
Abstract
Antitumour necrosis factor-alpha \((\hbox {TNF-}\upalpha )\) therapy is used as a clinical intervention for rheumatoid arthritis (RA) but differences exist in response to the treatment which makes the candidature of the screening of \(\hbox {TNF-}\upalpha \) alteration(s) at genetic and expression levels an important agenda prior to treatment. This study aims to determine the associative role of \(\hbox {TNF-}\upalpha \) –308G/A polymorphism and differential expression of \(\hbox {TNF-}\upalpha \) in the pathogenesis of RA. A case–control study where a total of 126 RA patients were enrolled based on ACR-EULAR (2010) criteria, along with 160 community matched age and sex controls over a period of three years. The differential expression level of \(\hbox {TNF-}\upalpha \) mRNA and protein level was studied and \(\hbox {TNF-}\upalpha \) –308G/A polymorphism was screened by T-ARMS PCR assay. All statistical analysis was performed using SPSS software. mRNA expression level of \(\hbox {TNF-}\upalpha \) was upregulated in RA cases (avg. \(15.85\pm 9.52\) fold) compared to control. \(\hbox {TNF-}\upalpha \) protein level was found to be higher in RA cases (\(28.62\pm 7.17\) pg/mL) compared to control (\(23.14\pm 6.91\) pg/mL). \(\hbox {TNF-}\upalpha \) –308 variant GA genotype was higher in RA (46.03%) than in control (25%). The presence of \(\hbox {TNF-}\upalpha \) –308 variant A allele was associated with increased risk of RA susceptibility (odds ratio \((\hbox {OR})=2.559\) at 95% confidence interval (CI), \(P < 0.001\)) but not severity (\(\hbox {OR}=1.617\) at 95% CI, \(P=0.571\)). The presence of –308 variant genotype was associated with a higher \(\hbox {TNF-}\upalpha \) mRNA and protein expression. The presence of \(\hbox {TNF-}\upalpha \) –308A allele is associated with increased risk of RA susceptibility and differential \(\hbox {TNF-}\upalpha \) expression, and has prognostic significance. Association of higher \(\hbox {TNF-}\upalpha \) pro-inflammatory cytokine levels with northeast Indian patients makes them suitable subjects for \(\hbox {anti-TNF-}\upalpha \) therapy.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
Rheumatoid arthritis (RA) is a chronic synovial inflammatory arthropathy and has been underlined as a multisystem autoimmune disease (Nemec et al. 2008). RA is known to affect individuals of any age group (Alamanos and Drosos 2005) and women are reported to be approximately three times more susceptible to the disease than men. Chronic inflammation of RA is also associated with cardiovascular disease which is found to be responsible for the mortality of RA patients (Dhawan and Quyyumi 2008). RA is a global issue and affects \(\sim \)0.5–1% population in developed countries (Bax et al. 2011). The last decade has seen an explosion in data on RA genetics, with multiple genetic linkage and association studies have confirmed the genetic basis of RA in different populations throughout the world (MacGregor et al. 2000; Raychaudhuri 2010) but existing data are still equivocal and inconclusive, which underlines the requirement of further scientific studies and evidences on the aetiology of RA pathogenesis.
Emerging genetic data with an emphasis on the immunogenetic links using molecular and immunological tools have been a centre stage of RA aetiological studies universally, with several immunologically important signalling pathways being screened or targeted for RA treatment (Messemaker et al. 2015) with variable results. Cytokines play an essential role in articular destruction in association with inflammation. Tumour necrosis factor-alpha \((\hbox {TNF-}\upalpha )\) is a potent pleiotropic proinflammatory cytokine, produced mostly by activated monocytes, macrophages and T-lymphocytes and promotes inflammatory responses that are important in the pathogenesis of RA. A high concentration of \(\hbox {TNF-}\upalpha \) is found in the synovial fluid of RA patients and is associated with erosions of the bone (Hassan et al. 2011). Positive data have been accumulated from in vitro, in vivo and patient-based clinical investigation studies. Gheita et al. (2015) have rationalized the use of \(\hbox {anti-TNF-}\upalpha \) antagonists such as infliximab (IFX), etanercept (ETN), adalimumab (ADA), golimumab (GOLI) and certolizumab pegol (CZP) for the treatment of RA which have definitely correlated with clinically relevant improvement in a subpopulation of RA cases. But certain patients exhibit a lack of response to \(\hbox {anti-TNF-}\upalpha \) therapy and clinical administration of \(\hbox {anti-TNF-}\upalpha \) antagonists may have adverse effects on these patients and predispose these cases to infections of multiple aetiologies which are unwanted. Therefore, studies on the importance and specificity of differential \(\hbox {TNF-}\upalpha \) expression in RA susceptibility and severity in specific geographical population may prove vital for efficient use of available \(\hbox {anti-TNF-}\upalpha \) antagonists in specific population and individuals and massive reduction in the possible adverse side effects.
The TNF gene is located on the short arm of chromosome 6 (6p21), and the promoter region of TNF gene is an extremely polymorphic locus which is enriched with a number of single nucleotide polymorphisms (SNPs). A transitional polymorphism from G to A nucleotide at position 308 (also known as \(TNF\hbox {-}\alpha \) –308 or rs1800629) of the promoter region of \(TNF\hbox {-}\alpha \) gene is associated with increased levels of \(\hbox {TNF-}\upalpha \) expression (Hajeer and Hutchinson 2000; Elahi et al. 2009). The rare A allele of \(TNF\hbox {-}\alpha \) gene is associated with worse clinical outcomes in different diseases like sepsis and multiple trauma in comparison to the G allele (Menges et al. 2008; Teuffel et al. 2010). Recent studies have provided evidences that \(\hbox {TNF-}\upalpha \) polymorphism plays an important role in RA disease although there are many discrepancies for different populations (Danis et al. 1995; Gambhir et al. 2010) with no scientific studies or data available from ethnically distinct northeast Indian population. Based on the existing literature and lacunae, presented here is a novel study evaluating the importance of profile changes in \(\hbox {TNF-}\upalpha \) expression and genotype as risk factors of RA susceptibility and severity in northeast Indian patients.
Materials and methods
Patient enrolment and stratification
A total of 126 RA patients who were clinically diagnosed with RA were enrolled in the present study with all clinical details. The patients were recruited from the Rheumatology OPD of Gauhati Medical College and Hospital, Guwahati, India based on ACR and EULAR (2010) criteria. The RA cases were further clinically stratified as mild RA cases (ACR and EULAR (2010) criteria score: 5–7; \(n\,{=}\,14\)) and severe RA cases (ACR and EULAR (2010) criteria score: 8–10; \(n=112\)). A group of 160 age and sex matched unrelated blood donors were randomly enrolled as healthy controls. The healthy control individuals were free of any clinical evidence of autoimmune diseases or familial history of RA. All RA patients and healthy controls were from the northeast region of India and informed written consent was obtained from both the patients and the healthy donors prior to enrolment. The study was approved by the institutional ethics committee of the participating institutes and is in accordance with the ethical standards laid down in the 1964 Declaration of Helsinki.
Differential mRNA and protein expression analysis
Blood samples were collected from the healthy control individuals and patients in EDTA and clot vials. The serum was separated and these samples were used for enzyme-linked immunosorbent assay based study for the cytokine (Merck, EZHTNFA). Total RNA was isolated from the samples using the standard Trizol method, quantified using nanodrop spectrophotometry and was converted to c-DNA using commercially available c-DNA synthesis kit (Applied Biosystems, Foster City, USA) following the manufacturer’s protocol. The c-DNA was used to determine the level of \(\hbox {TNF-}\upalpha \) by qPCR (7500 Fast Real-Time PCR System, Applied Biosystems) using SyBr green chemistry and \(\beta \)-actin gene as the internal normalization control. The difference in expression level of \(\hbox {TNF-}\upalpha \) was evaluated on the basis of fold change calculated based on \(2^{-\Delta \Delta \mathrm{Ct}}\) formulae.
\(\hbox {TNF-}\alpha \) –308 genotyping analysis
Genomic DNA was extracted from collected whole blood using the standard phenol–chloroform method. The quality of extracted genomic DNA was analysed by nanodrop spectrophotometry. The \(TNF\hbox {-}\alpha \) –308 G/A locus was amplified in a total volume of \(20~\mu \hbox {L}\) using allele-specific PCR (amplification refractory mutation system (ARMS)-PCR) based assays (table 1) and was analysed by agarose gel electrophoresis. The interpretation of the \(\hbox {TNF-}\upalpha \) genotype was based on the amplification sizes as stated in table 1.
Statistical analysis
All statistical analyses were performed by the standard methods using SPSS V13.0 software (SPSS, Chicago, USA). The significance was described as Pearson P value. Associations between \(TNF\hbox {-}\alpha \) –308 G/A polymorphism and RA susceptibility, and RA severity were calculated as odds ratio (OR) with 95% confidence intervals (CIs); a two-tailed \(P <0.05\) was considered statistically significant.
Results
Demographical and clinical profile
The present study was based on the northeast Indian population, with clinically proven RA patients being enrolled under the supervision of a registered medical practitioner. The details of the enrolled patients are shown in table 2.
Differential mRNA and protein expression analysis in RA cases
The differential \(\hbox {TNF-}\upalpha \) mRNA expression data evaluated by real-time PCR showed an increased expression of \(\hbox {TNF-}\upalpha \) in RA cases (avg. \(15.85\pm 9.52\) fold) in comparison with control. However, the difference in expression pattern was not observed between mild and severe RA cases (\(P=0.970\)) thus clearly indicating that transcriptional activation of \(\hbox {TNF-}\upalpha \) has a major role in disease susceptibility (figure 1). The typical range of serum \(\hbox {TNF-}\upalpha \) is in the range of 5–27 pg/mL. The \(\hbox {TNF-}\upalpha \) protein level data show a clear upregulation in RA cases (\(28.62\pm 7.17\) pg/mL) as compared to controls (\(23.14\pm 6.91\) pg/mL) (\(P=0.428\)), suggesting a role in disease susceptibility. The serum \(\hbox {TNF-}\upalpha \) levels were nonsignificantly higher in severe RA cases compared to mild RA cases (\(P=0.879\)) (figure 1), therefore underlining the importance of differential \(\hbox {TNF-}\upalpha \) expression in RA pathogenesis.

Top panel shows the real-time PCR based amplification for \(\hbox {TNF-}\upalpha \) gene c-DNA and difference in fold change in mild and severe RA cases compared to control. Bottom panel presents the difference in protein expression in control and RA cases, as well as between the mild and severe RA cases.
\(\hbox {TNF-}\alpha \) –308 promoter SNP distribution analysis
A nucleotide change at position –308 (G to A) in the promoter region of \(TNF\hbox {-}\alpha \) gene is associated with increased levels of \(\hbox {TNF-}\upalpha \) expression, and was thus evaluated by allele-specific PCR (ARMS-PCR) followed by agarose gel electrophoresis based analysis (figure 2). The difference in genotype distribution in the enrolled subjects is represented in tables 3 and 4.
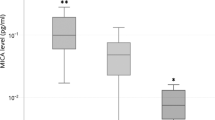
The presence of either wild-type GG genotype or heterozygous AG genotype was observed in the enrolled subjects. The distribution of heterozygous AG genotype was significantly higher in RA cases compared to control (\(P<0.001\)), and was associated with increased risk of RA susceptibility (\(\hbox {OR}=2.559\) (1.551–4.222) at 95% CI, \(P <0.001\)). The presence of heterozygous variant AG genotype was also higher in severe RA cases compared to mild cases (\(P =0.413\)), and was found to be associated with increased risk to RA severity (\(\hbox {OR}=1.617\) (0.510–5.129) at 95% CI, \(P =0.571\)). When the difference in expression of \(\hbox {TNF-}\upalpha \) was analysed based on \(\hbox {TNF-}\upalpha \) –308 genotype distribution, the AG variant genotype was associated with significantly increased \(\hbox {TNF-}\upalpha \) mRNA (\(P =0.002\)) and protein expression (\(P =0.009\)) (figure 3); which underlines the prognostic significance of \(\hbox {TNF-}\upalpha \) –308 genotype in RA pathogenesis.

Representative panel of agarose gel electrophoresis results for \(\hbox {TNF-}\upalpha \) (–308) SNP showing the presence of different amplified band patterns indicating the presence of either wild type (198 bp) or variant (homozygous (139 bp) or heterozygous (198+139 bp)) genotypes. The outer amplified product of 294 bp was common for both the genotypes.
Discussion
RA is a complex multifactorial systemic disease, which involves chronic inflammation induced by interplay of various cytokines that contribute significantly in the development and progression of the disease (Mosaad et al. 2011). An imbalance between proinflammatory and anti-inflammatory cytokine activities favours the induction of auto-immunity and chronic inflammation ultimately leading to joint deformity, crippling and loss of function. \(\hbox {TNF-}\upalpha \), being a proinflammatory cytokine, is considered to have a major role in disease induction. Over-production of \(\hbox {TNF-}\upalpha \) contributes to increased reactive oxygen species (ROS) release in RA patients, leading to tissue damage associated with inflammation (Hassan et al. 2011). \(\hbox {TNF-}\upalpha \) has been demonstrated to regulate multiple cytokines, vascular endothelial growth factor (VEGF) production, recruitment of immune and inflammatory cells into joints, angiogenesis and reduction of blood levels of matrix metalloproteinases which are known to be associated with RA pathogenesis (Feldmann and Maini 2001). Immunotherapy has markedly improved treatment outcomes in RA. \(\hbox {TNF-}\upalpha \) antagonists, such as infliximab (IFX), etanercept (ETN), adalimumab (ADA), golimumab (GOLI) and certolizumab pegol (CZP) have been widely used for the treatment of RA (Ma and Xu 2010). Although \(\hbox {TNF-}\upalpha \) antagonist based therapy has been reported to impart marked improvements in the disease conditions in RA patients, but adverse or lack of response has been documented in a subpopulation of RA patients which is a critical scenario. Therefore determining the candidature of the RA patients for \(\hbox {anti-TNF-}\upalpha \) therapy is a definite prerequisite. Molecular diagnostics based on differential expression of \(\hbox {TNF-}\upalpha \) and screening of genetic alterations in the \(\hbox {TNF-}\upalpha \) gene associated with differential \(\hbox {TNF-}\upalpha \) expression may be a useful prognostic tool for stratification of RA patients for candidature for \(\hbox {anti-TNF}\upalpha \) therapy in a given geographical population. A transitional polymorphism from G to A nucleotide at position 308 of the promoter region of \(TNF\hbox {-}\alpha \) gene is associated with increased levels of \(\hbox {TNF-}\upalpha \) expression. The variant A allele is associated with higher levels of TNF transcript than the G allele which is considered to be the ancestral allele (Hajeer and Hutchinson 2000; Elahi et al. 2009). Therefore, the present study involving RA cases from northeast Indian population was undertaken to screen the role of genetic alteration in \(\hbox {TNF-}\upalpha \) gene and associated differential \(\hbox {TNF-}\upalpha \) expression in the pathogenesis of RA in northeast Indian subjects, and predict the efficacy of \(\hbox {anti-TNF-}\upalpha \) drugs in the given population.

Box-plot analysis showing significantly increased \(\hbox {TNF-}\upalpha \) expression in subjects with variant genotype.
The ethnically distinct northeast Indian population has been found to be predisposed to RA with a prevalence rate of 0.2–0.4% (BJD India Copcord Survey 2001). A lacuna exists in terms of the role of difference in distribution of genetic alterations in \(\hbox {TNF-}\upalpha \) gene and linked differential \(\hbox {TNF-}\upalpha \) expression in RA pathogenesis from northeast India. Explorations of differential expression of \(\hbox {TNF-}\upalpha \) were suggestive of increased fold change at the mRNA level and serum protein levels being associated with RA susceptibility compared to controls in the northeast Indian population. A study with the north Indian population (Jahid et al. 2017) showed a positive correlation between circulating \(\hbox {TNF-}\upalpha \) levels and mRNA expression in RA patients as high as 4.5 times more than in healthy individuals. Moreover, \(\hbox {TNF-}\upalpha \) level was also reported to be higher in the serum of RA patients as compared to healthy controls from other groups also (Robak et al. 1998).
The differential \(\hbox {TNF-}\upalpha \) expression was comparative between mild and severe RA cases, but interestingly difference in \(\hbox {TNF-}\upalpha \) expression at both mRNA and serum protein levels was observed in a sub-population of cases belonging to both the mild and severe groups as well as in healthy subjects control group, which indicated that probably the underlying \(\hbox {TNF-}\upalpha \) genotype plays some influential role in expression status of the gene, and in turn disease pathogenesis. The –308 polymorphism is one of the most investigated variant in association with RA in different populations (Cvetkovic et al. 2002). However, the association of –308 polymorphism with susceptibility and severity to RA has been conflicting and equivocally documented (Brinkman et al. 1995; Wilson et al. 1997). Therefore, studying other relevant polymorphisms at the promoter region like -238 (G/A) or at the first intronic region of \(TNF\hbox {-}\alpha \) +489 (G/A) as reported in the available literature (van Krugten et al. 1999; Murdaca et al. 2014) becomes an interesting and important agenda to validate the fact whether these polymorphisms may influence \(\hbox {TNF-}\upalpha \) profile changes and maybe associated with RA pathogenesis through futuristic studies. These futuristic studies on different TNF polymorphisms may crucially help for stratifying patients predisposed to RA susceptibility and severity as well as in selecting the specific anti-cytokine drug therapy for selective group of patients, thereby underlining their prognostic and biomarker significance.
The data from this study strongly underlined the prognostic significance of ‘A’ allele in RA cases, as the heterozygous AG genotype was associated with increased risk of RA susceptibility (\(\hbox {OR}=2.559\), \(P<0.001\)) and severity (\(\hbox {OR}=1.617\), \(P=0.571\)), and more importantly was significantly associated with increased \(\hbox {TNF-}\upalpha \) mRNA (\(P=0.002\)) and protein expression (\(P=0.009\)). These data are in agreement and concurrence with studies involving Spanish population (Edith et al. 2008), Czech population (Brinkman et al. 1995; Nemec et al. 2008) and Egyptian population (Gheita et al. 2015). But the present data are contrary to the findings on Han Chinese population (Li et al. 2015), Southeast Asians (Yen et al. 2003) and in Finnish populations (Maury et al. 2003) which reported a negative association of \(\hbox {G}\rightarrow \hbox {A}\) polymorphism of \(\hbox {TNF-}\upalpha \) gene in RA pathogenesis; as well as existing data from other global pockets (Brinkman et al. 1995; Wilson et al. 1997; Lacki et al. 2000) or from other parts of India (Gambhir et al. 2010) who did not findany association of \(\hbox {TNF-}\upalpha \) –308 G/A polymorphism with RA pathogenesis. The aforementioned differences with respect to global or national data maybe due to ethnic differences among the populations under study, and therefore underlines the significance of studying the \(\hbox {TNF-}\upalpha \) genotype and differential expression for stratifying RA cases which may eventually benefit from the existing \(\hbox {anti-TNF}\upalpha \) therapies independently or in combinatorial therapies. This may definitely minimize the complications and side-effects of the standard disease modifying anti-rheumatoid drugs (DMARDs) still under use in any given population.
In conclusion, within the limitations, this first study from ethnically distinct northeast Indian population is suggestive of the role of \(\hbox {TNF-}\upalpha \) genotype and linked differential expression in RA disease pathogenesis. This study underlines the prognostic significance of \(\hbox {TNF-}\upalpha \) –308A allele in RA cases for clinical stratification and selecting drug regime. Since higher \(\hbox {TNF-}\upalpha \) proinflammatory cytokine levels were found to be associated with RA in northeast Indian patients, this study could be a possible solution to choose this population as suitable subjects for \(\hbox {anti-TNF-}\upalpha \) therapy and its benefits.
References
Alamanos Y. and Drosos A. A. 2005 Epidemiology of adult rheumatoid arthritis. Autoimmun. Rev. 4, 130–136.
Bax M., Jurgen van H., Huizinga Tom W. J. and Toes Rene E. M. 2011 Genetics of rheumatoid arthritis: what have we learned? Immunogenetics 63, 459–466.
Brinkman B. M., Zuijdeest D., Kaijzel E. L., Breedveld F. C. and Verweij C. L. 1995 Relevance of the tumor necrosis factor alpha (TNF alpha)-308 promoter polymorphism in TNF alpha gene regulation. J. Inflamm. 46, 32–41.
Cvetkovic J. T., Wallberg-Jonsson S., Stegmayr B., Rantapaa-Dahlqvist S. and Lefvert A. K. 2002 Susceptibility for and clinical manifestations of rheumatoid arthritis are associated with polymorphisms of the TNF-alpha, IL-1beta, and IL-1Ra genes. J. Rheumatol. 29, 212–219.
Danis V. A., Millington M., Hyland V., Lawford R., Huang Q. and Grennan D. 1995 Increased frequency of the uncommon allele of tumour necrosis factor alpha gene polymorphism in rheumatoid arthritis and systemic lupus erythematosus. Dis. Markers 12, 127–133.
Dhawan S. S. and Quyyumi A. A. 2008 Rheumatoid arthritis and cardiovascular disease. Curr. Atheroscler. Rep. 10, 128–133.
Edith O.-R., Vázquez-Del M. M., Luz R.-Q. S., Navarro-Hernández R. E., Rangel-Villalobos H., Martínez-Bonilla G. et al. 2008 Tumor necrosis factor \(\upalpha \)-308 and -238 polymorphisms in rheumatoid arthritis. Association with messenger RNA expression and sTNF-\(\upalpha \). J. Invest. Med. 56, 937–943.
Elahi M. M, Asotra K, Matata B. M. and Mastana S. S. 2009 Tumor necrosis factor alpha-308 gene locus promoter polymorphism: an analysis of association with health and disease. BBA Mol. Basis Dis. 1792, 163–172.
Feldmann M. and Maini R. N. 2001 Anti-TNF alpha therapy of rheumatoid arthritis: what have we learned. Annu. Rev. Immunol. 19, 163–196.
Gambhir D., Lawrence A., Aggarwal A., Misra R., Mandal S. K. and Naik S. 2010 Association of tumor necrosis factor alpha and IL-10 promoter polymorphisms with rheumatoid arthritis in North Indian population. Rheumatol. Int. 30, 1211–1217.
Gheita T. A., Azkalany Ghada S., Gaber W. and Mohey A. 2015 Clinical significance of serum TNF\(\upalpha \) and -308 G/A promoter polymorphism in rheumatoid arthritis. Egypt. Rheumatol. 37, 49–54.
Hajeer A. H and Hutchinson I. V. 2000 TNF-alpha gene polymorphism: clinical and biological implications. Microsc. Res. Tech. 50, 216–228.
Hassan S. Z., Gheita T. A., Kenawy S. A., Fahim A. T., El-Sorougy I. M. and Abdou M. S. 2011 Oxidative stress in systemic lupus erythematosus and rheumatoid arthritis patients: relationship to disease manifestations and activity. Int. J. Rheum. Dis. 14, 325–331.
Jahid M., Rehan-Ul-Haq, Jha P. K., Chawla D., Avasthi R. and Ahmed R. S. 2017 Tumor necrosis factor-\(\upalpha \)-308 polymorphism in North Indian rheumatoid arthritis patients and association with mRNA and serum \(\text{ TNF- }\upalpha \). Clin. Rheumatol. 36, 2209–2216.
Krugten van M. V., Huizinga T. W. J., Kaijzel E. L., Zanelli E., Drossaers-Bakker E. W., van de Linde P. et al. 1999 Association of the TNF +489 polymorphism with susceptibility and radiographic damage in rheumatoid arthritis. Genes Immun. 1, 91–96.
Lacki J. K., Moser R., Korczowska I., Mackiewicz S. and Muller W. 2000 \(\text{ TNF- }\upalpha \) gene polymorphism does not affect the clinical and radiological outcome of rheumatoid arthritis. Rheumatol. Int. 4, 137–140.
Li F., Xie X., Chen J., Gao J. and Lu F. 2015 Association of \(\text{ TNF- }\upalpha \) gene polymorphisms with the risk of rheumatoid arthritis in Han Chinese population from Hunan. J. Central South Univ. 40, 945–954.
Ma X. and Xu S. 2010 TNF inhibitor therapy for rheumatoid arthritis. Biomed. Rep. 1, 177–184.
MacGregor A. J., Snieder H., Rigby A. S., Koskenvuo M., Kaprio J., Aho K. et al. 2000 Characterizing the quantitative genetic contribution to rheumatoid arthritis using data from twins. Arthritis Rheum. 43, 30–37.
Maury C. P., Liljeström M., Laiho K., Tiitinen S., Kaarela K. and Hurme M. 2003 Tumor necrosis factor alpha, its soluble receptor I, and -308 gene promoter polymorphism in patients with rheumatoid arthritis with or without amyloidosis: implications for the pathogenesis of nephropathy and anemia of chronic disease in reactive amyloidosis. Arthritis Rheum. 48, 3068–3076.
Menges T., König I. R., Hossain H., Little S., Tchatalbachev S. and Thierer F. et al. 2008 Sepsis syndrome and death in trauma patients are associated with variation in the gene encoding tumor necrosis factor. Crit. Care Med. 36, 1456–1462.
Messemaker T. C., Huizinga T. W. and Kurreeman F. 2015 Immunogenetics of rheumatoid arthritis: understanding functional implications. J. Autoimmun. 64, 74–81.
Mosaad Y. M., Abdelsalam A. and El-bassiony S. R. 2011 Association of tumour necrosis factor-alpha-308 G/A promoter polymorphism with susceptibility and disease profile of rheumatoid arthritis. Int. J. Immunogenet. 38, 427–433.
Murdaca G., Gulli R., Spano F., Lantieri F., Burlando M., Parodi A. et al. 2014 \(\text{ TNF- }\upalpha \) gene polymorphisms: association with disease susceptibility and response to \(\text{ anti-TNF- }\upalpha \) treatment in psoriatic arthritis. J. Invest. Dermatol. 134, 2503–2509.
Nemec P., Pavkova-Goldbergova M., Stouracova M., Vasku A., Soucek M. and Gatterova J. 2008 Polymorphism in the tumor necrosis factor-alpha gene promoter is associated with severity of rheumatoid arthritis in the Czech population. Clin. Rheumatol. 27, 59–65.
Raychaudhuri S. 2010 Recent advances in the genetics of rheumatoid arthritis. Curr. Opin. Rheumatol. 22, 109–118.
Robak T., Gladalska A. and Stepień H. 1998 The tumour necrosis factor family of receptors/ligands in the serum of patients with rheumatoid arthritis. Eur. Cytokine Netw. 9, 145–154.
Teuffel O., Ethier M. C., Beyene J. and Sung L. 2010 Association between tumor necrosis factor-alpha promoter -308 A/G polymorphism and susceptibility to sepsis and sepsis mortality: a systematic review and meta-analysis. Crit. Care Med. 38, 276–282.
Wilson A. G., Symons J. A., McDowell T. L., McDevitt H. O. and Duff G. W. 1997 Effects of a polymorphism in the human tumor necrosis factor alpha promoter on transcriptional activation. Proc. Natl. Acad. Sci. USA 94, 3195–3199.
Yen J. H., Chen C. J., Tsai W. C., Lin C. H., Ou T. T., Hu C. J. et al. 2003 Manganese superoxide dismutase and cytochrome P450 1A1 genes polymorphisms in rheumatoid arthritis in Taiwan. Hum. Immunol. 64, 366–373.
Acknowledgements
The authors would like to acknowledge the staff of Gauhati Medical College and Hospital, Guwahati, Assam, India for their support in sample collection during the study.
Author information
Authors and Affiliations
Corresponding author
Additional information
Corresponding editor: Indrajit Nanda
Rights and permissions
About this article

Cite this article
Das, S., Baruah, C., Saikia, A.K. et al. Genetic and expression changes in \(\hbox {TNF-}\upalpha \) as a risk factor for rheumatoid arthritis pathogenesis in northeast India. J Genet 98, 3 (2019). https://doi.org/10.1007/s12041-018-1054-1
Received:
Revised:
Accepted:
Published:
DOI: https://doi.org/10.1007/s12041-018-1054-1