (Yamamura et al. 1998; Cao et al. 1999; Hayashi et al. 2008). Our study suggests that dual treatment with osteoinductive GFs in the form of TGF-β1 and BMP-2 induced HSP47, but did not contribute to induction of HSP27 and HSP70. The variation in response of each HSP to GFs may be caused by differences in their inherent function in cells. For example, in collagen-secreting cells, increased expression of HSP47 mRNA caused by TGF-β1 (Yamamura et al. 1998) can contribute to regulation of collagen synthesis by serving as a procollagen-binding molecular chaperone (Dafforn et al. 2001). However, HSP27 and HSP70 are related to intracellular apoptosis-related mechanisms (Lanneau et al. 2007).
In this study, gene expression of each bone-related protein in MC3T3-E1 cells, which were measured at 8, 24, and 72 h following thermal conditioning at 44°C for 0–8 min using identical methods as described for HSPs, showed varying induction trends depending on time points (Figs. 3, 5, and 6). This broad screening of bone-related proteins is critical to determine whether thermal stress conditioning and GFs individually or in combination may potentially cause beneficial or deleterious effects associated with osteogenesis, angiogenesis, and osteoclastogenesis in bone. The induction or suppression of the aforementioned bone proteins regulates bone physiological processes. The interaction among bone cells (Matsuo 2009) and between osteoblasts and endothelial cells (Grellier et al. 2009) is controlled by secretion of bioactive molecules such as BMP-2 and VEGF or mediation through ligands such as the receptor activator of NF-κB ligand (RANKL) which is related to OPG.
The upregulation of OCN mRNA by thermal stress preconditioning, as shown in our study, could potentially promote bone formation. Similarly, our study demonstrated the capability of thermal stress preconditioning to upregulate OPN mRNA. Previously, elevated levels of OCN mRNA have been accompanied with high mineralization in native bone-like MC3T3-E1-seeded scaffolds exhibiting specialized cell localization (Shea et al. 2000). Therefore, induction of OCN mRNA by heating, as shown in our study, could potentially accelerate in vitro osteogenesis. Similarly, prior literature (Klein-Nulend et al. 1997) has shown OPN gene induction by mechanical stress contributes to bone regeneration, thereby offering another benefit to our approach involving thermally-induced OPN mRNA. In addition, COX-2, which operates as an enzyme critical for prostaglandin production and is necessary for bone repair (Einhorn 2003; O’Keefe et al. 2006), was revealed here to be transiently induced more significantly with longer thermal stress duration and by GF addition (Fig. 5).
Our study demonstrates greater induction of OPG genes following conditioning with heating and GFs in combination compared to individual treatment with heating and GFs. In addition, we showed that either GF treatment or heating can downregulate MMP-9. Inhibition of bone resorptive enzymes, such as MMP-9, and promotion of anti-osteoclastogenic proteins such as OPG within the bone microenvironment could be utilized to mitigate bone over-resorption, which causes bone diseases such as osteoporosis (Rodan and Martin 2000). Although osteoinductive GF treatment (e.g., BMP-2 and TGF-β1) is known to promote OPG induction (Sato et al. 2009) and OPG/OCIF (osteoclastogenesis inhibitory factor) (Takai et al. 1998) in MC3T3-E1 cells, this is the first study investigating the effect of thermal stress alone and in combination with GFs on OPG induction. In addition to OPG’s inhibitory role in osteoclastogenesis, Grundt et al. suggested it can upregulate the level of ALP directly in a dose-dependent manner (Grundt et al. 2009). Taken together, our results suggest that conditioning with heating and GFs in combination could be applied to inhibit osteoclast differentiation and bone resorptive processes in the bone microenvironment.
The thermal preconditioning protocols utilized in this study hindered the induction of ALP and BSP genes (Fig. 3). Since mild heating at 42°C or lower for 30–60 min has been reported in studies to promote levels of ALP for PH periods of 4 days or longer than 1 week, it is difficult to compare prior results to our study due to the differences in heating methodology (Lee et al. 2008; Shui and Scutt 2001). Furthermore, type I collagen mRNA exhibited no significant difference following thermal conditioning and GFs for all three PH periods. These results, which may be a negative sign for bone healing, should be investigated further to develop more optimal protocols for inducing these proteins by using more mild-heating preconditioning or long-term analysis of bone-related proteins. In general, since we utilized only a single dose of thermal stress followed by analysis at a limited number of timepoints during short-term PH, the enhancement of bone-related proteins and HSPs may be transient and fail to induce sustained upregulation and appropriate timepoints for analysis may not be considered. Long-term stress conditioning using a 3D scaffold in a thermal conditioning bioreactor could provide bone cells sufficient time for in vitro bone formation and mineralization.
Our results (Fig. 6) showed the combination of thermal stress preconditioning and GFs could synergistically upregulate VEGF implying these methods may be conducive to enhancing angiogenesis in bone healing. Without GFs, thermal stress induced rapid upregulation of VEGF after short PH periods of several hours. Production of VEGF is critical to stimulation of blood vessel formation necessary for bone regeneration or neovascularization within a tissue-engineered scaffold. Although mechanical stress (Singh et al. 2007; Hurley et al. 2010) and biochemical cues (Deckers et al. 2002) have been extensively shown to enhance angiogenesis, the impact of thermal stress on VEGF regulation has not been sufficiently explored. According to Gong et al.’s study (Gong et al. 2006) using a rat animal model, cardiac tissue expressed more VEGF with upregulation of HSP70 when it was exposed to elevated temperatures (42°C for 15 min) generated by a heating pad.
Conclusion
This study revealed that preosteoblasts exposed to elevated temperature (44°C) and two exogenous osteoinductive GFs (BMP-2 and TGF-β1) individually or in combination showed potential angiogenic and anti-osteoclastic effects with varied alterations in expression of HSPs and bone-related proteins (e.g., VEGF). Heating at 44°C (for 8 min) rapidly induced OCN, OPN, OPG, VEGF, and COX-2 mRNA at 8 h post-heating. The addition of GFs with heating, induced OPG and VEGF genes more significantly compared to heating or GF addition alone. OPN, OCN, and OPG secretions increased with the addition of GFs. However, MMP-9 secretion was inhibited by heating, with more significant declines associated with GF inclusion. Further research should focus on optimizing the thermal stress conditioning parameters (e.g. dose and post-heating durations between repeated thermal stress events) using a more controlled and automated heating system. Based on this study, individual or combined conditioning with heating and exogenous GFs can potentially serve as a new strategy to inhibit the imbalance between osteoclasts and osteoblasts associated with bone diseases such as osteoporosis and induce bone formation and angiogenesis critical for bone regeneration.
References
Teitelbaum SL (2000) Osteoclasts, integrins, and osteoporosis. J Bone Miner Metab 18(6):344–349
Singh SP et al (2007) Cyclic mechanical strain increases production of regulators of bone healing in cultured murine osteoblasts. J Am Coll Surg 204(3):426–434
Sharp LA, Lee YW, Goldstein AS (2009) Effect of low-frequency pulsatile flow on expression of osteoblastic genes by bone marrow stromal cells. Ann Biomed Eng 37(3):445–453
Choi JY et al (1996) Expression patterns of bone-related proteins during osteoblastic differentiation in MC3T3-E1 cells. J Cell Biochem 61(4):609–618
Kandziora F et al (2002) IGF-I and TGF-beta1 application by a poly-(D,L-lactide)-coated cage promotes intervertebral bone matrix formation in the sheep cervical spine. Spine (Phila Pa 1976) 27(16):1710–1723
Wei G et al (2007) The enhancement of osteogenesis by nano-fibrous scaffolds incorporating rhBMP-7 nanospheres. Biomaterials 28(12):2087–2096
Jeong JC et al (2004) Drynariae Rhizoma promotes osteoblast differentiation and mineralization in MC3T3-E1 cells through regulation of bone morphogenetic protein-2, alkaline phosphatase, type I collagen and collagenase-1. Toxicol In Vitro 18(6):829–834
Zhang F et al (2009) Sustained BMP signaling in osteoblasts stimulates bone formation by promoting angiogenesis and osteoblast differentiation. J Bone Miner Res 24(7):1224–1233
Ryoo HM, Lee MH, Kim YJ (2006) Critical molecular switches involved in BMP-2-induced osteogenic differentiation of mesenchymal cells. Gene 366(1):51–57
Spinella-Jaegle S et al (2001) Opposite effects of bone morphogenetic protein-2 and transforming growth factor-beta1 on osteoblast differentiation. Bone 29(4):323–330
Peterson WJ, Tachiki KH, Yamaguchi DT (2004) Serial passage of MC3T3-E1 cells down-regulates proliferation during osteogenesis in vitro. Cell Prolif 37(5):325–336
Carano RA, Filvaroff EH (2003) Angiogenesis and bone repair. Drug Discov Today 8(21):980–989
Dai J, Rabie AB (2007) VEGF: an essential mediator of both angiogenesis and endochondral ossification. J Dent Res 86(10):937–950
Street J et al (2002) Vascular endothelial growth factor stimulates bone repair by promoting angiogenesis and bone turnover. Proc Natl Acad Sci U S A 99(15):9656–9661
Harder Y et al (2004) Improved skin flap survival after local heat preconditioning in pigs. J Surg Res 119(1):100–105
Riederer I et al (2008) Heat shock treatment increases engraftment of transplanted human myoblasts into immunodeficient mice. Transplant Proc 40(2):624–630
Hojo T et al (2003) Effect of heat stimulation on viability and proteoglycan metabolism of cultured chondrocytes: preliminary report. J Orthop Sci 8(3):396–399
Lee MW et al (2008) Heat stress induces alkaline phosphatase activity and heat shock protein 25 expression in cultured pulp cells. Int Endod J 41(2):158–162
Wang S et al (2008) HSP70 kinetics study by continuous observation of HSP-GFP fusion protein expression on a perfusion heating stage. Biotechnol Bioeng 99(1):146–154
Rylander MN et al (2005) Correlation of HSP70 expression and cell viability following thermal stimulation of bovine aortic endothelial cells. J Biomech Eng 127(5):751–757
Shui C, Scutt A (2001) Mild heat shock induces proliferation, alkaline phosphatase activity, and mineralization in human bone marrow stromal cells and Mg-63 cells in vitro. J Bone Miner Res 16(4):731–741
Yoshida K et al (2009) Influence of heat stress to matrix on bone formation. Clin Oral Implants Res 20:782–790
Ye CP et al (2007) Culture media conditioned by heat-shocked osteoblasts enhances the osteogenesis of bone marrow-derived mesenchymal stromal cells. Cell Biochem Funct 25(3):267–276
Leon SA et al (1993) Effects of hyperthermia on bone. II. Heating of bone in vivo and stimulation of bone growth. Int J Hyperthermia 9(1):77–87
Mestril R et al (1994) Isolation of a novel inducible rat heat-shock protein (HSP70) gene and its expression during ischaemia/hypoxia and heat shock. Biochem J 298(Pt 3):561–569
Wang S, Diller KR, Aggarwal SJ (2003) Kinetics study of endogenous heat shock protein 70 expression. J Biomech Eng 125(6):794–797
Lee J et al (2009) Controlled delivery of heat shock protein using an injectable microsphere/hydrogel combination system for the treatment of myocardial infarction. J Control Release 137(3):196–202
Cooper LF et al (2000) Estrogen-induced resistance to osteoblast apoptosis is associated with increased hsp27 expression. J Cell Physiol 185(3):401–407
Hebb MO, Myers TL, Clarke DB (2006) Enhanced expression of heat shock protein 27 is correlated with axonal regeneration in mature retinal ganglion cells. Brain Res 1073–1074:146–150
Laplante AF et al (1998) Expression of heat shock proteins in mouse skin during wound healing. J Histochem Cytochem 46(11):1291–1301
Russotti G et al (1997) Studies of heat and PGA(1)-induced cold tolerance show that HSP27 may help preserve actin morphology during hypothermia. Tissue Engineering 3(2):135–147
Lanneau D et al (2007) Apoptosis versus cell differentiation: role of heat shock proteins HSP90, HSP70 and HSP27. Prion 1(1):53–60
Hatakeyama D et al (2002) Upregulation by retinoic acid of transforming growth factor-beta-stimulated heat shock protein 27 induction in osteoblasts: involvement of mitogen-activated protein kinases. Biochim Biophys Acta 1589(1):15–30
Cooper LF, Uoshima K (1994) Differential estrogenic regulation of small M(r) heat shock protein expression in osteoblasts. J Biol Chem 269(11):7869–7873
Tokuda H et al (2003) Involvement of stress-activated protein kinase/c-Jun N-terminal kinase in endothelin-1-induced heat shock protein 27 in osteoblasts. Eur J Endocrinol 149(3):239–245
Kozawa O et al (2001) Mechanism of prostaglandin D(2)-stimulated heat shock protein 27 induction in osteoblasts. Cell Signal 13(8):535–541
Nagata K (1998) Expression and function of heat shock protein 47: a collagen-specific molecular chaperone in the endoplasmic reticulum. Matrix Biol 16(7):379–386
Dafforn TR, Della M, Miller AD (2001) The molecular interactions of heat shock protein 47 (Hsp47) and their implications for collagen biosynthesis. J Biol Chem 276(52):49310–49319
Yamamura I et al (1998) Transcriptional activation of the mouse HSP47 gene in mouse osteoblast MC3T3-E1 cells by TGF-beta 1. Biochem Biophys Res Commun 244(1):68–74
Tiffee JC, Griffin JP, Cooper LF (2000) Immunolocalization of stress proteins and extracellular matrix proteins in the rat tibia. Tissue Cell 32(2):141–147
Garrido C et al (2006) Heat shock proteins 27 and 70: anti-apoptotic proteins with tumorigenic properties. Cell Cycle 5(22):2592–2601
Terauchi R et al (2003) Hsp70 prevents nitric oxide-induced apoptosis in articular chondrocytes. Arthritis Rheum 48(6):1562–1568
Dressel R et al (2000) Heat shock protein 70 is able to prevent heat shock-induced resistance of target cells to CTL. J Immunol 164(5):2362–2371
Klein-Nulend J et al (1997) Mechanical stimulation of osteopontin mRNA expression and synthesis in bone cell cultures. J Cell Physiol 170(2):174–181
Shea LD et al (2000) Engineered bone development from a pre-osteoblast cell line on three-dimensional scaffolds. Tissue Eng 6(6):605–617
Tang L, Lin Z, Li YM (2006) Effects of different magnitudes of mechanical strain on osteoblasts in vitro. Biochem Biophys Res Commun 344(1):122–128
Mizutani A et al (2001) Expression of matrix metalloproteinases during ascorbate-induced differentiation of osteoblastic MC3T3-E1 cells. J Bone Miner Res 16(11):2043–2049
Santos MI, Reis RL (2009) Vascularization in bone tissue engineering: physiology, current strategies, major hurdles and future challenges. Macromol Biosci 10(1):12–27
Aubin JE, Bonnelye E (2000) Osteoprotegerin and its ligand: a new paradigm for regulation of osteoclastogenesis and bone resorption. Osteoporos Int 11(11):905–913
Kaneshita Y, Goda S, Kawamoto T (2007) The effect of matrix metalloproteinase-9 on the differentiation into osteoclast cells on RAW264 cells. Orthodontic Waves 66(4):122–128
Rodan GA, Martin TJ (2000) Therapeutic approaches to bone diseases. Science 289(5484):1508–1514
Rylander MN et al (2010) Measurement and mathematical modeling of thermally induced injury and heat shock protein expression kinetics in normal and cancerous prostate cells. Int J Hyperthermia 26(8):748–764
Chung CY et al (1999) Serial passage of MC3T3-E1 cells alters osteoblastic function and responsiveness to transforming growth factor-beta1 and bone morphogenetic protein-2. Biochem Biophys Res Commun 265(1):246–251
Lee YW et al (2004) Gene expression profile in interleukin-4-stimulated human vascular endothelial cells. Mol Med 10(1–6):19–27
Russotti G et al (1996) Induction of tolerance to hypothermia by previous heat shock using human fibroblasts in culture. Cryobiology 33(5):567–580
Luna C et al (2009) Alterations in gene expression induced by cyclic mechanical stress in trabecular meshwork cells. Mol Vis 15:534–544
Kaarniranta K et al (2003) Stress responses of mammalian cells to high hydrostatic pressure. Biorheology 40(1–3):87–92
Sawatzky DA et al (2005) Heat-shock proteins and their role in chondrocyte protection, an application for autologous transplantation. Inflammopharmacology 12(5–6):569–589
Cao Y et al (1999) TGF-beta1 mediates 70-kDa heat shock protein induction due to ultraviolet irradiation in human skin fibroblasts. Pflugers Arch 438(3):239–244
Hayashi K et al (2008) (−)-Epigallocatechin gallate reduces transforming growth factor beta-stimulated HSP27 induction through the suppression of stress-activated protein kinase/c-Jun N-terminal kinase in osteoblasts. Life Sci 82(19–20):1012–1017
Matsuo K (2009) Cross-talk among bone cells. Curr Opin Nephrol Hypertens 18(4):292–297
Grellier M, Bordenave L, Amedee J (2009) Cell-to-cell communication between osteogenic and endothelial lineages: implications for tissue engineering. Trends Biotechnol 27(10):562–571
Einhorn TA (2003) Cox-2: where are we in 2003?—The role of cyclooxygenase-2 in bone repair. Arthritis Res Ther 5(1):5–7
O’Keefe RJ et al (2006) COX-2 has a critical role during incorporation of structural bone allografts. Skeletal Development and Remodeling in Health, Disease, and Aging 1068:532–542
Sato MM et al (2009) Bone morphogenetic protein-2 enhances Wnt/beta-catenin signaling-induced osteoprotegerin expression. Genes Cells 14(2):141–153
Takai H et al (1998) Transforming growth factor-beta stimulates the production of osteoprotegerin/osteoclastogenesis inhibitory factor by bone marrow stromal cells. J Biol Chem 273(42):27091–27096
Grundt A et al (2009) Direct effects of osteoprotegerin on human bone cell metabolism. Biochem Biophys Res Commun 389:550–555
Hurley NE et al (2010) Modulating the functional contributions of c-Myc to the human endothelial cell cyclic strain response. J Vasc Res 47(1):80–90
Deckers MM et al (2002) Bone morphogenetic proteins stimulate angiogenesis through osteoblast-derived vascular endothelial growth factor A. Endocrinology 143(4):1545–1553
Gong B et al (2006) Whole-body hyperthermia induces up-regulation of vascular endothelial growth factor accompanied by neovascularization in cardiac tissue. Life Sci 79(19):1781–1788
Funding sources
The following are the funding sources of this work: National Science Foundation grant CBET 0966546 and Institute for Critical Technologies and Applied Sciences (ICTAS) at Virginia Tech.
Conflict of interest
All authors have no conflicts of interest.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article
Cite this article
Chung, E., Rylander, M.N. Response of preosteoblasts to thermal stress conditioning and osteoinductive growth factors. Cell Stress and Chaperones 17, 203–214 (2012). https://doi.org/10.1007/s12192-011-0300-8
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s12192-011-0300-8