
Abstract
Purpose
A novel tumor suppressor gene CKLF-like MARVEL transmembrane domain-containing member 3 (CMTM3) is reduced or undetectable in many kinds of cancers and relates tumor malignant features. We detected its role in prostate cancer for possibility of target therapy as accumulating evidence has shown that CMTM3 is a promising tumor suppressor gene (TSG) for gene therapy.
Methods
The expression of CMTM3 detected in prostate tissue microarray, specimens and cell lines were evaluated by immunohistochemistry and semi-quantitative PCR and Western blot, respectively. After being transfected with CMTM3 adenovirus or vector (mock), the proliferation and migration and invasion of LNCaP cells were detected by transwell assay and matrigel assay, respectively. Furthermore, the effects of CMTM3 on tumor growth were performed in nude mice xenograft in vivo.
Results
We found CMTM3 was reduced in PCa tissues and cells compared with BPH tissues, and its expression in PCa tissues was related to the Gleason score. Moreover, after being transfected with adenovirus, ectopic expression of CMTM3 in LNCaP cells led to significant inhibition of cell proliferation and migration and invasion compared with the control (P < 0.05), which may be attributed to decreased Erk1/2 activity as p-Erk1/2 was remarkably reduced when CMTM3 was overexpressed. Finally, restoration of CMTM3 significantly suppressed xenograft tumor growth in vivo (P < 0.01).
Similar content being viewed by others
Avoid common mistakes on your manuscript.
Introduction
Prostate cancer (PCa) is the most common malignant tumor and the second leading cause of cancer-related death among males in the USA [1]. And the incidence of prostate cancer is increasing rapidly in Asian countries especially in China, which is now economically fast expanding and population aging. It is a critical public health problem and remains incurable in the metastatic setting with mortality that usually occurs as a result of castration-resistant disease. Therefore, better understanding of molecular mechanisms that are involved in regulating prostate tumor development and metastasis will help to optimize survival and quality of life for the patients. In light of a growing array of existing and novel treatment options, we are striving to discover new targets for CRPC and relieve the pain those patients are suffering from.
CKLF-like MAL and related proteins for vesicle trafficking and membrane link (MARVEL) transmembrane domain-containing family (CMTM) is a novel family of proteins consisting of nine members, chemokine-like factor (CKLF) and CMTM1–CMTM8. The members of CMTM link to chemokines and the transmembrane 4 superfamily (TM4SF) has important functions in the immune system, male reproductive system, and tumorigenesis [2, 3]. CKLF-like MARVEL transmembrane domain-containing member 3 (CMTM3) belongs to the chemokine-like factor gene superfamily located in a cluster on chromosome 16q22. CMTM3 protein contains one leucine zipper and two LXXLL motifs which localized to the cytoplasm and serves as a scaffold for proteins in the endoplasmic reticulum and the nuclear membrane [4]. Previous studies indicated that CMTM3 is silenced or down-regulated in gastric, breast, nasopharyngeal, esophageal, colon and renal carcinomas and its expression inversely correlates with grade and phase in prostate cancer when combined with IL-30 study [5–7]. The re-expression of CMTM3 in tumor cells lacking its expression leads to the suppression of cell growth and migration and invasion but its role in cancer development and progression has not been clearly defined until now.
In this study, we observed that CMTM3 was proportionally down-regulated in prostate cancer tissues and identified the effects of CMTM3 restoration on LNCaP cells proliferation and migration and invasion in vitro and proliferation in nude mice xenograft model in vivo.
Materials and methods
Chemicals, reagents and antibodies
The adenovirus carrying the CMTM3 gene (Ad-CMTM3) and the empty adenovirus (Ad-null) and Rabbit antihuman CMTM3 polyclonal antibody were kindly provided by Dr. Han’s Lab and (Center for Human Disease Genomics, Peking University, Beijing, PR China). Rabbit antihuman GAPDH monoclonal antibody was purchased from Cell Signaling Technology (Beverly, MA, USA). Antibody specific for Erk1/2, p-Erk1/2, β-actin and GAPDH were from Cell Signaling Technologies (Danvers, MA, USA). Tissue microarray was purchased from Shanghai outdo biotech co., LTD. Other prostate cancer tissues were obtained from patients undergoing radical prostatectomy of the prostate at the Peking University People’s Hospital with the patients’ consent and institutional ethics approval.
Immunohistochemistry
A tissue microarray included 180 spots of human prostate tissue samples and adjacent BPH (benign prostatic hyperplasia) tissues (99 PCa and 81 BPH tissues) and 9 other BPH and 13 PCa surgical specimens in Peking University People’s Hospital were used with patients’ consent and ethics committee’s approval. The tissue microarray and samples were dewaxed, rehydrated and blocked with 10 % normal goat serum for 35 min and incubated with 0.5 μg/ml purified anti-CMTM3 in a humidified chamber at 4 °C overnight. Immunodetection was performed using the Envision TM ABC Kit (Gene Tech Company Limited, Shanghai, China). Previously established immune histochemical scoring system for prostate carcinomas was used after counterstained with hematoxylin and then dehydrated and mounted [8]. The extent and intensity of CMTM3 staining were evaluated by two experienced observers, independently and without clinical data and estimated in a consecutive analysis to ensure maximal internal consistency. The overall results of CMTM3 staining were assigned as “low expression” when the sum was 1–3, and “high expression” when 3–6 (except 3). All evaluations were conducted using a LEICA DM4000B/M microscope.
Cell culture and infection
Two human metastatic androgen-independent cell lines DU145, PC3 were obtained from the American Type Culture Collection (ATCC, Manassas, VA, USA). LNCaP was purchased from the Cell Resource Center of Peking Union Medical College (Beijing, China). Cell culture medium and reagents were from Invitrogen (Carlsbad, CA, USA). Cells were maintained in RPMI 1640, 10 % FBS, penicillin (100 U/ml) and streptomycin (100 μg/ml). The normal epithelial prostate cell line RWPE-1 and its complete growth medium Keratinocyte Serum-Free Medium (K-SFM), Invitrogen (GIBCO), with each of the two additives was required to grow this cell line [bovine pituitary extract (BPE) and human recombinant epidermal growth factor (EGF)]. Cell lines above were cultured in a humidified atmosphere of 5 % CO2 air. Cells were infected with ad-CMTM3 or vector-containing adenovirus as described [3, 9]. Infection was accomplished by exposing the cells to adenovirus at the required multiplicity of infection (MOI) in serum-containing media for 1–4 days. The efficiency of infection was monitored by ad-pEGFP. Cells with >75 %infection efficiency were used for further experiments.
Cell migration and invasion assay
The migration assay was done in 24-well transwell (corning, USA) containing a polycarbonate filter (8 μm pore size, BD, USA). Conditioned medium 500 μL containing 10 % fetal bovine serum was added to the bottom part of the chamber. The cells were trypsinized and washed twice, and 200 μL of the suspension (5 × 104 cells in serum-free RPMI 1640) was added to the upper well. Then the cells were incubated in a 5 % carbon dioxide incubator at 37 °C for 24 h. After incubation, attached but not migrated cells in the upper chamber were scraped off and the migrated cells on the bottom of the filter were fixed with methanol and stained with crystal violet, counted and photographed. Each experiment was run in triplicate. The number of cells was counted in five high-power (magnification, ×400) randomized fields under a microscope. Results were obtained from at least three individual experiments. The invasion assay was done by polycarbonate filter covered with matrigel upper well (8 μm pore size, BD, USA) and 200 μL of the suspension (1 × 105 cells in serum-free RPMI 1640) was added to the upper well and the others are same as above.
Cell proliferation
LNCaP cells were infected as above, and cell proliferation was measured at 24, 48 and 72 h using the Cell Counting Kit-8 (CCK-8 invitrogen, USA) following the manufacturers’ instruction. The spectrophotometric absorbance at 490 nm was measured using a microplate reader. The data are reported as the mean of at least three independent experiments.
Cell counting assay
For the further analysis of cell proliferation, cell counting assay was done in 12-well plates with LNCaP cells infected as above, and cells were measured at 2, 4, 6 days after being trypsinized and neutralized using cell counting chamber under microscope carefully, at least three times for group Ad-CMTM3 and Ad-null, and the data are reported as the mean of at least three independent experiments.
Protein extraction and Western blot analysis
Western blotting was carried out as described previously [10]. For the detection of CMTM3, 100 μg total protein of each sample was loaded.
Semi-quantitative reverse transcription-PCR
The isolation of total RNA and reverse transcription were performed as previously described [11]. The semi-quantitative PCR analysis of CMTM3 was performed using specific primers, and GAPDH was used as the internal control. CMTM3 and GAPDH primers were used as described previously [12]. The assay was conducted in triplicate using different primer sets and the relative expression level of CMTM3 was determined by using the 2DDCt method [13].
Tumor xenograft model in nude mice
Animal experimental protocols were approved by Peking University Institutional Animal Care and Use Committee. Twelve male BALB/c nude mice (6 weeks) were obtained from Peking University Experimental Animal Center with similar weight. LNCaP cells (4 × 106) were removed by trypsinization, washed twice with PBS, and then injected into mice with ad-CMTM3 or vector (MOI = 50) subcutaneously into the left dorsal flank of the nude mice (Day0). Once palpable tumors were formed, tumor growth was monitored as described [14]. There were six mice in the Mock group and six in the CMTM3 group. The animals were raised in specific pathogen-free experimental animal rooms with free access to water and were fed ad libitum. One mouse in each group died in the middle of the experiment. After 4 weeks, 10 mice were humanely killed by carbon dioxide narcosis (Day28). Tumors were measured, weighed and photographed and part of the tumors was excised for protein extraction and Western blot analysis. The rest of specimens were fixed in formalin and embedded in paraffin for IHC staining.
Statistical analysis
Statistical analysis was performed by statistical package of the social sciences (SPSS) software 17.0 (SPSS Inc, Chicago, IL, USA). Data were presented as values of mean ± SEM. Statistical analysis was carried out with Chi-square test and Student’s test. Values of P < 0.05 were considered as statistically significant difference.
Results
CMTM3 expression is down-regulated in PCa tissues and lower in LNCaP cell line
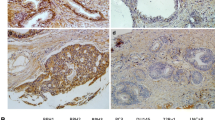
In BPH (benign prostatic hyperplasia) tissues, CMTM3 expression was mainly detected in the membrane of the prostatic epithelia cells, but in PCa tissues with high CMTM3 expression, it was mainly located in the cytoplasm of cancer cells (Fig. 1a). It indicated that 60 % (54/90) of BPH tissues had high CMTM3 expression. However, CMTM3 was decreased markedly in 56.25 % (63/112) of PCa tissues (Table 1). There was a significant difference in the ratio of high expression of CMTM3 between PCa (49/112, 43.75 %) and BPH tissues (54/90, 60.00 %) (P < 0.05). The expression of CMTM3 had no correlation with the age, clinical stage status. However, when compared with the expression levels of CMTM3 in PCa tissues with different Gleason scores, we found that there was a significant difference (P = 0.0307) between the well and moderately differentiated tumors (GS ≤ 7) and the poorly differentiated tumors (GS > 8). The percentages of high-expression cases were 42.65 vs 22.73 %, which suggested that CMTM3 expression might be related with the malignancy of prostate cancer.

Expression patterns of CMTM3 in prostate tissues and cells. a Representative images from immunohistochemical staining of CMTM3 protein in BPH (c, d) and PCa tissues (a, b). a, c Low expression. b, d High expression. b CMTM3 expression in PCa cell lines
To better understand the discovery, semi-quantitative reverse transcription-PCR and Western blot was done to detect the CMTM3 expression in PCa cell lines. As shown in Fig. 1b, CMTM3 expression in LNCaP cell lines is significantly lower than other cell lines.
Re-expression of CMTM3 inhibits cell growth and cell migration and invasion
The down-regulation in PCa samples and silencing of CMTM3 in LNCaP cells indicated that CMTM3 might be a functional tumor suppressor in carcinogenesis [15, 16]. To investigate the growth-inhibitory effect of CMTM3, the re-expression of CMTM3 in the transiently infected LNCaP cells was confirmed by Western blotting (Fig. 2a). As shown in Fig. 2b, CCK8 assay and cell counting assay revealed that cell growth was significantly decreased in Ad-CMTM3-infected LNCaP cells compared with Ad-null-infected control cells.

Ectopic expression of CMTM3 inhibited cell proliferation and migration, invasion of androgen-dependent LNCaP cells. a Re-expression of CMTM3 in the transiently infected LNCaP cells was confirmed by Western blotting. b Cell growth was decreased in Ad-CMTM3-infected LNCaP cells. Data represent the mean ± SEM of OD570 of at least three independent experiments. c Ectopic CMTM3 expression reduced cell migration and invasion ability. d Protein expressions of CMTM3, Akt, p-Akt, Erk1/2, p-Erk1/2 and EGFR were detected by Western blot (*P < 0.05)
In addition, the effect of CMTM3 on cell motility and invasion was examined by a transwell chamber assay and matrigel assay, respectively. Ectopic CMTM3 expression led to significantly reduced cell migration and invasion ability compared to Ad-null-infected cells (Fig. 2c).
To further explore the molecular mechanisms underlying the tumor cell growth, migration and invasion inhibition by CMTM3, several proteins which may participate in the process were assessed by Western blot. There were significant decreased protein levels of phosphorylated Erk1/2 Ad-CMTM3-infected LNCaP cells compared with Ad-null-infected control cells. Results suggest that CMTM3 may repress PCa cells through regulating Erk1/2 signaling, which is one of the most important pathways in tumor progression (Fig. 2d) [17].
Restoration of CMTM3 inhibits xenograft tumor growth in vivo
To confirm the in vivo tumor-suppressive ability of CMTM3, tumor formation in nude mice was induced by injecting LNCaP cells (Table 2, Fig. 3a). The solid tumor of the CMTM3 group had a slower growth curve than the control group (Fig. 3b). In addition, the weight of tumors of the CMTM3 group was smaller than the Mock group (P < 0.05; Fig. 3c). These results indicated that tumor growth may be suppressed in the animals with restoration of CMTM3.

Restoration of CMTM3 inhibited tumor growth in vivo. a Representative images of the xenograft tumors and re-expression of CMTM3 was confirmed by Western blotting ensuring that it would be possible abound on this restoration of CMTM3. Imunohistochemical staining of CMTM3 in tumor tissues after ad-CMTM3 or vector injection. b,c CMTM3 effectively suppressed tumor growth in nude mice. Data are presented as the mean tumor volume and weight ± SE of the surviving animals in each groups. P < 0.01 groups had five mice available
Discussion
CMTM3 is located at 16q22.1, an important tumor suppressor locus with pathogenesis of multiple carcinomas [18]. Cell migration and invasion are two essential steps in the progression of cancer metastasis [19]. Our findings strongly indicate that CMTM3 plays important roles in migration and invasion of PCa cells. Previous study showed that CMTM3 was frequently silenced in many carcinoma cell lines and primary tumors. Re-expression of CMTM3 inhibited tumor cell colony formation and viability, and induced tumor cell apoptosis through caspase-3 activation [20]. The molecular mechanism of how CMTM3 inhibits tumor cell growth and migration is unclear. As a member of CMTM family, the CMTM3 protein has predicted MAL-related proteins for vesicle trafficking and membrane link (MARVEL) domain. MARVEL, a common feature of CMTM families, is a membrane-associating domain and may be a part of the machinery of membrane apposition events [21] such as transport vesicle biogenesis, neurotransmitter secretion and polarized membrane trafficking.
In the present study, the expression of CMTM3 was reduced in most of the PCa tissues (56.25 %). More importantly, CMTM3 expression had no correlation with patients’ clinical stage and metastasis status in these cases, but there was a significant difference in its expression between the two groups of patients with different Gleason scores, supporting that CMTM3 may be involved in the progression of PCa.
Exogenous CMTM3 expression in silenced and androgen-dependent cancer cells LNCaP dramatically suppressed not only tumor cell proliferation, but also their migration, invasion abilities through attenuating Erk1/2 activity. Moreover, introducing the CMTM3 gene into xenograft tumor growth in vivo slowed down tumor growth. As a primary tumor cell lines of LNCaP, CMTM3 may be used as a new alternative of castration, which was typical but incurable way of prostate cancer. Previous studies showed that the down-regulation of tumor suppressors is associated with transcriptional inhibition through the induction of repressive epigenetic modifications in the promoter, including DNA methylation and histone modification [22]. In sum, for the first time, our study shows the function of CMTM3 and the relationship between its expression and clinical features in prostate cancer. CMTM3 may be used as a marker for the treatment for prostate cancer, leading to application of a new PCa therapeutic target.
Conclusions
CMTM3 is significantly down-regulated or undetectable in PCa tissues and LNCaP cell lines. Restoration of CMTM3 inhibits PCa cell proliferation and migration, invasion in vitro and tumor growth in vivo. Thus, CMTM3 acts as a tumor suppressor in PCa extending the current knowledge of CMTM’s function in tumorigenesis and CMTM3’s potential as a target of gene therapy. Further investigations are warranted to bolster the present conclusion due to the limited tissue samples available and research technology available. However, further study of molecular mechanisms of the role CMTM3 plays in the development of prostate cancer is indeed not enough in our study, results above reveal CMTM3 acts as a potential tumor suppressor in prostate cancer and targeting it may constitute a possible therapeutic method for CRPC.
References
Siegel R, Naishadham D, Jemal A. Cancer statistics, 2012. CA Cancer J Clin. 2012;62(1):10–29.
Han W, Ding P, Xu M, Wang L, Rui M, Shi S, et al. Identification of eight genes encoding chemokine-like factor superfamily members 1–8 (CKLFSF1–8) by in silico cloning and experimental validation. Genomics. 2003;81(6):609–17.
Su Yu, Lin Yi, Zhang Lianhai, Liu Baocai, Yuan Wanqiong, Mo Xiaoning, et al. CMTM3 inhibits cell migration and invasion and correlates with favorable prognosis in gastric cancer. Cancer Sci. 2014;105(1):26–34.
Li Zesong, Xie Jun, Jianting Wu, Li Wenjie, Nie Liping, Sun Xiaojuan, et al. CMTM3 inhibits human testicular cancer cell growth through inducing cell-cycle arrest and apoptosis. PLoS One. 2014;9(2):e88965.
Wang Y, Li J, Cui Y, Li T, Ng KM, Geng H, et al. CMTM3, located at the critical tumor suppressor locus 16q22.1, is silenced by CpG methylation in carcinomas and inhibits tumor cell growth through inducing apoptosis. Cancer Res. 2009;69:5194–201.
Xie J, Yuan Y, Liu Z, Xiao Y, Zhang X, Qin C, et al. CMTM3 is frequently reduced in clear cell renal cell carcinoma and exhibits tumor suppressor activities. Clin Transl Oncol. 2013;16:402–9.
Di Meo S, Airoldi I, Sorrentino C, Zorzoli A, Esposito S, Di E, et al. Interleukin-30 expression in prostate cancer and its draining lymph nodes correlates with advanced grade and stage. Clin Cancer Res. 2014;20(3):585–94.
Sasaki T, Ryo A, Uemura H, Ishiguro H, Inayama Y, Yamanaka S, et al. An immunohistochemical scoring system of prolyl isomerase Pin1 for predicting relapse of prostate carcinoma after radical prostatectomy. Pathol Res Pract. 2006;202(5):357–64.
Shao L, Cui Y, Li H, Liu Y, Zhao H, Wang Y, et al. CMTM5 exhibits tumor suppressor activities and is frequently silenced by methylation in carcinoma cell lines. Clin Cancer Res. 2007;13(19):5756–62.
Wang Y, Li T, Qiu X, Mo X, Zhang Y, Song Q, et al. CMTM3 can affect the transcription activity of androgen receptor and inhibit the expression level of PSA in LNCaP cells. Biochem Biophys Res Commun. 2008;371:54–8.
Lessel D, Gamulin M, Kulis T, Toliat MR, Grgic M, Friedrich K, et al. Replication of genetic susceptibility loci for testicular germ cell cancer in the Croatian population. Carcinogenesis. 2012;33:1548–52.
Sanchez-Pulido L, Martin-Belmonte F, Valencia A, Alonso MA. MARVEL: a conserved domain involved in membrane apposition events. Trends Biochem Sci. 2002;27(12):599–601.
Livak KJ, Schmittgen TD. Analysis of relative gene expression data using real-time quantitative PCR and the 2(-Delta C (T)) method. Methods. 2001;25:402–8.
Y Xiao, Y Yuan, Y Zhang, J Li,Z Liu, X Zhang et al. CMTM5 is reduced in prostate cancer and inhibits cancer cell growth in vitro and in vivo. Clin Transl Oncol. 2014 [Epub ahead of print].
Sasaki T, Ryo A, Uemura H, Ishiguro H, Inayama Y, Yamanaka S. Animmunohistochemical scoring system of prolyl isomerase Pin1 for predictingrelapse of prostate carcinoma after radical prostatectomy. Pathol Res Pract. 2006;202:357–64.
Chen Y, Zheng L, Liu J, Zhou Z, Cao X, Lv X, et al. Shikonin inhibits prostate cancer cells metastasis by reducing matrix metalloproteinase-2/-9 expression via AKT/mTOR and ROS/ERK1/2 pathways. Int Immunopharmacol. 2014;21(2):447–55.
Carvajal-Carmona LGl, Cazier JB, Jones AM, Howarth K, Broderick P, Pittman A. Fine-mapping of colorectal cancer susceptibility loci at 8q23.3, 16q22.1 and 19q13.11: refinement of association signals and use of in silico analysis to suggest functional variation and unexpected candidate target genes. Hum Mol Genet. 2011;20:2879–88.
Friedl P, Wolf K. Tumour-cell invasion and migration: diversity and escape mechanisms. Nat Rev Cancer. 2003;3:362–74.
Wang Y, Li J, Cui Y, Li T, Ng KM, Geng H, et al. CMTM3, located at the critical tumor suppressor locus 16q22.1, is silenced by CpG methylation in carcinomas and inhibits tumor cell growth through inducing apoptosis. Cancer Res. 2009;69(12):5194–201.
Sridhar SC, Miranti CK. Tetraspanin KAI1/CD82 suppresses invasion by inhibiting integrin-dependent crosstalk with c-Met receptor and Src kinases. Oncogene. 2006;25(16):2367–78.
Sharma S, Kelly TK, Jones PA. Epigenetics in cancer. Carcinogenesis. 2010;31:27–36.
Acknowledgments
This work was supported by the National Natural Science Foundation of China (No. 31171341). We thank Professor Han and Peking University Center for Human Disease Genomics for their generous offer of CMTM3 antibody and adenovirus vector. We also thank the Peking University Department of Pathology for their technology support of immunohistochemistry and staining evaluation.
Conflict of interest
The authors declare no conflict of interest.
Author information
Authors and Affiliations
Corresponding authors
Rights and permissions
About this article

Cite this article
Hu, F., Yuan, W., Wang, X. et al. CMTM3 is reduced in prostate cancer and inhibits migration, invasion and growth of LNCaP cells. Clin Transl Oncol 17, 632–639 (2015). https://doi.org/10.1007/s12094-015-1288-9
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s12094-015-1288-9