
Abstract
Invasion speeds can be calculated from matrix integrodifference equation models that incorporate stage-specific demography and dispersal. These models also permit the calculation of the sensitivity and elasticity of invasion speed to changes in demographic and dispersal parameters. Such calculations have been used to understand the factors determining invasion speed and to explore possible tactics to manage invasive species. In this paper, we extend these calculations to temporally varying environments. We present formulas for the invasion speed and its sensitivity and elasticity in both periodic and stochastic environments. Periodic models can describe seasonal variation within a year, or can be used to study the frequency of occurrence of events (e.g., floods, fires) on interannual time scales. Stochastic models can incorporate variances, covariances, and temporal autocorrelation of parameters. We show that the invasion speed is calculated from a growth rate which is in turn calculated from a periodic or stochastic product of moment-generating function matrices. We present a new formulation of sensitivity analysis, using matrix calculus, that applies equally to constant, periodic, and stochastic environments.
Similar content being viewed by others
Avoid common mistakes on your manuscript.
Introduction
When a population is introduced into a region where it is initially absent, it may spread across the landscape with a characteristic speed. In the idealized case of a homogenous, time-invariant, and infinite landscape, the invasion proceeds as a wave of fixed shape moving at a constant speed (given some assumptions described below). The speed of this invasion wave depends on both demography (i.e., on the rates of survival, development, reproduction, etc.) and on dispersal (i.e., on the probability distribution of distances dispersed by individuals at each stage of their life cycle), and can be calculated from an integrodifference equation model. Because the invasion speed, c *, integrates demography and dispersal into a single index of population spread, it plays a role analogous to that played by the population growth rate (λ or r = logλ) in demographic analysis (Neubert and Caswell 2000). Sensitivity and elasticity analyses of c * provide insight into how demographic and dispersal parameters influence invasion speed, and permit LTRE decomposition of observed differences in invasion speed into contributions from both kinds of parameters (Caswell et al. 2003).
Environments fluctuate, but such fluctuations do not appear in these idealized calculations. In this paper, we derive the invasion speed, and its sensitivity and elasticity, from models for periodically and stochastically varying environments. Periodic models can be used to describe seasonal variation within a year. Seasonality has obvious effects on both demography and dispersal, because in many species breeding biology and dispersal behavior exhibit strong seasonal variation. Periodic models can also be used on interannual time scales to study the effects of the frequency of occurrence of some environmental event (e.g., fire, flood, etc.). Stochastic models extend such calculations to include random components (variance, covariance, and temporal autocorrelation) of environmental fluctuations. Schreiber and Ryan (2010), in an accompanying paper, rightly emphasize the importance of stochastic models to address predictions of increased interannual stochastic variability as a result of climate change.
Neubert et al. (2000) derived expressions for invasion speed in variable environments in the special case of unstructured, scalar populations. Those results, however, do not apply to invasions by age- or stage-structured populations, because of the interaction between the fluctuating environment and the changing population structure. Our approach here, which includes any demographic stage structure, extends the approach of Neubert and Caswell (2000). In that paper, we derived the invasion speed from the dominant eigenvalue of a matrix that includes, in a very particular way, both demography and dispersal. In this paper, we derive invasion speeds in periodic and stochastic environments from periodic and stochastic products of such matrices.
Time-invariant invasion models have by now been applied to many species, in order to explore the properties responsible for invasion speed or to identify targets for potential management strategies (e.g., Miller and Tenhumberg 2010; Smith et al. 2009; Tinker et al. 2008; Jongejans et al. 2008; Bullock et al. 2008; Skarpaas and Shea 2007; Garnier and Lecomte 2006; Vellend et al. 2006; Jacquemyn et al. 2005; Buckley et al. 2005; Le Corff and Horvitz 2005; Bullock and Pywell 2005; Powell et al. 2005; Shea 2004; Neubert and Parker 2004; Caswell et al. 2003; Fagan et al. 2002). Our results will extend these applications to variable environments.
This paper presents four results: the invasion speed in periodic environments, the invasion speed in stochastic environments, the sensitivity analysis in periodic environments, and the sensitivity analysis in stochastic environments. We state these results in “Periodic environments”, “Stochastic environments”, and “Sensitivity and elasticity analysis”. The derivations of these results are an essential part of the paper; for clarity, we have collected these in an Appendix A. We present a simple example in “An example” and conclude with a discussion.
Invasion speed in constant environments
We suppose that the life cycle has been divided into a set of demographically relevant stages. The state of the population is given by a vector n(x, t), whose entries are the densities of the stages at location x and time t. We assume that the landscape is one dimensional.
The demographic rates are incorporated into a density-dependent projection matrix B [n(x, t)]. In the absence of dispersal, the population at location x would grow according to
This system has an equilibrium at n = 0; the matrix of the linear approximation near this equilibrium is \({\bf B}[{\boldsymbol{0}}] \equiv {\bf A}\). This low-density approximation describes population dynamics near the front of the invasion wave, and will be used in calculating the wave speed.
Movement is described by a matrix K(x, y) of dispersal kernels, where k ij (x, y) is the probability density function of the location x of an individual at time t + 1 given that it was at location y at time t, and that it moved from stage j to stage i during the interval. If the environment is spatially homogeneous (as we will assume here), K(x, y) depends only on the distance between x and y, and can be written K(x − y).
Dispersal and demography are combined into the integrodifference equation
where ∘ is the Hadamard, or element-by-element product. The model (2) says that the population at every location x at time t + 1 derives from the growth of the population at all other locations and on the arrival of individuals from those locations, integrated over the entire landscape.
We make the usual biological assumptions that B[n] is non-negative and primitive for all n ≥ 0. We also assume that the density-dependence includes no Allee effects, satisfies
Lui (1989), Neubert and Caswell (2000), and is cooperative; i.e.,
(e.g., Weinberger et al. 2002). Under these conditions, the invasion speed is governed by the low-density leading edge of the invasion wave, where the demography is described by the linearization A , and the invasion is described by the linear integrodifference equation
The wavespeed is calculated from the moment-generating function of the dispersal kernel matrix
In these calculations, we make frequent use of the matrix
This matrix projects, from one time to the next, a wave with an exponential shape proportional to e − sx; hence we call it the wave projection matrix (see Eq. 57).
In terms of the wave projection matrix, the asymptotic invasion speed is given by
where ρ(s) is the dominant eigenvalue of H(s) (Neubert and Caswell 2000). We denote by s * the value of s associated with c *.
Periodic environments
Suppose now that the environment cycles, with a period p, through a set of distinct phases (e.g., seasons). Associated with each phase i is a density-dependent matrix B i [n], a low-density linearization A i , a dispersal kernel matrix K i (x − y), a moment-generating function matrix M i (s), and a wave projection matrix H i (s). The wave projection matrix over a complete environmental cycle, from t to t + p, is the product of the H i ,
(note the order of the matrices, from right to left, in the product). Let ρ per(s) be the dominant eigenvalue of H(s). The invasion speed measured as distance per environmental cycle length (i.e., from t to t + p) is
The average invasion speed, measured as distance per unit time, is
For the derivation of Eq. 10, see “Invasion speed in periodic environments: derivation”.
Stochastic environments
Suppose now that A and/or K are generated by a stochastic environmental process, producing sequences A t , K t , H t , and H t . We assume that the environmental process is stationary and ergodic. A stationary process is one in which, roughly speaking, the statistical properties of the environment do not change over time. More exactly, the joint distribution of the states at any set of times {t, t + δ 1,...,t + δ n } is constant, independent of the starting time t, the intervals δ i , and the number of intervals n. An ergodic process is one that eventually “forgets” the effects of its starting state. This class is wide enough to include some of the most commonly used stochastic ecological models, including independent and identically distributed (iid) environments, finite state Markov chains, and ARIMA processes (Tuljapurkar 1990). We will describe assumptions on A t and K t below.
Let logρ stoch be the stochastic growth rate (more generally, the dominant Lyapunov exponent) of the random matrix product of the H t (s); that is,
where w is an arbitrary non-negative vector. This is the time-averaged growth rate over the distribution generated in a single long realization of the environmental process. The long-term average invasion speed in this stochastic environment is, with probability 1,
For the derivation of the stochastic invasion speed, see “Invasion speed in stochastic environments: derivation”.
Conditions sufficient for the convergence of Eq. 12 are well known in demography (e.g., Cohen 1976; Tuljapurkar and Orzack 1980; Tuljapurkar 1990). In general, it is sufficient that the stochastic process defining the environment be ergodic and that the matrices be non-negative and sufficiently well behaved that any sufficiently long sequence of them forgets the initial state of the population. Since the A t are non-negative and the M t are positive, the H t (s) have the same pattern of zero and positive entries as do the A t . Thus, a sufficient condition for ergodicity of the H t is that the A t are drawn from an ergodic set of matrices; i.e., a set with the property that a product of any large enough number of members of the set will be positive with probability 1. An example of an ergodic set is a set of primitive matrices that share a common incidence matrix.
Sensitivity and elasticity analysis
Invasion speed depends on a set of stage-specific parameters, some related to demography, some to dispersal, and some possibly to both. One of the advantages of calculating invasion speed from a model is that it permits perturbation analysis, which gives the sensitivity and elasticity of c * to changes in the parameters (Neubert and Caswell 2000). The results may suggest targets for management interventions designed to slow the spread of invasive pests, or hasten the recovery of reintroduced species. They also permit LTRE analyses to explain differences in invasion speed in terms of contributions from the vital rates and the dispersal parameters (Caswell et al. 2003).
The invasion speeds in constant environments (Eq. 8), periodic environments (Eq. 10) and stochastic environments (Eq. 13) each depend on a growth rate measure ρ. Sensitivity analysis in the time-invariant case was presented by Neubert and Caswell (2000). Here, we present a new, general, formulation for sensitivity that encompasses the constant, periodic, and stochastic cases, using the matrix calculus developed by Magnus and Neudecker (1985, 1988); see Caswell (2006, 2007, 2008, 2009a, b), Verdy and Caswell (2008), Jenouvrier et al. (2010), Klepac and Caswell (2010) for other ecological applications. In what follows, the derivative of a m×1 vector z with respect to a n×1 vector u is the m×n Jacobian matrix
Derivatives of matrices are calculated by transforming the matrix into a vector using the vec operator, which stacks the columns of the matrix one above the other.
The elasticity of a z with respect to u isFootnote 1
where \(\mathfrak{D}({\bf u})\) is a diagonal matrix with the vector u on the diagonal and zeros elsewhere (produced in Matlab by the command diag(x)).
Suppose that demography, dispersal, or both depend on a vector \({\boldsymbol{\theta}}\) (dimension q ×1) of parameters. These parameters might include matrix elements, lower-level vital rates, or parameters appearing in the dispersal kernel. The vector \({\boldsymbol{\theta}}\) might also include parameters that define the response of demography or dispersal to the state of the environment. The sensitivities of c * to \({\boldsymbol{\theta}}\) are given by the (1 ×q) derivative vector
The details of the calculation depend on the nature of ρ, which in turn depends on whether the environment is constant, periodic, or stochastic. For the derivation of Eq. 16, see “Perturbation analysis: derivations”.
Sensitivity in constant environments
In a constant environment, ρ is the dominant eigenvalue of H(s *) = A ∘ M (s *). Let w and v be the right and left eigenvectors of H(s *), scaled so that \({\bf v}^{\mbox{\tiny \sf T}} {\bf w} =1 \).
The invasion speed c * is given by Eq. 8. Its sensitivity is
where the matrix calculus version (Caswell 2010) of the well-known eigenvalue sensitivity equation (Caswell 2001) is
and
Here, ⊗ denotes the Kronecker product and \({\mathfrak{D} \,}({\bf u})\) is a matrix with u on the diagonal and zeros elsewhere. The elasticities of c * are given by
Sensitivity in periodic environments
The invasion speed c * is given by Eq. 10, where ρ per is the dominant eigenvalue of the periodic product
Again, let w and v be the eigenvectors of H(s *). The parameter vector will generally vary with time; let the value at phase i of the cycle be \({\boldsymbol{\theta}}_i\). Then
where
with
and
The matrices H i and M i and all derivatives are evaluated at s = s * and \({\boldsymbol{\theta}} ={\boldsymbol{\theta}}_i\).
The elasticities of c * are
The sensitivity of the average speed over one cycle of the environment, \(\bar{c}^*\), is
The elasticities of \(\bar{c}^*\) are the same as the elasticities of c *. For derivations, see “Perturbation analysis: derivations”.
Note that environment-specific sensitivities, giving the effect of perturbations applied only in certain phases of the cycle, can be obtained by defining a set \(\mathfrak E\) of environmental states in which the perturbation is applied, and then setting
(Caswell 2005; Aberg et al. 2009).
Sensitivity in stochastic environments
In a stochastic environment, the long-term average invasion speed is given by Eq. 13, where logρ stoch is the familiar stochastic growth rate generated by the random matrix product
The sensitivity of \(\bar{c}^*\) is
The derivative of logρ stoch is calculated using the algorithm of Tuljapurkar (1990), extended to lower-level vital rates (Caswell 2005) and rewritten in terms of matrix calculus (Caswell 2010). For some large T, generate a sequence of matrices \({\bf H}_i(s^*)\) for i = 0,...,T − 1. Pick an arbitrary w(0) with || w(0) || = 1 and generate forward sequences of structure vectors
and 1-step growth rates
Pick an arbitrary v(T), with || v(T)|| = 1 and generate the backward sequence of stochastic reproductive value vectors
Then the derivative of logρ stoch is
where \(d {\mbox{vec} \,} {\bf H}_i/ d{\boldsymbol{\theta}}^{{\mbox{\tiny \sf T}}}\) is given by Eq. 23 and H i and the derivatives are evaluated at s = s * and \({\boldsymbol{\theta}}={\boldsymbol{\theta}}_i\). The elasticity of \(\bar{c}^*\) is
Environment-specific sensitivities can be calculated as in Eq. 27. Note that, regardless of whether the environment is constant, periodic, or stochastic, if the parameters in \({\boldsymbol{\theta}}\) affect only demography, or only dispersal, then terms involving \(d {\mbox{vec} \,} {\bf M}_i/d {\boldsymbol{\theta}}^{{\mbox{\tiny \sf T}}}\) or \(d {\mbox{vec} \,} {\bf A}_i/ d {\boldsymbol{\theta}}^{{\mbox{\tiny \sf T}}}\), respectively, will be zero. The present formulation, however, permits calculating all the sensitivities and elasticities at once, as well as including parameters that affect both demography and dispersal.
An example
Here we present an example, and use it to compare the calculated invasion speeds with the speeds measured in simulation. We also carry out perform sensitivity and elasticity analysis on the invasion speed. We purposely make both demography and dispersal simple.
Model construction
The demographic model is a modified version of the two-stage (juvenile and adult) model of Neubert and Caswell (2000). Adult fertility (ϕ) is a decreasing function of the local density of adults. The other vital rates—juvenile survival (σ 1), maturation (γ), and adult survival (σ 2)—are density-independent. We suppose that environmental variability affects only fertility, so that ϕ is a function of time and the remaining vital rates are constant. The resulting density-dependent projection matrix is
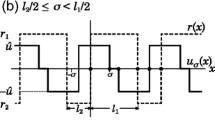
We suppose that dispersal occurs only when juveniles become adults; i.e., in the (2,1) position in K. As a dispersal kernel we use the generalized beta distribution
with |x| ≤ b, b > 0, ν > 0, and where B(a, b) is Euler’s beta function (Whittaker and Watson 1935).
This kernel has a flexible shape (Fig. 1) and has the satisfying property that infinite distance dispersal is impossible. It is symmetric around the origin, and vanishes beyond a finite maximum dispersal distance b. When the shape parameter ν < 1 the kernel is bimodal, with peaks near the maximum dispersal distance, reminiscent of ballistic kernels (Neubert and Parker 2004). When ν > 1 the mode is at the origin; as ν grows the kernel becomes more peaked. The moment-generating function of k 21(x) is the confluent hypergeometric functionFootnote 2

The generalized beta dispersal kernel (Eq. 35) with scale parameter (maximum dispersal distance) b = 1
We suppose that all the dispersal parameters are constant except for the scale parameter (the maximum dispersal distance) b, which varies with time as b(t). The dispersal kernel and moment-generating function matrices are then
where δ(x) is the Dirac delta function.
Invasion speed
As a simple example, we consider an environment that switches between two states, “good” and “bad”, with fertility, ϕ(t), and dispersal distance, b(t), both reduced in a bad year. We set parameters in the two types of years to:
We suppose that, in the periodic environment, good and bad years alternate, and a stochastic environment in which good and bad years occur independently with a probability of 0.5. The invasion speeds, calculated from Eq. 11 in the periodic environment and from Eq. 13 in the stochastic environment, are
where logρ stoch is calculated from Eq. 12 with \(T=\mbox{10,000}\).
As a comparison with these calculated invasion speeds, we simulated both periodic and stochastic environments. In the periodic simulation, the oscillation of the shape of the invasion wave front is apparent in Fig. 2. Fig. 3 shows the location of the wave front as a function of time; the front advances during good years, and retreats during bad years, but regardless of whether it is measured in good years or bad years, it advances at a constant speed. That speed should be \(\bar{c}^*\) as calculated from Eq. 11, and Fig. 4 shows the convergence of the average speed to time t of the simulation to the calculated value of \(\bar{c}^* = 0.0347\). Similarly, Fig. 5 shows the convergence of the stochastic simulation to the calculated value of \(\bar{c}^*=0.0333\).


The location of the juvenile and adult wave fronts in the simulation shown in Fig. 2, in a periodic environment


a The location of the invasion front in 20 realizations of a stochastic environment. b The average speed of the wave to time t for 20 realizations. The model is the stochastic version of that shown in Fig. 2, but with good and bad years occurring independently with a probability of 0.5. The average speed (solid lines) converges to the prediction (dashed line) based on the linearization given by formula (13) Parameter values are given in Eq. 39
Sensitivity analysis
We define the parameter vector as
where the first four parameters affect only demography (in A) and the last two affect only dispersal [in M(s)]. The derivatives \(d {\mbox{vec} \,} {\bf A} / d {\boldsymbol{\theta}}^{{\mbox{\tiny \sf T}}}\) and \(d {\mbox{vec} \,} {\bf M}/d {\boldsymbol{\theta}}^{{\mbox{\tiny \sf T}}}\) required in Eqs. 19 and 23 are given in Appendix B.
The sensitivities and elasticities of \(\bar{c}^*\) are shown in Fig. 6. In this case (and it need not be so in general), the patterns for the periodic environment and the stochastic environment are qualitatively similar. The sensitivities and elasticities to all the demographic parameters and to the scale of dispersal (b) are all positive, while the sensitivity to the shape parameter (ν) is negative.

Sensitivity and elasticity of the invasion wave speed \(\bar{c}^*\) to changes in each of the demographic and dispersal parameters; parameter values given in Eq. 39. In the periodic case, the environment alternates between the two states. In the stochastic case, the two environmental states occur independently with probability 1/2
In this hypothetical example, management tactics that reduce juvenile survival (σ 1) would be an attractive option, although proportional reductions in ϕ would have almost as great an effect.
Discussion
The invasion speed of a structured population is obtained from a product (periodic or stochastic, as the case may be) of demography-dispersal matrices. Because matrix multiplication is not commutative, this implies that the order in which the environmental states appear must be taken into account. Just as in demographic models, one cannot apply results for scalar populations (Neubert et al. 2000) to the eigenvalues of the matrices \({\bf H}_i(s^*)\). Situations exist where invasion is impossible in every environmental state (i.e., \(\rho \left[ {\bf H}_i(s^*) \right] <1\) for all i) and yet it is possible in a periodic environment that varies among the states (i.e., \(\rho \left[ {\bf H}_p(s^*) \cdots {\bf H}_1(s^*) \right]>1\)). Or vice-versa (e.g., Caswell 2001, Example 13.2).
Matrix integrodifference equation models are easily parameterized from commonly collected kinds of demographic and dispersal data (Neubert and Caswell 2000). This extends to the periodic and stochastic versions, which require only population projection matrices and/or dispersal kernels expressed as functions of the environmental state. As in the example in “An example”, this may require as few as two projection matrices and/or dispersal kernels. Stochastic analyses have become common in demographic studies; we expect that they will also do so in studies of invasion. For example, studies of how the statistics of the environment (e.g., the frequency of good or bad years) affects invasion speeds are only beginning (Schreiber and Ryan 2010).
Our findings complete a set of results on invasion speed and its perturbation analysis for scalar and structured populations in constant, periodic, and stochastic environments (Kot et al. 1996; Neubert and Caswell 2000; Neubert et al. 2000). Table 1 summarizes the calculation of invasion speed in each of these six cases. In each case, the invasion speed is the minimum, over the shape parameter s, of a growth rate calculated from a demographic model and the moment-generating function of a dispersal kernel.
These calculations share a related set of assumptions. They assume that the environment is spatially homogeneous and infinite in extent. They assume that the linear conjecture (van den Bosch et al. 1990; Weinberger et al. 2002) holds, so that the invasion speed is determined by the linearization of the demographic model around zero. This requires the assumption that no Allee effects operate (for an analysis of such effects, see Wang et al. 2002). The calculations in Table 1 also assume that an asymptotic wave speed exists, which requires that the dispersal kernels are exponentially bounded, so that the moment-generating functions exist. If this assumption is violated, the invasion may result in a continually accelerating invasion wave. Such accelerating invasions, however, cannot occur if there is a finite maximum dispersal distance, no matter how large.
Convergence and transients
Like the population growth rate, the invasion wave speed is an asymptotic property. There is every reason to expect short-term transients to be important in the early stages of an invasion. Comparing time-invariant, periodic, and stochastic models (Table 1), we see that convergence to the asymptotic invasion speed requires as many as three distinct kinds of convergence, each of which requires the system to “forget” one kind of initial condition:
-
1.
Convergence of the environment to its stationary distribution of states. The rate of this convergence is a property of the stochastic model for the environment. A constant environment is always in its stationary state.
-
2.
Convergence of the stage structure to its stable form, or, in a stochastic environment, convergence of the probability distribution of the stage structure to an invariant distribution. The rate of this convergence depends on the structure of the life cycle underlying the demographic matrices B i . A scalar population is always at its stable structure.
-
3.
Convergence of the spatial distribution of the population to its travelling wave form. The rate of this convergence depends on the dispersal kernel causing the population to forget its initial form.
Each “dimension” introduces another rate of convergence. A scalar population in a constant environment has only a single dimension of convergence (convergence of the spatial structure to the travelling wave shape). A structured population in a constant environment must converge in stage structure and spatial structure. A scalar population in a variable environment, must converge in a spatial structure, and the environment must converge to its stationary form. A structured population in a stochastic environment must converge in all three dimensions. To our knowledge, the operation of the factors that determine these rates of convergence, and the transient dynamics during convergence, have not yet been studied.
Notes
There seems to be no standard notation for elasticity corresponding to that used for derivatives. The notation, \(\varepsilon {\bf z} / \varepsilon {\bf u}^{\mbox{\tiny \sf T}}\), that we use here is adapted from Samuelson (1947).
The function 0 F 1 can be obtained in software packages supporting a limited set of special functions using the identity \(_{0}F_{1}(a,z) = \Gamma (a)\, (-z)^{(1 - a)/2}\, J_{a - 1} \left( 2 \sqrt{-z} \right)\), and noting that 0 F 1(a, 0) = 1, where J a (z) is the Bessel function of order a of the first kind (Watson 1944).
References
Aberg P, Svensson CJ, Caswell H, Pavia H (2009) Environment-specific elasticity and sensitivity analysis of the stochastic growth rate. Ecol Model 220:605–610
Buckley YM, Brockerhoff E, Langer L, Ledgard N, North H, Rees M (2005) Slowing down a pine invasion despite uncertainty in demography and dispersal. J Appl Ecol 42:1020–1030
Bullock JM, Pywell RF (2005) Rhinanthus: a tool for restoring diverse grassland? Folia Geobot 40:273–288
Bullock JM, Pywell RF, Coulson-Phillips SJ (2008) Managing plant population spread: prediction and analysis using a simple model. Ecol Appl 18:945–953
Caswell H (2001) Matrix population models: construction, analysis, and interpretation, 2nd edn. Sinauer Associates, Sunderland, Massachusetts, USA
Caswell H (2005) Sensitivity analysis of the stochastic growth rate: three extensions. Aust N Z J Stat 47:75–85
Caswell H (2006) Applications of Markov chains in demography. In Langville AN, Stewart WJ (eds) MAM2006: Markov anniversary meeting. Boson Books, Raleigh, North Carolina, USA, pp 319–334
Caswell H (2007) Sensitivity analysis of transient population dynamics. Ecol Lett 10:1–15
Caswell H (2008) Perturbation analysis of nonlinear matrix population models. Demographic Research 18:59–116
Caswell H (2009a) Sensitivity and elasticity of density-dependent population models. J Differ Equ Appl 15:349–369
Caswell H (2009b) Stage, age, and individual stochasticity in demography. Oikos 118:1763–1782
Caswell H (2010) Reproductive value, the stable stage distribution, and the sensitivity of the population growth rate to changes in vital rates. Demogr Res 23:531–548. doi:10.4054/DemRes.2010.23.19
Caswell H, Lensink R, Neubert MG (2003) Demography and dispersal: life table response experiments for invasion speed. Ecology 84:1968–1978
Cohen JE (1976) Ergodicity of age structure in populations with Markovian vital rates, I: countable states. J Am Stat Assoc 71:335–339
Fagan WF, Lewis MA, Neubert MG, van den Driessche P (2002) Invasion theory and biological control. Ecol Lett 5:148–157
Furstenberg H, Kesten H (1960) Products of random matrices. Ann Math Stat 31:457–469
Garnier A, Lecomte J (2006) Using a spatial and stage-structured invasion model to assess the spread of feral populations of transgenic oilseed rape. Ecol Model 194:141–149
Grimmett GR, Stirzaker DR (1992) Probability and random processes. Oxford University Press, Oxford
Jacquemyn H, Brys R, Neubert MG (2005) Fire increases invasive spread of Molinia caerulea mainly through changes in demographic parameters. Ecol Appl 15:2097–2108
Jenouvrier S, Caswell H, Barbraud C, Weimerskirch H (2010) Mating behavior, population growth and the operational sex ratio: a periodic two-sex model approach. Am Nat 175:739–752
Jongejans E, Shea K, Skarpaas O, Kelly D, Sheppard AW, Woodburn TL (2008) Dispersal and demography contributions to population spread of Carduus nutans in its native and invaded ranges. J Ecol 96:687–697
Klepac P, Caswell H (2010) The stage-structured epidemic: a multi-state matrix population model approach. Theoretical Ecology. doi:10.1007/s12080-010-0079-8
Kot M, Lewis MA, van den Driessche P (1996) Dispersal data and the spread of invading organisms. Ecology 77:2027–2042
Le Corff J, Horvitz CC (2005) Population growth versus population spread of an ant-dispersed neotropical herb with a mixed reproductive strategy. Ecol Model 188:41–51
Lui R (1989) Biological growth and spread modeled by systems of recursions. I. Mathematical theory. Math Biosci 93:269–295
Magnus JR, Neudecker H (1985) Matrix differential calculus with applications to simple, Hadamard, and Kronecker products. J Math Psychol 29:474–492
Magnus JR, Neudecker H (1988) Matrix differential calculus with applications in statistics and econometrics. Wiley, New York, New York, USA
Miller TEX, Tenhumberg B (2010) Contributions of demography and dispersal parameters to the spatial spread of a stage-structured insect invasion. Ecol Appl 20:620–633
Neubert MG, Caswell H (2000) Demography and dispersal: calculation and sensitivity analysis of invasion speed for structured populations. Ecology 81:1613–1628
Neubert MG, Parker IM (2004) Projecting rates of spread for invasive species. Risk Anal 24:817–831
Neubert MG, Kot M, Lewis MA (2000) Invasion speed in fluctuating environments. Proc R Soc Lond B 267:1603–1610
Powell JA, Slapnicar I, van der Werf W (2005) Epidemic spread of a lesion-forming plant pathogen: analysis of a mechanistic model with infinite age structure. Linear Algebra Appl 398:117–140
Samuelson PA (1947) Foundations of economic analysis. Harvard University Press, Cambridge, MA, USA
Schreiber SJ, Ryan ME (2010) Invasion speeds for structured populations in fluctuating environments. Theoretical Ecology (in press)
Shea K (2004) Models for improving the targeting and implementation of biological control of weeds. Weed Technol 18:1578–1581
Skarpaas O, Shea K (2007) Dispersal patterns, dispersal mechanisms, and invasion wave speeds for invasive thistles. Am Nat 170:421–430
Smith CA, Giladi I, Lee YS (2009) A reanalysis of competing hypotheses for the spread of the California sea otter. Ecology 90:2503–2512
Tinker MT, Doak DF, Estes JA (2008) Using demography and movement behavior to predict range expansion of the southern sea otter. Ecol Appl 18:1781–1794
Tuljapurkar S (1990) Population dynamics in variable environments. Springer, New York, USA
Tuljapurkar SD, Orzack SH (1980) Population dynamics in variable environments I. Long-run growth rates and extinction. Theor Popul Biol 18:314–342
van den Bosch F, Metz JAJ, Diekmann O (1990) The velocity of spatial population expansion. J Math Biol 28:529–565
Vellend M, Knight TM, Drake JM (2006) Antagonistic effects of seed dispersal and herbivory on plant migration. Ecol Lett 9:316–323
Verdy A, Caswell H (2008) Sensitivity analysis of reactive ecological dynamics. Bull Math Biol 70:1634–1659
Wang M-H, Kot M, Neubert MG (2002) Integrodifference equations, Allee effects, and invasions. J Math Biol 44:150–168
Watson GN (1944) A Treatise on the Theory of Bessel Functions, 2nd edn. Cambridge University Press, Cambridge, UK
Weinberger HF, Lewis MA, Li B (2002) Analysis of linear determinacy for spread in cooperative models. J Math Biol 45:183–218
Whittaker ET, Watson GN (1935) A Course of Modern Analysis, 4th edn. Cambridge University Press. Cambridge, UK
Wolfram Research (2010). Available at: http://functions.wolfram.com/07.17.20.0004.01 and http://functions.wolfram.com/07.17.20.0001.01. Accessed 23 May 2010
Acknowledgements
This research has been supported by NSF Grants DEB-0235692, DEB-0816514, DEB-0343820, OCE-0530830, and OCE-1031256. Two reviewers made helpful comments, and we thank S. Schreiber and M. Ryan for sharing their manuscript with us.
Author information
Authors and Affiliations
Corresponding author
Appendices
Appendix A: Derivations
Invasion speed in periodic environments: derivation
We assume an environmental cycle with p phases (e.g., 12 months within a year). Associated with phase i are matrices B i and K i , for i = 1,...,p, and we suppose we have numbered the phases so that we project the population starting in phase 1. Then, the periodic integrodifference equation is
Over a full cycle, from t to t + p, the model is time-invariant, and the invasion speed is determined by its linearization around n = 0:
Each step in this cycle is a convolution of the matrix K ∘ A with the population distribution n(y). To simplify notation, let * denote convolution; i.e., write
Then the linearized model can be written
where we define \(\bigvee\) to denote the multiple convolution operator
Note the order of the convolution, written from right to left, corresponding to the order in which the phases appear in the cycle.
The model (45) is simply the constant environment invasion model (5), projecting the population from t to t + p, using the p-fold convolution
as the dispersal and demography matrix.
To compute the wavespeed over an entire cycle of the environment, we need the moment-generating function of the p-fold convolution matrix (Eq. 47), i.e., we need
It is well known that the moment-generating function of a convolution is the product of the moment-generating functions of the distributions in the convolution (e.g., Grimmett and Stirzaker 1992, Section 5.7), so
The wave speed per one period of the environment (i.e., from t to t + p) is obtained, as in any constant environment, as
where ρ(s) is the dominant eigenvalue of H(s) (Neubert and Caswell 2000). The average wave speed per unit time is then
Invasion speed in stochastic environments: derivation
We suppose that the matrices B t [n] and K t are generated by a stationary, ergodic stochastic environmental process, and that the matrices A t form an ergodic set. An ergodic set is a set with the property that a product of any large enough number of members of the set will be positive with probability 1. An example is a set of primitive matrices with a common incidence matrix.
The integrodifference equation for the low-density linearization is
We begin by considering a single realization of the stochastic environment, and thus a single-specific sequence of A t and K t . We calculate the average invasion speed for this realization by finding an expression for the location, at time t, of the “front” of the wave, defined as the point at which the population density exceeds some (low) threshold value.
We consider an initial condition exponentially distributed in space,
where w is an arbitrary vector and s > 0. Then Eq. 52 says that
Define a new variable u = x − y, we rewrite Eq. 54 as
Iterating this process leads to the solution to Eq. 52 for this particular initial condition,
where the product of the H i must be taken in the order
Define the total population size as N = || n ||, and choose a critical population level N cr that defines the location of the front of the invasion. Then the location X(t) of the invasion at time t is the value of x where N(x, t) = N cr. We have
Setting the two expressions for N cr equal, we have
which implies that
Between t = 0 and t = T the location of the wave front has advanced from X(0) to X(T), and hence the average invasion speed between t = 0 and t = T is
The long-term average invasion speed is obtained by taking the limit as T → ∞:
where ρ stoch now denotes the stochastic growth rate (more generally, the dominant Lyapunov exponent) of the stochastic matrix process defined by the H i (s).
Equation 64 was derived for a single realization of the stochastic environment. However, stochastic ergodic theory (Furstenberg and Kesten 1960; Cohen 1976; Tuljapurkar and Orzack 1980; Tuljapurkar 1990) says that, under our assumptions on the environmental process and the matrices H i , (Eq. 64) converges with probability 1 to the fixed value logρ stoch, and thus that (Eq. 65) gives the long-term average invasion speed of every realization except for a set of measure zero.
Initial conditions
The solution (Eq. 58) was derived by assuming an initial condition with an exponential spatial distribution. However, following Neubert and Caswell (2000), we can use this solution to obtain a least upper bound on the invasion wave speed for an arbitrary initial condition with compact support.
Let n(x, 0) be an initial population with compact support. For any s > 0 there exists a scalar β and a vector w with || w || = 1 such that
By the assumption (4) on density dependence, this implies that the solution originating from n(x, 0) can never exceed that originating from β w e − sx. Thus
and
which, by (Eq. 61), implies
Thus, for any s, the speed of the invasion resulting from n(x, 0) with compact support is no greater than that resulting from an exponential initial condition with shape s. So, picking the s that gives the slowest exponential invasion gives a least upper bound to the speed of the invasion of the arbitrary initial condition n(x, 0). Thus the arbitrary initial condition cannot, in the long run, spread faster than \(c^* = \min_s c(s)\). It also cannot spread slower than c * if it maintains its exponential shape.
In the absence of either proof or numerical examples to the contrary, we assume that in fact the non-exponential initial condition eventually converges to the speed given by the slowest exponential wave.
Perturbation analysis: derivations
We begin with the expression for the invasion speed for an arbitrary s, written as a function of a parameter vector \({\boldsymbol{\theta}}\),
where the parameters in \({\boldsymbol{\theta}}\) may affect demography, dispersal, or both. The nature of the growth rate ρ depends on whether the environment is constant, periodic, or stochastic.
Differentiating both sides of Eq. 71 gives
But logρ = sc and d logρ = (1/ρ) d ρ. Making these substitutions into Eq. 72 gives
Substituting Eq. 73 into Eq. 72 gives
The realized invasion speed c * is the minimum over s of c(s). Thus, at s = s * it follows dc/ds = 0, and hence that
and thus the derivative of c * with respect to \({\boldsymbol{\theta}}\) is
The sensitivity results in “Sensitivity and elasticity analysis” are obtained by using the appropriate results for the derivative of logρ, depending on whether the environment is constant, periodic, or stochastic.
Derivatives of H
The wave projection matrix is given by
The derivatives of H, in Eqs. 19 and 23, are needed for sensitivity analysis. Differentiating both sides yields
Next, apply the vec operator, noting that
to get
The chain rule gives the expression (19) for \(d {\mbox{vec} \,} {\bf H} / d {\boldsymbol{\theta}}^{{\mbox{\tiny \sf T}}}\).
Appendix B: Derivatives for the numerical example
The calculation of sensitivities in Eqs. 19 and 23 requires the derivatives of A and M(s) with respect to the parameter vector \({\boldsymbol{\theta}}\). The derivatives of the vec of the matrix A, with respect to each of the parameters, are arranged as columns of the matrix
The vertical line separates derivatives with respect to the demographic parameters (θ 1–θ 4) and those with respect to dispersal parameters (θ 5–θ 6).
The derivatives of the moment-generating function of the dispersal kernel (Eq. 36) with respect to b and ν are
where 0 F 1 is the confluent hypergeometric function, Γ(·) is the gamma function, and Ψ(·) is the polygamma function (Wolfram Research 2010).
The resulting matrix of derivatives is
Rights and permissions
About this article
Cite this article
Caswell, H., Neubert, M.G. & Hunter, C.M. Demography and dispersal: invasion speeds and sensitivity analysis in periodic and stochastic environments. Theor Ecol 4, 407–421 (2011). https://doi.org/10.1007/s12080-010-0091-z
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s12080-010-0091-z