
Abstract
The aim of this paper is to provide a unified Lyapunov functional for an age-structured model describing a virus infection. Our main contribution is to consider a very general nonlinear infection function, gathering almost all usual ones, for the following problem:
where T(t), i(t, a) and V(t) are the populations of uninfected cells, infected cells with infection age a and free virus at time t respectively. The functions \(\delta (a),\) p(a), are respectively, the age-dependent per capita death, and the viral production rate of infected cells with age a. The global asymptotic analysis is established, among other results, by the use of compact attractor and strongly uniform persistence. Finally some numerical simulations illustrating our results are presented.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
1 Introduction
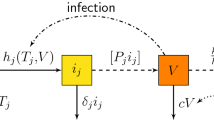
There is a huge number of works in the existing literature; proposing and studying virus infection mathematical models; we invite the reader to see for example [1, 3, 5, 7, 8, 14, 15, 23, 25, 26, 33] and the references therein. In the particular case of an HIV infection, the corresponding model is usually divided into three classes; namely uninfected cells, T, infected cells, i and free virus particles, V.
In order to establish a more realistic HIV model, many authors have considered the age state in their models, as for example in [6, 16, 24, 27, 34]. This was done with the aim to make distinction between cells, so the infection age that is time since cell is infected, is introduced in the subpopulation of infected cells, which makes, among other considerations, a variation of the mortality in this class.
Another important aspect to be considered, is the infection function, which represents the interaction between the virus and the uninfected cells. Frequently, this interaction is described by a bilinear function (mass action) or some standard functions. However these cited functions are not always a good representation of this complicated phenomenon [11, 12, 32].
It is established that the process and the evolution of these infections, are governed by partial differential equations, and thus it is not obvious to study the global properties of their solutions, see [4, 20,21,22, 31].
One of the leading works, modeling age-structured HIV-1 infection treatment with bilinear interaction was proposed by Nelson et al. [24], where the local stability of equilibria was established. More recently, Wang et al. [34], and Yang et al. [35] have introduced an age structured HIV infection model with a nonlinear interaction. In [34], the rate infection function that is considered is \(\dfrac{T(t)V(t)}{1+\alpha V(t)}\) and in [35], the authors considered, the Beddington–DeAngelis infection function, namely \(\dfrac{T(t)V(t)}{1+\alpha _1T(t)+\alpha _2 V(t)}.\) In these two papers, the global asymptotic stability of the equilibria is proved, using suitable Lyapunov functions.
Motivated by these works, we propose and investigate an age-structured HIV infection model with a very general nonlinear infection function,
with the boundary and initial conditions
Here, T(t), i(t, a) and V(t) are the populations of uninfected cells, infected cells with infection age a and free virus at time t respectively. The functions \(\delta (a),\) p(a), are respectively, the age-dependent per capita death, the viral production rate of infected cells with age a. The parameters c, d, are respectively the clearance rate of virus and the death rate of uninfected cells. The parameter A represent the entering flux into the target cells in class T.
The novelty of our work is to propose a general Lyapunov functional that ensures global stability of equilibria; and this is done for a large class of infection functions, some of them being untreated in the literature as the family of functions \(f(T,V)=\dfrac{TV}{1+\alpha V^n}\) with \(0\le n\le 1,\) numerically illustrated in the last example in this paper.
Next, it will be assumed that :
-
The function \(\delta (a) \ge \delta _0>0\) for almost every \(a\ge 0\) and \(\delta \in L_{+}^{\infty }(\mathbb {R}^{+}).\)
-
The function p is assumed to belong to the set \(L_{+}^{\infty }(\mathbb {R}^{+})\setminus {\{0\}}\), and \( p^{max}:=ess\sup _{a\in \mathbb {R}^{+}} p(a)\) for almost every \(a\ge 0\).
-
The parameters A, d and c are supposed to be positive.
We present now our basic assumptions for the infection function f and notations that will be used from now on.
We suppose that the infection response function f satisfies :
-
\(f(\cdot p,V)\) is increasing for \(V> 0.\) Moreover \(f(0,V)=f(T,0)=0\) for all \(T, V\ge 0,\)
-
The function \(\dfrac{\partial f}{\partial V}(.,0)\) is continuous positive on every compact set K.
-
The function f is locally Lipschitz continuous in T and V, with a Lipschitz constant \(L>0,\) i.e. for every \(C>0\) there exists some \(L:=L_C>0\) such that
$$\begin{aligned} |f(T_2,V_2)-f(T_1,V_1)|\le L(|T_2-T_1|+|V_2-V_1|), \end{aligned}$$(1.3)whenever \(0\le T_2,T_1, V_2, V_1\le C.\)
Let us define the functional space \(X:=\mathbb {R}\times L^{1}(\mathbb {R}^{+})\times \mathbb {R}\) equipped with the norm
Moreover, \(X_{+}=\mathbb {R}^{+}\times L_{+}^{1}(\mathbb {R}^{+})\times \mathbb {R}^{+}\) denote the positive cone of X.
Throughout this paper, we denote the probability that an infected cell survives to age a, by \(\Pi (a):\)
and thus
represents the total number of viral particles produced by an infected cell in its life span.
For the model (1.1), the number \(R_0\) of secondary infections produced by a single infected cell during its lifetime [10] is defined by
Next, we set \(\Phi (t,a)=(T(t),i(t,a),V(t)),\) \(\Phi _0(a)=\left( T_0, i_0(a),V_0\right) \)
We begin by stating the following theorem, its proof is standard, see for instance [35].
Theorem 1.1
Assume that (1.3) holds and let us suppose \( (T_0,i_{0}(.),V_0) \in X_{+}.\) Then there exists a unique non-negative solution \( (T,i,V) \in C^{1}(\mathbb {R}^{+})\times C(\mathbb {R}^{+}; L^{1}(\mathbb {R^{+}}))\times C^{1}(\mathbb {R}^{+})\) of problem (1.1)–(1.2). Moreover, we have the following estimates,
for all \(t\ge 0,\) with \(\alpha := \min \{d,\delta _0\}.\) Moreover
and
Finally,
with \(\Lambda :=\dfrac{A}{d+L},\) and L is a Lipschitz constant defined in (1.3).
The rest of this paper is organized as follows: The next section focuses on proving the existence of compact attractor and determining the total trajectories. Then, we will prove that the infection-free equilibrium is globally asymptotically stable whenever \(R_0\le 1.\) Finally, we will investigate the global dynamic of the infection equilibrium, whenever it exists. Observe that the Lyapunov functional that will ensure the global stability is constructed independently of the choice of the infection function, and is valid for almost all usual functions; bilinear or nonlinear as Holling or Beddington.
2 Global compact attractor and total trajectories
We begin by writing the problem (1.2) in the form of a Volterra type equation,
Now, it is not difficult to show the existence of a continuous semiflow
with (T, i, V) is solution of the autonomous problem (1.1)–(1.2).
We choose \(X=\mathbb {R}^{+}\times L^{1}(\mathbb {R}^{+})\times \mathbb {R}^{+}\) endowed with the natural norm \(||(T,i,V)||_X=|T|+||i||_1+|V|.\)
The following theorem state the existence of a compact attractor of all bounded sets of X, (the concept of global attractor is presented in e.g. [19],[28, 30]).
Theorem 2.1
The semiflow \(\Phi \) has a compact attractor \(\mathbf {A}\) of bounded sets of X.
Proof
By Theorem 1.1, the semiflow \(\Phi \) is point-dissipative and eventually bounded on bounded sets on X.
Hence, from Theorem 2.33 in [28], we only need to show the asymptotic smoothness of \(\Phi \) to complete the proof. In order to prove this property, we apply Theorem 2.46 in [28]. We define
as
and \(\Theta _2(t,(T_0,i_0(.),V_0))=(T(t),w_2(t,.),V(t))\) with
Notice that
Now let C be a bounded closed subset of initial data in X, that is forward invariant under \(\Phi .\) First, in view of Theorem 1.1 observe that
with \(\tilde{M}=\max \{\dfrac{A}{\alpha }, ||i_0||_1+T_0\}+\max \{V_0,\dfrac{p^{max}}{c}\max \{\dfrac{A}{\alpha }, ||i_0||_1+T_0\}\}.\)
Then \(\Theta _1\) satisfies,
hence \(\Theta _1\rightarrow 0\) as \(t\rightarrow \infty \) uniformly for all initial data in C. We set
we claim that \(I_h\rightarrow 0\) as \(h\rightarrow 0\) uniformly for all initial data in C. Notice that the second term of \(I_h\) tends to 0 uniformly for all initial data in C. So by setting
and applying the Lipschitz condition of f, namely (1.3) we have
with \(f(T,V)\le f^{\infty }\) for all \((T,V)\in K\) where K is a compact. Now observe that
and
for all \(t\ge 0.\) Therefore we easily conclude that \(I^{1}_h\) tends to 0 as h tends to 0, uniformly for all initial data in C. This completes the proof. \(\square \)
Further, we describe the total trajectories of system (1.1), that are solutions of (1.1) defined for all \(t\in \mathbb {R}.\) These extended solutions play an important role in proving the global asymptotic stability of equilibria.
We consider \(\bar{\phi }\) a total \(\Phi -\)trajectory, \(\bar{\phi }(t)=(T(t),i(t,.),V(t)).\) Then by a straightforward calculation, see also [2, 28] we obtain, for all \(t\in \mathbb {R},\)
Our next result relies on properties of the total trajectory, related to the compact attractor \(\mathbf {A}.\)
Lemma 2.2
For all \((T_0,i_0,V_0)\in \mathbf {A},\) we have,
where L is the Lipschitz constant defined in (1.3) and \(\Gamma \) is a positive constant.
Proof
First, we set
Differentiating this last equation and using the definition of \(\Pi \) we get,
using \(\delta (a)\ge \delta _0,\) and from the equation of T in (2.5),
with \(\alpha :=\min {\{d,\delta _0\}}.\) Now integrating this last inequality over the interval (r, t) we obtain
Letting r goes to \(-\infty \) we get
Moreover from the equation of V in (2.5) we have
thus, a straightforward computation leads to,
Therefore, adding (2.6) and (2.7) we find,
In addition since T and V belong to a compact subset, and f is a continuous function, then there exists a positive constant \(\Gamma \) such that
Finally, concerning the lower bound of T, using the Lipschitz hypothesis of f,
after integration,
This completes the proof.\(\square \)
3 The global stability of the infection-free equilibrium
This section is devoted to prove the global asymptotic stability of the infection-free equilibrium. Throughout this section we suppose that :
-
\(\mathbf {(H1)}\) the function f is concave with respect to V.
We observe that the system (1.2) always has an infection-free equilibrium \((\dfrac{A}{d},0,0).\)
Theorem 3.1
Suppose that (H1) holds. Then, the disease free equilibrium \((\dfrac{A}{d},0,0)\) is globally asymptotically stable whenever \(R_0\le 1.\)
Proof
Let us define the function \(\psi \) as
which is solution of the following problem
Then for \(x:=(T_0,i_0(.),V_0)\in \mathbf A ,\) we consider as the Lyapunov functional
where
and
Let \(\chi : \mathbb {R}\rightarrow \mathbf {A}\) be a total \(\Phi -\)trajectory, \(\chi (t)=(T(t),i(t,.),V(t)),\) \(T(0)=T_0,\) \(i(0,a)=i_0(a),\) and \(V(0)=V_0\) with (T(t), i(t, a), V(t)) is solution of problem (2.5).
Next,
from the expression of i in (2.5) we have
with
Following the same arguments as in the proof of Lemma 9.18 in [28] we can show that \(U_2\) is absolutely continuous and
thus, in view of (3.2) it yields,
from this, and (3.1) we obtain,
Now we analyze \(U':=U_1'+U_2'+\dfrac{V'}{N},\) then by using (2.5), adding and subtracting \(V\dfrac{\partial f}{\partial V}(\dfrac{A}{d},0)\) we obtain
by the definition of \(R_0\) observe that,
thus,
On the other hand, we compute
finally the concavity of f with respect to V ensures that
Hence, using the fact that \(\dfrac{\partial f}{\partial V}(.,0)\) is continuous positive on every compact set K and \(R_0\le 1\) we get
Further, notice that, \(\dfrac{d}{dt}U(\chi (t))=0\) implies that \(T(t)=\dfrac{A}{d}.\) Let Q be the largest invariant set, for which \(\dfrac{d}{dt}U(\chi (t))=0.\) Then in Q we must have \(T(t)=\dfrac{A}{d}\) for all \(t\in \mathbb {R}.\) We substitute this into the equation of T in (2.5) we get \(V(t)=0\) for all \(t\in \mathbb {R}\) and thus, from the equation of i in (2.5) we obtain \(i(t,.)=0\) for all \(t\in \mathbb {R}.\) Then the largest invariant set with the property that \(\dfrac{d}{dt}U(\chi (t))=0\) is \((\dfrac{A}{d},0,0)\) (LaSalle’s Invariant Principle). Now, since \(\mathbf {A}\) is compact, the \(\omega (x)\) and \(\alpha (x)\) are non-empty, compact, invariant and attract \(\chi (t)\) as \(t\rightarrow \pm \infty ,\) respectively. We know that U is constant on the \(\omega (x)\) and \(\alpha (x),\) and thus \(\omega (x)=\alpha (x)=\{(\dfrac{A}{d},0,0)\}.\) Consequently \(\lim \limits _{t\longrightarrow \pm \infty }\chi (t)=(\dfrac{A}{d},0,0)\) and
Since \(U(\chi (t))\) is a decreasing function of t, we obtain \(U(\chi (t))=U(\dfrac{A}{d},0,0)\) for all \(t\in \mathbb {R},\) and thus \(\chi (t)=(\dfrac{A}{d},0,0)\) for all \(t\in \mathbb {R}.\) In particular \((T_0,i_0(.),V_0)=(T(0),i(0,.),V(0))=(\dfrac{A}{d},0,0).\) Therefore the attractor \(\mathbf {A},\) is the singleton set formed by the disease free equilibrium \((\dfrac{A}{d},0,0).\) By Theorem 2.39 in [28], the infection-free equilibrium is globally asymptotically stable.\(\square \)
4 Existence of infection equilibrium states and uniform persistence
In this section, we first ensures the existence of a positive equilibrium states and next, we establish the strongly uniform persistence of the solution to problem (1.2).
Lemma 4.1
Let \(\lim \limits _{V\rightarrow 0^{+}}\dfrac{f(\dfrac{A}{d},V)}{f(T,J)}>1\) for \(T\in [0,\dfrac{A}{d}).\) Then, if \(R_0>1,\) system (1.1)–(1.2) has positive equilibrium states .
Proof
An infection equilibrium is a fixed point of the semiflow \(\Phi \),
From (2.1), we obtain
and
First remark that if \(i^{*}(a)\) is given by the first case in (4.1), it also satisfies the second case. Indeed, for \(t<a<2t\) we have
and thus
Now we proceed by iteration in order to prove the result. Therefore
Combining the equations (4.2) and (4.3) we get,
with N is defined in (1.5). Following the same arguments as [17, 18] we prove the existence of positive equilibrium states.\(\square \)
We emphasis now on the uniform persistence see for instance [13, 19, 28, 29]; for this purpose we apply Theorem 5.2 in [28].
We first make the following assumptions on the infection function f.
We suppose that there exists a positive equilibrium \((T^{*},V^{*})\) verifying (4.4) such that for all \(T>0\) we have
There exists \(\varepsilon >0\) and there exists \(\eta >0\) such that for all \(T\in [\dfrac{A}{d}-\varepsilon ,\dfrac{A}{d}+\varepsilon ]\) we have
for all \(0< V_1 \le V_2\le \eta .\)
Finally we suppose that
Remark 4.2
If the function f is increasing and concave with respect to V then (4.5) and (4.6) are clearly verified.
We define a persistence function \(\rho : X\rightarrow \mathbb {R}^{+}\) by
then
with \(x=(T_0,i_0(.),V_0).\)
The following lemma affirm that the hypothesis (H1) in Theorem 5.2. [28] holds.
Lemma 4.3
Under assumption (4.7), the function \(\rho (\phi (t))\) is positive on \(\mathbb {R},\) with \(\phi \) is a total \(\Phi -\)trajectory defined in (2.5).
Proof
We claim that \(V(t)>0\) for all \(t\in \mathbb {R}\) with V is defined in (2.5). Indeed, suppose first that there exists \(r\in \mathbb {R}\) such that \(V(t)=0\) for all \(t\le r.\) So for \(t>r\) we have
We show, in this case, that \(V(t)=0\) for all \(t\in \mathbb {R}.\) Assume by contradiction that there exists \(t_1>0\) such that \(V(t_1)=0,\) \(V'(t_1)>0\) and \(V(t)=0\) for all \(t\le t_1.\) Thus
consequently, \(V(t)=0\) for all \(t\in \mathbb {R},\) in addition, using the definition of i in (2.5) we get \(V(t)+\int _0^{\infty }i(t,a)da=0\) for all \(t\in \mathbb {R}\). However from (4.7), this is not possible, so there exists sequence \((t_n)_n\) and \(t_n\rightarrow -\infty \) such that \(V(t_n)>0.\) On the other hand remark that, by integration the equation of V over \((t_n,t)\),
now it is easily to conclude that \(V(t)>0\) for all \(t\in \mathbb {R}.\) Hence we conclude.\(\square \)
Now we are ready to prove the strong uniform persistence of the disease.
Theorem 4.4
Suppose (4.6), (4.7) hold. Then there exists some \(\varepsilon >0\) such that
for all nonnegative solutions of (1.2) provided that \(R_0>1.\)
Proof
By Theorem 5.2. in [28], and Lemma 4.3, the solution of problem (1.1)–(1.2) is strongly uniformly persistent if it is weakly uniformly persistent.
Suppose that the disease is not uniformly weakly persistent, that is, there exists an arbitrarily small \(\varepsilon >0\) such that
so,
Next, we set \(\lim \inf \limits _{t\rightarrow \infty }T(t)=T_{\infty },\) using the fluctuation method see for instance [30], there exists a sequence \((t_k)_k\) such that \(\lim \limits _{t_k\rightarrow \infty }T'(t_k)= 0\) and \(\lim \limits _{t_k\rightarrow \infty }T(t_k)=T_{\infty }.\)
First, in view of the continuity of the function f we have
Combining this with the equation of T in (1.1), we have (for large t)
therefore,
with \(\theta (\varepsilon )=\dfrac{\varepsilon }{d}\). Now since \(R_0>1,\) then there exists \(\varepsilon _1>0\) so small and \(\lambda :=\lambda _{\varepsilon _1}>0\) such that
We set
On the other hand, we introduce the following auxiliary problem
The solution of this problem is given by
In addition, remarking that the function \(\phi \) is uniformly bounded, more precisely
Now we analyze de derivative of \(I_1(t):=\int _0^{\infty }\phi (a)i(t,a)da+V(t),\) so by a simple calculation using (4.9) we obtain,
Now for t so large and the monotonicity of f with respect to T, we have,
On the other hand, since \(V(t)\le \varepsilon _1\) then using (4.6) we get
Therefore
with \(\beta _{\varepsilon _1}=\min \{\alpha _{\varepsilon _1},\lambda \}\) where \(\alpha _{\varepsilon _1}\) is defined in (4.8).
Finally
hence due to (4.7) \(I_1(t)\rightarrow +\infty \) which is a contradiction with the boundedness of V and \(\int _0^{\infty }\phi (a)i(t,a)da\). The result is reached. \(\square \)
Let \(X_0\) be a subset defined as
From Theorem 5.7 in [28] we have the following result
Theorem 4.5
Under the assumptions (4.6), (4.7), there exists a compact attractor \(\mathbf {A_1}\) that attracts all solutions with initial condition belonging to \(X_{+}\setminus X_0.\) Furthermore \(\mathbf {A_1}\) is \(\rho -\) uniformly positive, i.e., there exists some \(\gamma >0\) such that,
5 The global stability and uniqueness of the infection steady state
In this section, we discuss the global stability of the infection equilibrium \((S^{*},v^{*}(.),i^{*}(.))\) of system (2.5). Before stating the main result of this section, we need the following estimate, which guarantees that all solutions of (2.5) with initial data satisfying (4.7), are bounded away from 0. We will use a simple modification of the idea proposed in the proof of Claim 5.3 in [9].
Proposition 5.1
Assume (4.6), (4.7) hold. Then, there exist positive constants \(\alpha \) and \(\Gamma \) such that, for all \((T_0,i_0(.),V_0)\in \mathbf {A_1},\)
Proof
Since \(\mathbf {A_1}\) is invariant, then, there exists a total trajectory \(\Psi :\mathbb {R}\rightarrow \mathbf {A_1},\) \(\Psi (t)=(T(t),i(t,.),V(t))\) with \(T(0)=T_0,\) \(i(0,a)=i_0(a)\) and \(V(0)=V_0.\) Suppose by contradiction that \(\liminf \limits _{t\rightarrow \infty }V(t)=0,\) then there exists a sequence \(t_n\rightarrow \infty \) such that \(V(t_n)\rightarrow 0.\) We set \(T_n(t)=T(t+t_n),\) \(V_n(t)=V(t+t_n)\) and \(i_n(t,.)=i(t+t_n,.).\) Then up to a subsequence one may assume that \((T_n(t),V_n(t),i_n(t,.))\rightarrow (\tilde{T}(t),\tilde{V}(t),\tilde{i}(t,.))\) locally uniformly with \(((\tilde{T}(t),\tilde{V}(t),\tilde{i}(t,.))\) is a solution of (2.5) such that \(\tilde{V}(0)=0.\) Now employing the same argument as in the proof of lemma 4.3 we reach a contradiction with (4.10) and \(\tilde{V}(0)=0.\) Moreover,
due to Lemma 2.2, and (4.3) there exists \(\Gamma >0\) such that,
\(\square \)
Now we are able to state the main result of this section.
Theorem 5.2
Under the assumptions of (4.5), (4.6), (4.7), the problem (2.5) has a unique positive infection equilibrium \((T^{*},i^{*}(a),V^{*})\) which is globally asymptotically stable in \(X_{+}\setminus X_0\).
Proof
Let \(\Psi : \mathbb {R}\rightarrow \mathbf {A_1}\) be a total \(\Phi -\)trajectory, \(\Psi (t)=(T(t),i(t,.),V(t)),\) \(T(0)=T_0,\) \(i(0,a)=i_0(a),\) and \(V(0)=V_0,\) with (T(t), i(t, a), V(t)) is solution of problem (2.5).
We set,
and
with N is defined in (1.5).
Then, for \(x:=(T_0,i_0(.),V_0)\in \mathbf {A_1},\) we consider the following Lyapunov functional
with,
and
By analyzing the derivative of \(W_1,\) using the definition of the positive steady state in (4.4) we have,
Now, concerning \(W_2.\) Following the same arguments as in the proof of Lemma 9.18 in [28] and (2.5) we find,
Using the definition of H we have
thus,
Next, for \(W_3\) we get,
From (4.4) namely, \(cV^{*}=Nf(T^{*},V^{*})\) we obtain,
hence, for \(W=W_1+W_2+W_3\) we get,
Observe that the function \(-ln(x)-\dfrac{1}{x}+1\le 0,\) for all \(x>0,\) then the first two terms of the above equation are negative. Next we will claim that the last term is negative. For this we set,
Let us consider a time t such that \(\dfrac{V(t)}{V^*}<1,\) then, from the hypothesis (4.5) we have,
Hence, since \(H(1)=0\) and H is decreasing in (0, 1), it yields,
thus,
Now since H is a convex function then,
so,
For other values of t i.e. \(\dfrac{V(t)}{V^*}>1,\) again from (4.5), and by taking into account that H is an increasing function over \((1,\infty )\) we also arrive to prove that \(D(t)\le 0\). Consequently,
Further, \(\dfrac{d}{dt}V(\Psi (t))=0\) implies that \(T(t)=T^{*}.\) Now we look for the largest invariant set Q for which \(\dfrac{d}{dt}V(\Psi (t))=0.\) In Q, we must have \(T(t)=T^{*}\) for all \(t\in \mathbb {R}.\) First, according to equations of T and \(T^{*}\) in (2.5) and (4.1) it is clear that,
and finally in view of (2.5),
Following the same arguments as in the proof of Theorem 3.1 we conclude the global asymptotic stability of the infection equilibrium. Uniqueness is a direct consequence of the fact that \(\frac{d}{dt}V(\Psi (t))=0\) holds only on the line \(T=T^{*}.\) This ends the proof of the theorem.\(\square \)
6 Example and numerical simulations
In this section, we propose some mathematical models describing a virus dynamics to illustrate the different results obtained in the previous sections. We also realize some numerical simulations for each example.
Example 1
Let f be the Beddington–DeAngelis function
We replace the function f in system (1.1), we obtain
where \(\beta , \alpha _1\) and \(\alpha _2\) are positive constants, we denote by \(\beta \) the rate of infection, \(\alpha _1\) and \(\alpha _2\) represent the Beddington–DeAngelis infection rate, for more details about this system, we refer to [35] (Figs. 1, 2).

a The evolution of the solution T and V with respect to time. b The age distribution of i with respect to time and infection age

a The evolution of the solution T and V with respect to time. b The age distribution of i with respect to time and infection age
The function f satisfies all hypothesis defined in the first part, and by theorems 3.1 and 5.2, we obtain the following result:
-
If \(R_0 \leqslant 1\), the disease free equilibrium is globally asymptotically stable.
-
If \(R_0>1\), the positive infection equilibrium is globally asymptotically stable.
The number \(R_0\) of this model is defined by
We give the following values to some parameters to do the numerical simulations of this model,
with the initial conditions
We take different values of \(\tau \) to simulate all possible cases of the asymptotic dynamics of system (1), when \(\tau =6\), then \(R_0=0.6676 > 1,\)
And when \(\tau =1\), then \(R_0=1.8146 > 1\),
Example 2
We choose the following function f for our second example
the function h being defined as

Graphical representation of the function h with \(\beta =0.1\) and \(\alpha =0.2\)

a The evolution of the solution T and V with respect to time. b The age distribution of i with respect to time and infection age
for a special case \(n=1\) and \(n=0\), function h is similar to Holling type I and II functional response (Figs. 3, 4, 5).
We can see that the function f satisfies all the assumptions made in the previous sections.
The number \(R_0\) is defined by
The choice of the parameter \(A=2, \quad d=0.1, \quad c=1, \quad \delta =0.3, \quad \beta = 0.1, \quad \alpha _1=0.2 \text { and } n=0.8.\) Letting \(\tau \) free in
with the initial conditions
We change the values of \(\tau \) in order to have \(R_0 \leqslant 1\) or to have \(R_0>1.\) If we choose \(\tau =5\), then \(R_0=0.7162 < 1\),
And if we choose \(\tau =1\), then \(R_0=2.3779 > 1\),

a The evolution of the solution T and V with respect to time. b The age distribution of i with respect to time and infection age
Remark 6.1
We made the choice to let \(\tau \) free, and to discuss with respect to its different values. One can choose to fix \(\tau \) and let any other parameter free, this will lead us the same conclusion provided that either \(R_0\le 1\) or \(R_0>1.\)
7 Conclusion
In this paper we have considered an age-structured HIV infection model with a very general nonlinear infection function,
with the boundary and initial conditions
This kind of problems is widely treated in the literature for particular choices of f, our main contribution consists in considering a very general class of infection functions f(T(t), V(t)), observe that the assumptions made on f are not restrictive, as almost all types of usual infection functions fulfill them. We propose a Lyupunov function for these problems, confirming the results obtained by previous works and showing new ones as for the case of \(f(T,V)=\dfrac{TV}{1+\alpha V^n}\) with \(0\le n\le 1.\)
Even if a lot is biologically known about the virus evolution in its current form, however the interaction between uninfected cells and the virus is a very complicated phenomenon, in addition a virus can mutate, and develop other ways to contaminate cells than those known till now, hence the relevance of considering a function f in its most general form to prevent any possible behavioral change of the virus.
Our results are illustrated by numerical simulations, we have chosen some examples that are untreated in the existing the literature, we made the distinction between the cases \(R_0 \le 1\) and \(R_0 >1.\)
As a good perspective to the present work, one can include a treatment or a vaccination function with different strategies of vaccination; and study the asymptotic behavior of the solutions to the considered model.
References
Althaus, C.L., De Boer, R.J.: Dynamics of immune escape during HIV/SIV infection. PloS Comput. Biol. 4, e10000103 (2008)
Bentout, S., Touaoula, T.M.: Global analysis of an infection age model with a class of nonlinear incidence rates. J. Math. Anal. Appl. 434, 1211–1239 (2016)
Brauer, F., Castillo-Chavez, C.: Mathematical Models in Population Biology and Epidemiology. Springer, New York (2000)
Brauer, F., Shuai, Z., van den Driessche, P.: Dynamics of an age of infection cholera model. Math. Biosci. Eng. 10(5–6), 1335–1349 (2013)
Cai, L., Martcheva, M., Li, X.-Z.: Epidemic models with age of infection, indirect transmission and incomplete treatment. Discrete Contin. Dyn. Syst. B 18, 2239–2265 (2013)
Castillo-Chavez, C., Hethecote, H.W., Andreasen, V., Levin, S.A., Liu, W.M.: Epidemiological models with age structure, proportionate mixing and cross-immunity. J. Math. Biol. 27, 240–260 (1989)
Culshaw, R.V., Ruan, S.: A delay-differential equation model of HIV infection of \(CD4^{+}\) T-cells. Math. Biosci. 165, 27–39 (2000)
De leenheer, P., Smith, H.L.: Virus dynamics: a global analysis. SIAM J. Appl. Math. 63, 1313–1327 (2003)
Demass, R.D., Ducrot, A.: An age-strutured within-host model for multistrain malaria infections. SIAM J. Appl. Math. 73, 572–593 (2013)
Diekmann, O., Heesterbeek, J.A.P.: Mathematical Epidemiology of Infectious Diseases: Model Building, Analysis and Interpretation. Wiley, Chichester (2000)
Feng, Z., Thieme, H.R.: Endemic models with arbitrarily distributed periods of infection I: fundamental properties of the model. SIAM J. Appl. Math. 61(3), 803–833 (2000)
Feng, Z., Thieme, H.R.: Endemic models with arbitrarily distributed periods of infection II: fast disease dynamics and permanent recovery. SIAM J. Appl. Math. 61(3), 983–1012 (2000)
Hale, J., Waltman, P.: Persistence in infinite dimensional systems. SIAM J. Math. Anal. 20, 388–395 (1989)
Huang, G., Ma, W., Takeuchi, Y.: Global properties for virus dynamics model with Beddington–DeAngelis functional response. Appl. Math. Lett. 22, 1690–1693 (2009)
Huang, G., Takeuchi, Y., Ma, W.: Lyapunov functionals for delay differential equations model of viral infections. SIAM J. Appl. Math. 70, 2693–2708 (2010)
Huang, G., Liu, X., Takeuchi, Y.: Lyapunov functions and global stability for age-structured HIV infection model. SIAM J. Appl. Math. 72, 25–38 (2012)
Korobeinikov, A.: Lyapunov functions and global stability for SIR and SIRS epidemiological models with non-linear transmission. Bull. Math. Biol. 68, 615–626 (2006)
Korobeinikov, A., Global, A.: Properties of infectious disease models with nonlinear incidence. Bull. Math. Biol. 69, 1871–1886 (2007)
Magal, P., Zhao, X.-Q.: Global attractors and steady states for uniformly persistent dynamical systems. SIAM J. Math. Anal. 37, 251–275 (2005)
Magal, P., McCluskey, C.C., Webb, G.F.: Lyapunov functional and global asymptotic stability for an infection-age model. Appl. Anal. 89(7), 1109–1140 (2010)
McCluskey, C.C.: Global stability for an SEIR epidemiological model with varying infectivity and infinite delay. Math. Biosci. Eng. 6, 603–610 (2009)
McCluskey, C.C.: Complete global stability for a SIR epidemic model with delay-distributed or discrete. Nonlinear Anal. 11, 55–59 (2010)
Nelson, P.W., Perelson, A.S.: Mathematical analysis of delay differential equation models of HIV-1 infection. Math. Biosci. 179, 73–94 (2002)
Nelson, P.W., Gilchrist, M.A., Coombs, D., Hyman, J.M., Perelson, A.S.: An age-structured model of HIV infection that allows for variations in the production rate of viral particles and the death rate of productively infected cells. Math. Biosci. Eng. 1, 267–288 (2004)
Nowak, M.A., May, R.M.: Virus Dynamics: Mathematical Principles of Immunology and Virology. Oxford University Press, Oxford (2000)
Perelson, A.S., Nelson, P.W.: Mathematical analysis of HIV-1 dynamics in vivo. SIAM Rev. 41, 3–44 (1999)
Rong, L., Feng, Z., Perelson, A.S.: Mathematical analysis of age-structured HIV-1 dynamics with combination antiretroviral therapy. SIAM J. Appl. Math. 67, 731–756 (2007)
Smith, H.L., Thieme, H.R.: Dynamical Systems and Population Persistence. Graduate studies in mathematics. AMS, Providence (2011)
Thieme, H.R.: Uniform persistence and permanence for nonautonomus semiflows in population biology. Math. Biosci. 166, 173–201 (2000)
Thieme, H.R.: Mathematics in Population Biology. Princeton University Press, Princeton (2003)
Thieme, H.R.: Global stability of the endemic equilibrium in infinite dimension : Lyapunov functions and positive operators. J. Differ. Equ. 250, 3772–3801 (2011)
Thieme, H.R., Castillo-Chavez, C.: How may infection-age-dependent infectivity affect the dynamics of HIV/AIDS? SIAM J. Appl. Math. 53, 1447–1479 (1993)
Vargas-De-Leon, C., Esteva, L., Korobeinikov, A.: Age-dependency in host-vector models: the global analysis. Appl. Math. Comput. 243, 969–981 (2014)
Wang, J., Zhang, R., Kuniya, T.: Mathematical analysis for an age-structured HIV infection model with saturation infection rate. Electron. J. Differ. Equ. 33, 1–19 (2015)
Yang, Y., Ruan, S., Xiao, D.: Global stability of an age-structured virus dynamics model with Beddington–DeAngelis infection function. Math. Biosci. Eng. 12(4), 859–877 (2015)
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article

Cite this article
Frioui, M.N., Miri, S.Eh. & Touaoula, T.M. Unified Lyapunov functional for an age-structured virus model with very general nonlinear infection response. J. Appl. Math. Comput. 58, 47–73 (2018). https://doi.org/10.1007/s12190-017-1133-0
Received:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s12190-017-1133-0
Keywords
- Age structure
- Virus dynamics model
- General nonlinear infection function
- Compact attractor
- Total trajectories
- Lyapunov functional
- Global stability