
Abstract
Down syndrome (DS) is one of the most common genetic diseases. Patients with DS display growth delay and intellectual disabilities and develop Alzheimer’s disease (AD) neuropathology after middle age, including neuritic plaques and neurofibrillary tangles. Beta-site amyloid β precursor protein (APP) cleaving enzyme 1 (BACE1), essential for Aβ production and neuritic plaque formation, is elevated in DS patients. However, its homolog, β-site APP cleaving enzyme 2 (BACE2), functions as θ-secretase and plays a differential role in plaque formation. In this study, by using Two-dimensional sodium dodecyl sulfate polyacrylamide gel electrophoresis (2D SDS-PAGE) and LC-MS/MS proteomic profiling analysis, we found that the SET oncogene protein (SET) expression was associated with BACE1 but not BACE2. SET protein was increased in BACE1 overexpressing cells and was markedly reduced in the BACE1 knockout mice. We found that the overexpression of BACE1 or SET significantly inhibited cell proliferation. Moreover, knockdown of SET in BACE1 overexpression cells significantly rescued BACE1-induced cell growth suppression. Furthermore, both BACE1 and SET protein levels were increased in Down syndrome patients. It suggests that BACE1 overexpression-induced SET upregulation may contribute to growth delay and cognitive impairment in DS patients. Our work provides a new insight that BACE1 overexpression not only promotes neuritic plaque formation but may also potentiate neurodegeneration mediated by SET elevation in Alzheimer-associated dementia in DS.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
Down syndrome (DS) is one of the most common genetic diseases. Most of the patients carry an extra copy of chromosome 21 (Trisomy-21 syndrome). It affects one in 800 new births [1]. Patients with DS develop characteristic Alzheimer’s disease (AD) neuropathology after middle age, including amyloid plaques, neurofibrillary tangles, and neuronal loss [2, 3]. The mechanism underlying AD pathogenesis in DS remains elusive. Studies have shown that several genes on chromosome 21, such as amyloid β precursor protein (APP) and regulator of calcineurin 1 (RCAN1) contribute to the overproduction of amyloid β protein (Aβ), the key component of amyloid plaques, and neuronal loss detected in the brain of AD patients and implicated in AD pathogenesis in DS patients [4-8].
Beta-site APP cleaving enzyme 1 (BACE1) plays a critical role in AD pathogenesis. BACE1 is the β-secretase in vivo for Aβ production [9-12]. BACE1 cleaves APP at two β-secretase sites, the Asp1 site to generate C99 fragment and the Glu11 site to generate C89 [13, 14]. Subsequent cleavage of C99 and C89 by γ-secretase results in full-length Aβ and truncated Aβ generation, respectively. Cleavage of APP at the Asp1β site by BACE1 is required for generating Aβ. However, the Aβ Glu11 is the predominant β-secretase site of BACE1 to process APP under normal conditions [14]. Aβ production and deposition initiate the cascade of neuronal cell stress and neuronal cell death, and it is believed to be the driving force for AD development. Our lab reported that BACE1 expression and maturation are upregulated in DS patients and increased APP processing by BACE1 contributes to AD development in DS [7].
BACE1 is essential for Aβ production in AD pathogenesis, and suppression of BACE1 expression or activity significantly reduces APP processing and Aβ generation in vivo and in vitro [15-20]. Knockout of BACE1 rescues the memory deficit present in AD mouse model [21]. Furthermore, inhibition of GSK3β-mediated BACE1 expression reduces Alzheimer-associated phenotypes [20]. These studies suggest that specific inhibition of BACE1 enzymatic activity is a direct and effective way to reduce Aβ production and to prevent the development of AD. However, studies have shown that inhibition of BACE1 could result in many unwanted negative biological consequences including hypomyelination, retinal pathology, impaired movement coordination by affecting the formation and maturation of the muscle spindle; and schizophrenic behaviors in BACE1-deficient mice [22-26]. Nevertheless, BACE1 modulators that reduce Aβ generation without significantly affecting neuregulin and other substrates could be valid candidates for AD drug development.
Beta-site APP cleaving enzyme 2 (BACE2) is a homolog of BACE1. BACE2 gene is located on chromosome 21q22.3, encodings for a 518 amino acid single transmembrane domain protein [27-30]. BACE2 protein degradation is mediated by the macroautophagy-lysosome pathway [31]. BACE2 shares 75 % homologous amino acids sequences with BACE1 [32]. However, expression of BACE2 and BACE1 are regulated distinctly at transcription and translation levels, and BACE2 is not a β-secretase but functions as a novel θ-secretase, mainly processing APP at the θ site between Aβ-Phe+19 and -Phe+20, resulting in inhibition of Aβ generation [6, 32]. These studies suggested that BACE2 and BACE1 could have distinct downstream targets and modulates different signaling pathways in pathological conditions. Nevertheless, both of them can serve as valid therapeutic targets for AD drug development.
SET nuclear oncogene (SET), also known as template-activating factor-I (TAF-1), is a multifunctional nuclear phosphor-protein. It was first discovered in 1992 as part of a fusion protein with nucleoporin Nup214 (CAN) in a patient with acute undifferentiated leukemia [33]. The SET gene is located on chromosome 9q34, and its transcription is controlled by two different promoters [34], resulting in expression of two distinct isoforms of protein designated as SET/TAF-1α (290 amino acids) and SET/TAF-1β (277 amino acids) [35]. The two isoforms only differ in the first 30 amino acid sequence in the N-terminal [35].
SET is widely expressed in a variety of tissues in human [36] and mouse [37] and is involved in many biological functions. SET is a potent inhibitor of the protein phosphatase 2A (PP2A) [38]. Recombinant human SET exhibits strong and specific inhibitory effect on the phosphatase activity of PP2A in vitro [39], and the N-terminal region appears essential for its inhibitory activity [40]. Set/TAF-1β is also involved in forming the complex of endogenous inhibitor of histone acetyltransferases (INHAT), which binds to histone, blocks histone acetylation, and inhibits gene transcription [41]. Moreover, SET is actively involved in cell death mediated by both caspase-cleaved APP fragment and SET mislocalization from the nuclear to the cytoplasm [42, 43]. Recently, studies showed that SET potentiates Tau hyperphosphorylation for neurofibrillary tangle formation, a neuropathological hallmark of AD [44-47].
In this study, by analyzing the protein expression profiles using BACE1 and BACE2 overexpression cell models by two-dimensional gel electrophoresis, we found that the expression of SET, a potent phosphatase inhibitor, was significantly associated with BACE1 expression. Overexpression of BACE1 significantly upregulated the level of SET. The increase in SET level was associated with cell growth inhibition. Moreover, the protein level of SET was significantly higher in the brain tissue of DS patients compared to the controls. Since SET is implicated in AD pathogenesis by regulating Aβ production and tau accumulation, the upregulated SET expression in DS patients could play a critical role in the development of AD among these patients.
Materials and Methods
Cell Culture and Protein Extraction
The cell lines 20E2, 2 EB2, and 4 EB2 were analyzed in this study. 20E2 is a human embryonic kidney (HEK) cell line which stably overexpresses Swedish APP695. 2 EB2 cells stably overexpress Swedish APP695 and mycHis-tagged BACE1. 4 EB2 cells stably overexpress Swedish APP695 and mycHis-tagged BACE2. 293B2 and 4B25 cells are HEK cell line stably overexpressing human BACE1 and BACE2, respectively. The stable cell lines were maintained in complete Dulbecco’s modified Eagle’s medium (DMEM) containing antibiotics neomycin and/or zeocin at 37 °C in an incubator containing 5 % CO2. Harvested cell pellets were lysed in radioimmunoprecipitation assay (RIPA)-deoxycholate (DOC) buffer supplemented with protease inhibitor cocktail (Roche). The cell lysate was quantified using Bio-Rad DC Protein Assay Kit (Bio-Rad).
Plasmid Constructs and Generation of SET Stable Cell Lines
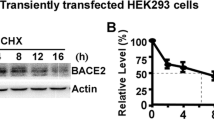
Plasmid pzBACE1-mycHis was used to express human BACE1 protein [48]. To construct SET expression plasmid, RNA was extracted from SH-SY5Y cells. Human SET cDNA was amplified by RT-PCR using following primers 5′-ccgaagcttgccaccatgtcggcgccggcggccaa and 5′aagtctagagtcatcttctccttcatcct. The PCR fragments were cloned into pcDNA4-mycHis vector (Invitrogen) at HindIII and XbaI sites to generate SET expression plasmid termed pzSET-mycHis. The plasmids were confirmed by restriction enzyme digestion and sequencing. To generate SET stably transfected cell lines in HEK293 and N2A cells, plasmid pzSET-mycHis was transiently transfected into these two cell lines and selected with Zeocin. SM3 and NSM were the two clones with stable SET overexpression in HEK293 and in N2A cells, respectively. All the stable cell lines were maintained with 50 mg/L of Zeocin.
Two-Dimensional Sodium Dodecyl Sulfate Polyacrylamide Gel Electrophoresis (2D SDS-PAGE) and Sypro Ruby Staining.
The 2D SDS-PAGE was performed to compare the protein composition between BACE1 and BACE2 overexpressing cell lines. Briefly, cell lysate of 2 EB2 and 4 EB2 were first cleaned up using ReadyPre 2-D Cleanup Kit (Bio-Rad). The purified protein pellets were resuspended in 2D-rehydration buffer 1 (Bio-Rad). The protein concentration was measured using fluorescent-based EZQ protein quantification kit (Invitrogen). A total of 150 μg of protein in 135 μl 2D rehydration buffer was loaded onto 17 cm immobilized pH 3–10 gradient (IPG) strips with linear pH gradient and rehydrated overnight. Isoelectric focusing electrophoresis (IEF) was performed at 250 V for 30 min and 4,000–10,000 V h at 20 °C using Protean IEF cell (Bio-Rad). Separation in the second dimension was performed on 12 % glycine gel using the MiniProtean II Electrophoresis system (Bio-Rad). The SDS-PAGE gel was then stained with Sypro Ruby (Invitrogen) overnight and visualized and documented using GelDoc-It imaging system. Three replicates were performed independently. Dots consistently showing different staining intensity between cell lines were excised manually and subject to liquid chromatography (LC)-mass spectrometry (MS)/MS analysis.
Immunoblotting and Immunoprecipitation
Brain tissues of DS patients were obtained from University of Maryland Brain and Tissue Bank for Developmental Disorders (Baltimore, MD, USA). The average ages of 12 controls and 15 DS patients were 34.8 ± 38.1 years and 26.2 ± 27.3 years, respectively. The causes of death in control cases were not related to brain diseases. BACE1 knockout mice were purchased from JAX lab. Brain tissue or cells were lysed in RIPA-DOC lysis buffer (1 % Triton X100, 1 % sodium deoxycholate, 0.1–4 % SDS, 0.15 M NaCl, 0.05 M Tris-HCl, pH 7.2) supplemented with protease inhibitors (Complete, Boehringer Mannheim). The lysate were separated by SDS-PAGE and transferred to a PVDF membrane. Following blocking of the membranes was incubated in primary and IRDye infrared secondary antibody. Finally, the blots were scanned and analyzed using LI-COR imaging system. Quantification was based on at least three independent experiments. β-actin was used as an internal control for normalization. The following primary antibodies were used: anti-SET (Cell Signaling Technology), anti-RCAN1 (DCT3) [4], anti-BACE1 (208) [49] and anti-myc (9E10). To investigate the interaction between SET and BACE1 and BACE2, immunoprecipitation was performed using Pierce crosslink immunoprecipitation kit (Thermo Scientific). The immunoprecipitates were separated on 12 % Tris-Glycine gel and detected using antibodies for co-immunoprecipitation.
Cell Growth and BrdU Proliferation Assay
To monitor the growth rate of SET overexpression stable cell line, 2.5 × 105 cells were seeded on 35-mm dishes, and the cell number was counted and recorded after 3 days. The cells were continuously seeded and counted every 3 to 4 days. For the BrdU assay, the cells were incubated with 10 μM BrdU for 6 h. After incubation, the cells were washed with PBS and fixed with 4 % paraformaldehyde for 20 min at room temperature. Two molar HCl was applied for 30 min at 37 °C and followed by neutralization for 30 min with 0.1 M sodium borate. The cells were permeabilized in 0.2 % Triton solution for 15 min at room temperature. Fixed cells were blocked with goat serum and then incubated at room temperature for 2 h with goat anti-BrdU antibody (Sigma). Following the incubation, cells were washed with PBS and incubated with Cy3-conjugated anti-goat IgG (Thermo Scientific) for 1 h. After washing with PBS, cells were stained with 4,6-diamidino-2-phenylindole (DAPI) (Sigma). Cell images were recorded by fluorescent microscope (Axiovert200, Carl Zeiss Inc.).
Statistical Analysis
Student’s t test and ANOVA were performed. Values of P < 0.05 were considered significant.
Results
Upregulated Expression of SET in BACE1 Overexpression Cells
2D SDS-PAGE analysis was performed to examine the difference in protein composition between the Swedish mutant APP and BACE1 overexpressing cell line 2 EB2 and the Swedish mutant APP and BACE2 overexpressing cell line 4 EB2. A total of three independent experiments were performed. One dot (marked by arrow in Fig. 1a) in the acidic area in 2 EB2 cells consistently showed stronger staining intensity compared to that in the 4 EB2 cells. A subsequent LC-MS/MS analysis and Mascot search of the excised dot revealed that the protein SET was the top hit among the several identified proteins (Fig. 1b). We next validated the finding by Western analysis (Fig. 1c). The result confirmed that SET was significantly upregulated in 2 EB2 cells compared to the non-BACE1 overexpressing control cell 20E2 (146.93 ± 11.03 % relative to control, p < 0.05) (Fig. 1d). No significant change in the level of SET expression was observed in 4 EB2 cells.

SET expression was upregulated in BACE1 overexpression cells. a Cell lysates from 2 EB2 and 4 EB2 cells were separated by 2D SDS-PAGE and stained with Sypro Ruby. The spots indicated with an arrow were excised for MS analysis; b LC-MS/MS analysis of the dot in a revealed that four peptides of SET were hit; the sequences of the 4-hit peptide were numbered with their corresponding position in SET; c The expression level of SET in 20E2 cells, 2 EB2 cells, and 4 EB2 cells were analyzed using Western blotting. SET was detected by SET N-terminal antibody. β-actin was served as loading control; d SET expression were quantified and normalized to β-actin. The values represent mean ± SEM; *p < 0.05 by Student’s t test, n = 3
Next, we examined the level of SET in another three cell lines 293B2, 4B25, and HEK293. 293B2 is a HEK293 cell line stably expressing BACE1, and 4B25 is a HEK293 cell line stably overexpressing BACE2 [6]. The level of SET expression was markedly increased in 293B2 cells compared to control cell line HEK293 (145.71 ± 6.57 % relative to control, P < 0.05) (Fig. 2a, b). SET expression was not significantly changed in 4B25 cells (Fig. 2a, b). To examine whether the increase in SET protein level was due to upregulated transcription, the mRNA level of SET was assayed. No significant difference was observed between HEK293, 293B2, and 4B25 cell lines (Fig. 2c, d). Moreover, there was no detectable interaction between BACE1 and SET by co-immunoprecipitation (data not shown). These results clearly showed that overexpression of BACE1 but not BACE2 upregulates SET protein level.

SET expression in the BACE1 or BACE2 stably-expressing cells. a The cell lysates of HEK293 cells, 2 EB2 cells, and 4B25 cells were subjected to Western analysis. SET was detected by SET N-terminal antibody. β-actin served as loading control. SET expression was quantified and normalized to β-actin; b quantitative analysis of the SET protein levels. Compared to HEK 293 cells, the protein level of SET in 293B2 cells was increased, and there were no significant changes observed in 4B25. The values represent mean ± SEM. *p < 0.05 by Student’s t test, n = 3; c semiquantitative RT-PCR was performed to measure mRNA level of SET and β-actinin HEK293 cells, 293B2 cells, and 4B25 cells. β-actin served as an internal control; the samples were analyzed on 1 % agarose gel; d quantification of SET mRNA. Compared to the parental cell HEK293, there was no significant difference in the mRNA level of SET in 293B2 cells and 4B25 cells. The values represent mean ± SEM and were analyzed by Student’s t test, n = 3
SET Expression Was Reduced in the Brain of BACE1 Knockout Mice
We have shown SET upregulation in BACE1 overexpressing cells lines. To examine whether inhibiting BACE1 expression could reduce SET expression, the protein level of SET in the brain of BACE1 knockout mice was measured by Western blot analysis. Disruption of BACE1 gene in the BACE1 knockout mice was confirmed by PCR genotyping, and the lack of BACE1 protein expression was further confirmed by Western blot. Compared to the wild type mice and the BACE1 overexpressing 293B2 cells, BACE1 protein was not detected in the brain of BACE1 knockout mice (Fig. 3a). Compared to the wildtype mice, SET expression was dramatically reduced in the brain of BACE1 knockout mice (70.64 ± 8.21 % relative to control, p < 0.01) (Fig. 3b, c). It is indicated that disruption of BACE1 expression resulted in reduced SET expression in vivo.

SET expression was reduced in the brain of BACE1 knockout mice. a Western blot analysis was performed to examine BACE1 level in the brain tissue of wild-type mouse (WT), the BACE1 overexpressing cells (293B2), and the brain tissue of BACE1 knockout mouse (KO). BACE1 was detected by BACE1 antibody 208. Compared to the wild-type mouse and cells with BACE1 overexpression, there was no detectable BACE1 protein in the brain tissue of BACE1 knockout mouse; b SET protein level in the brain tissue of wild-type mice (WT) and BACE1 knockout mice (KO) was assayed by Western blotting. SET was detected by SET N-terminal antibody. β-actin served as loading control; c Quantification of SET protein level in the mouse brain tissues. SET expression was significantly reduced in BACE1 knockout mice. The values represent mean ± SEM and were analyzed by Student’s t test. **p < 0.01, n = 5 in each group
Overexpression of BACE1 Inhibited Cell Proliferation Mediated by SET Upregulation
To assess the effect of BACE1 and SET on cell proliferation, the numbers of cell population doublings between BACE1/SET overexpression cell line and their parental cell line were examined. After 12 days of culture, the growth rate of the BACE1-overexpressing 293B2 cells was 67.40 % of their parental control cells HEK293, with an 8.17 ± 0.09 population doublings (PDs) versus 12.13 ± 0.07 PDs (P < 0.01) (Fig. 4a). To study SET’s effect, a SET stable cell line SM3 was generated by stably-transfected pzSET-mycHis plasmid into HEK293 cells. SM3 cells robustly expressed the mycHis-tagged SET protein, as was detected by both SET-NT antibody and anti-myc antibody 9E10 (Fig. 4b). SM3 cells exhibited 24.13 % decrease of growth rate compared to the parental cell line HEK293, 9.17 ± 0.11 PDs versus 12.13 ± 0.07 PDs at day 12, respectively (P < 0.01) (Fig. 4c). The rate of BrdU uptake between SM3 cells and control HEK293 cells was measured to further examine the effect of SET on cell proliferation. After 6 h incubation, HEK293 had significantly more cells labeled with BrdU compared to SM3 cells (48.26 ± 3.60 vs 31.67 ± 1.79 %, p < 0.01) (Fig. 4d, e).

SET overexpression inhibited cell proliferation. a Growth curve of HEK293 cells and 293B2 cells. The growth rate of 293B2 cells was 67.40 % of HEK293 cells. **p < 0.01 by ANOVA, n = 3; b generation of SM3 stable cell line overexpressing SET-mycHis protein. Cell lysates from HEK293 and SM3 were subjected to Western analysis. Exogenous mycHis-tagged SET protein and endogenous SET protein were detected with SET N-terminal antibody (SET-NT), and exogenous mycHis-tagged SET was also detected by anti-myc antibody 9E10; c Growth curve of HEK293 cells and SM3 cells. The growth rate of SM3 cells was 75.59 % of HEK293 cells at day 12. **p < 0.01 by ANOVA, n = 3; d BrdU cell proliferation assay of HEK293 and SM3 cells. BrdU incorporation was detected by BrdU antibody followed by cy3-conjugated anti-goat IgG (red). Nuclei were stained with DAPI (blue). e The percentage of BrdU-positive cells in SM3 cells was significantly lower than that in HEK293 cells. **p < 0.01 by Student’s t test, n = 4; f Western analysis of N2A cells stably expressing SET. Myc-tagged SET was only detected in NSM cells by 9E10, and endogenous SET was detected by the SET-NT antibody in both N2A and NSM cells. g Growth curve of N2A cells and NSM cells. The growth rate of NSM cells was 78.18 % of N2A cells at day 16. **p < 0.01 by ANOVA, n = 3. h Western blot analysis of 293B2 cells transfected with scramble and SET siRNA. The expression of SET was marked reduced at day 6, while its expression was gradually recovered at day 9. i Growth curve of 293B2 cells transfected with scramble and SET siRNA. The growth rate of 293B2 cells transfected with SET siRNA was significantly increased compared with scramble siRNA transfected 293B2 cells at day 9. **p < 0.01 by ANOVA, n = 3
Since SET is mainly expressed in the brain and its effect could be related to Alzheimer pathogenesis, we further determined its growth-inhibiting effect on neuronal cells. We generated a stable cell line NSM which constitutively overexpressed SET in parental neuroblastoma N2A cells (Fig. 4f). NSM cells showed a marked growth rate reduction, 6.20 ± 0.16 PDs versus 7.94 ± 0.35 PDs at day 6 (P < 0.01) (Fig. 4g). Taken together, these data clearly demonstrate that SET overexpression inhibited cell proliferation in both HEK293 cells and neuronal N2A cells.
To determine whether BACE1-induced inhibition of cell proliferation is mediated by SET, SET knockdown experiments were performed in 293B2 cells by using SET siRNA. Six days after SET siRNA transfection, the expression of SET was markedly reduced compared with cells transfected with scramble siRNA, while its expression was gradually recovered at day 9 (Fig. 4h). Meanwhile, the cell proliferation rate of 293B2 cells transfected with scramble siRNA and SET siRNA was monitored. We found that SET knockdown significantly rescued BACE1-induced growth suppression, 7.89 ± 0.08 versus 7.33 ± 0.07 PDs at day 9 (p < 0.01). It highly suggests that BACE1-induced cell growth suppression is mediated by SET upregulation.
BACE1 and SET Were Upregulated in the Brain of DS Patients
BACE1 plays an important role in AD pathogenesis. In BACE1 overexpression cell model, we have observed that SET was significantly upregulated. We then examined the expression level of SET and BACE1 in the tissue of frontal cortex in a total of 15 DS patients and 12 controls by western blotting (Fig. 5a). SET and BACE1 protein levels were significantly increased in DS patients by 276.29 ± 36.50 % (p < 0.01 relative to control) (Fig. 5b) and 309.90 ± 23.60 % (p < 0.01 relative to control) (Fig. 5c), respectively.

BACE1 and SET were upregulated in the brain of DS patients. a Brain lysates from Down syndrome patients and controls were resolved on 10 % SDS-PAGE and followed by Western blot. SET was detected by SET-NT antibody. BACE1 was detected by BACE1 antibody 208. β-actin was served as loading control; b quantification of SET protein level. The level of SET in the brain tissue of DS patients (N = 15) was markedly increased compared to the control (N = 12) **p < 0.01 by student t test; c quantification of BACE1 protein level. The level of BACE1 in DS patients was also significantly upregulated; **p < 0.01 by Student’s t test
Discussion
AD is the most common neurodegenerative disorder leading to dementia. Patients with DS usually develop Alzheimer-type dementia at the middle age. The incidence rate can reach 60–75 % when they are over 60 years old [50-52]. The mechanism by which DS patients have high incidence of AD at early age remains poorly understood. The presence of an extra copy of chromosome 21 and overexpression of the genes on the chromosome could be causative. APP, encode by APP gene on chromosome 21, plays an essential role in AD pathogenesis. The triplication of APP gene contributes significantly to the increased prevalence of AD-like dementia in people with DS. Overexpressed APP has been directly linked to the neuropathological abnormality observed in DS with dementia [7]. A number of studies have documented that APP was significantly overexpressed in the brain of adult DS patients [53-55]. APP is the precursor protein for Aβ production. APP overexpression significantly reduced cholinergic neuron’s ability to transport neural growth factor (NGF) in mouse model and consequently led to neuronal cell degeneration [56]. APP overexpression is also the driving force for the excessive production and accumulation of the fibrillary Aβ42 protein, which set off a series of processes and eventually lead to neuronal cell death and AD [57]. Duplication of a short segment of chromosome 21 which spans the APP locus resulted in early onset AD in otherwise phenotypically normal individuals [58, 59]. In addition to APP, other genes located on chromosome 21 such as RCAN1 also contribute significantly to the pathogenesis of AD-like dementia in people with DS [60].
In this study, we have showed that the protein level of BACE1 was significantly increased in DS patients compared to the controls. BACE1, the key protease in vivo for Aβ production, has been directly implicated in AD pathogenesis [61]. BACE1 activity has been established as the rate-limiting step in the process of Aβ production. The protein level of BACE1 could be the major regulatory factor for the production of Aβ besides APP level. Together with increased expression of APP, upregulated BACE1 could significantly lead to overproduction of Aβ and the formation of extracellular amyloid plaque beginning early in life in DS. Previously, we have shown that people with DS had altered BACE1 trafficking and significantly higher levels of mature BACE1 proteins, which contributed to the formation of AD-like pathological changes in DS [7].
In the stable cell line overexpressing BACE1, we found that the protein level of SET was significantly upregulated. This was first detected by 2D SDS-PAGE and then confirmed by Western blotting. No significant change in the mRNA level of SET in the BACE1 overexpressing cells suggest that the regulations of SET expression by BACE1 mainly occur at posttranscriptional level. We also examined the interactions between BACE1 and SET by co-immunoprecipitation. However, no direct interaction was observed (data not shown). We found that both BACE1 and SET protein levels were significantly increased in DS patients. Alteration of SET overexpression could contribute to the occurrence of AD-like dementia. SET, also known as TAF-1β, is a nuclear phosphor-protein widely distributed in a variety of human tissues and involved in various physiological activities [35, 36, 41]. First, SET is a potent inhibitor of protein phosphatase 2A (PP2A) [62, 63]. PP2A is one of the major serine/threonine phosphatases in mammalian cells and the major tau phosphatase in human brain [64-67]. Aberrant phosphorylated Tau is insoluble and more prone to form intraneuronal fibrillary tangles which characterize the histopathological changes in the brain of AD patients [68]. Reduced PP2A activity has been observed to induce Tau hyperphosphorylation in transgenic mice [62, 65] and in AD patients [63]. In DS brain, Tau is hyperphosphorylated. Therefore, it is plausible that significantly upregulated SET in DS contributes to Tau hyperphosphorylation by suppressing PP2A activity. Further studies are warranted to investigate the hypothesis.
SET was implicated in APP-induced neuronal cell death [42]. APP is a substrate of several caspases [69-73]. The fragment Jcaps produced by caspase-induced proteolytic cleavage exhibited strong cytotoxic effect on neuronal cells in AD [73]. Immunoprecipitation and mass spectrometry studies revealed that SET was one of the interacting proteins to facilitate the cytotoxic activity of Jcasp. Downregulation of SET reduced Jcasp-induced cell death [42]. In this study, we observed increased SET expression in the brain of DS patients, which could be associated with accelerated neuronal cell death. On the other hand, we found that SET overexpression inhibited cell proliferation in both peripheral cells and neuronal cells. The doubling time of SET overexpressing cell line was markedly reduced. When we used BrdU labeling as an index of cell proliferation, the amount of BrdU incorporation was significantly less in SET overexpressing cell lines. The growth rate in cells overexpressing BACE1 was also significantly reduced, which could be rescued by SET knockdown. Previous study showed the involvement of SET in forming the endogenous IHAT complex in vivo [41], and SET overexpression could greatly inhibit the process of histone acetylation and gene transcription, and ultimately slowdown cell growth and proliferation. In DS patients, it is conceivable that the upregulated BACE1 and SET could negatively affect the process of neurogenesis of the brain and facilitate neurodegeneration, thus contributing to the early onset of AD-like dementia.
References
Wiseman FK, Alford KA, Tybulewicz VL, Fisher EM (2009) Down syndrome—recent progress and future prospects. Hum Mol Genet 18:R75–R83
Glenner GG, Wong CW (1984) Alzheimer’s disease and Down’s syndrome: sharing of a unique cerebrovascular amyloid fibril protein. Biochem Biophys Res Commun 122:1131–1135
Glenner GG, Wong CW, Quaranta V, Eanes ED (1984) The amyloid deposits in Alzheimer’s disease: their nature and pathogenesis. Appl Pathol 2:357–369
Sun X, Wu Y, Chen B, Zhang Z, Zhou W, Tong Y, Yuan J, Xia K, Gronemeyer H, Flavell RA, Song W (2011) Regulator of calcineurin 1 (RCAN1) facilitates neuronal apoptosis through caspase-3 activation. J Biol Chem 286:9049–9062
Wu Y, Song W (2013) Regulation of RCAN1 translation and its role in oxidative stress-induced apoptosis. FASEB J27:208–221
Sun X, He G, Song W (2006) BACE2, as a novel APP theta-secretase, is not responsible for the pathogenesis of Alzheimer’s disease in Down syndrome. FASEB J20:1369–1376
Sun X, Tong Y, Qing H, Chen CH, Song W (2006) Increased BACE1 maturation contributes to the pathogenesis of Alzheimer’s disease in Down syndrome. FASEB J20:1361–1368
Sun X, Wu Y, Herculano B, Song W (2014) RCAN1 Overexpression exacerbates calcium overloading-induced neuronal apoptosis. PLoS One. doi:10.1371/journal.pone.0095471
Vassar R, Bennett BD, Babu-Khan S, Kahn S, Mendiaz EA, Denis P, Teplow DB, Ross S, Amarante P, Loeloff R, Luo Y, Fisher S, Fuller J, Edenson S, Lile J, Jarosinski MA, Biere AL, Curran E, Burgess T, Louis JC, Collins F, Treanor J, Rogers G, Citron M (1999) Beta-secretase cleavage of Alzheimer’s amyloid precursor protein by the transmembrane aspartic protease BACE. Science 286:735–741
Hussain I, Powell D, Howlett DR, Tew DG, Meek TD, Chapman C, Gloger IS, Murphy KE, Southan CD, Ryan DM, Smith TS, Simmons DL, Walsh FS, Dingwall C, Christie G (1999) Identification of a novel aspartic protease (Asp 2) as beta-secretase. Mol Cell Neurosci 14:419–427
Sinha S, Anderson JP, Barbour R, Basi GS, Caccavello R, Davis D, Doan M, Dovey HF, Frigon N, Hong J, Jacobson-Croak K, Jewett N, Keim P, Knops J, Lieberburg I, Power M, Tan H, Tatsuno G, Tung J, Schenk D, Seubert P, Suomensaari SM, Wang S, Walker D, John V et al (1999) Purification and cloning of amyloid precursor protein beta-secretase from human brain. Nature 402:537–540
Yan R, Bienkowski MJ, Shuck ME, Miao H, Tory MC, Pauley AM, Brashier JR, Stratman NC, Mathews WR, Buhl AE, Carter DB, Tomasselli AG, Parodi LA, Heinrikson RL, Gurney ME (1999) Membrane-anchored aspartyl protease with Alzheimer’s disease beta-secretase activity. Nature 402:533–537
Li Y, Zhou W, Tong Y, He G, Song W (2006) Control of APP processing and Abeta generation level by BACE1 enzymatic activity and transcription. FASEB J20:285–292
Deng Y, Wang Z, Wang R, Zhang X, Zhang S, Wu Y, Staufenbiel M, Cai F, Song W (2013) Amyloid-beta protein (Abeta) Glu11 is the major beta-secretase site of beta-site amyloid-beta precursor protein-cleaving enzyme 1(BACE1), and shifting the cleavage site to Abeta Asp1 contributes to Alzheimer pathogenesis. Eur J Neurosci 37:1962–1969
Luo Y, Bolon B, Kahn S, Bennett BD, Babu-Khan S, Denis P, Fan W, Kha H, Zhang J, Gong Y, Martin L, Louis JC, Yan Q, Richards WG, Citron M, Vassar R (2001) Mice deficient in BACE1, the Alzheimer’s beta-secretase, have normal phenotype and abolished beta-amyloid generation. Nat Neurosci 4:231–232
Cai H, Wang Y, McCarthy D, Wen H, Borchelt DR, Price DL, Wong PC (2001) BACE1 is the major beta-secretase for generation of Abeta peptides by neurons. Nat Neurosci 4:233–234
Roberds SL, Anderson J, Basi G, Bienkowski MJ, Branstetter DG, Chen KS, Freedman SB, Frigon NL, Games D, Hu K, Johnson-Wood K, Kappenman KE, Kawabe TT, Kola I, Kuehn R, Lee M, Liu W, Motter R, Nichols NF, Power M, Robertson DW, Schenk D, Schoor M, Shopp GM, Shuck ME, Sinha S, Svensson KA, Tatsuno G, Tintrup H, Wijsman J, Wright S, McConlogue L (2001) BACE knockout mice are healthy despite lacking the primary beta-secretase activity in brain: implications for Alzheimer's disease therapeutics. Hum Mol Genet 10:1317–1324
Kao S-C, Krichevsky AM, Kosik KS, Tsai L-H (2004) BACE1 Suppression by RNA Interference in Primary Cortical Neurons. J Biol Chem 279:1942–1949
Hussain I, Hawkins J, Harrison D, Hille C, Wayne G, Cutler L, Buck T, Walter D, Demont E, Howes C, Naylor A, Jeffrey P, Gonzalez MI, Dingwall C, Michel A, Redshaw S, Davis JB (2007) Oral administration of a potent and selective non-peptidic BACE-1 inhibitor decreases beta-cleavage of amyloid precursor protein and amyloid-beta production in vivo. J Neurochem 100:802–809
Ly PT, Wu Y, Zou H, Wang R, Zhou W, Kinoshita A, Zhang M, Yang Y, Cai F, Woodgett J, Song W (2013) Inhibition of GSK3beta-mediated BACE1 expression reduces Alzheimer-associated phenotypes. J Clin Invest 123:224–235
Ohno M, Sametsky EA, Younkin LH, Oakley H, Younkin SG, Citron M, Vassar R, Disterhoft JF (2004) BACE1 Deficiency rescues memory deficits and cholinergic dysfunction in a mouse model of Alzheimer’s disease. Neuron 41:27–33
Hu X, Hicks CW, He W, Wong P, Macklin WB, Trapp BD, Yan R (2006) Bace1 modulates myelination in the central and peripheral nervous system. Nat Neurosci 9:1520–1525
Willem M, Garratt AN, Novak B, Citron M, Kaufmann S, Rittger A, DeStrooper B, Saftig P, Birchmeier C, Haass C (2006) Control of peripheral nerve myelination by the {beta}-secretase BACE1. Science 314:664–666
Cheret C, Willem M, Fricker FR, Wende H, Wulf-Goldenberg A, Tahirovic S, Nave KA, Saftig P, Haass C, Garratt AN, Bennett DL, Birchmeier C (2013) Bace1 and Neuregulin-1 cooperate to control formation and maintenance of muscle spindles. EMBO J32:2015–2028
Savonenko AV, Melnikova T, Laird FM, Stewart KA, Price DL, Wong PC (2008) Alteration of BACE1-dependent NRG1/ErbB4 signaling and schizophrenia-like phenotypes in BACE1-null mice. Proc Natl Acad Sci U S A A105:5585–5590
Cai J, Qi X, Kociok N, Skosyrski S, Emilio A, Ruan Q, Han S, Liu L, Chen Z, Bowes Rickman C, Golde T, Grant MB, Saftig P, Serneels L, de Strooper B, Joussen AM, Boulton ME (2012) beta-Secretase (BACE1) inhibition causes retinal pathology by vascular dysregulation and accumulation of age pigment. EMBO Mol Med 4:980–991
Acquati F, Accarino M, Nucci C, Fumagalli P, Jovine L, Ottolenghi S, Taramelli R (2000) The gene encoding DRAP (BACE2), a glycosylated transmembrane protein of the aspartic protease family, maps to the down critical region. FEBS Lett 468:59–64
Bennett BD, Babu-Khan S, Loeloff R, Louis JC, Curran E, Citron M, Vassar R (2000) Expression analysis of BACE2 in brain and peripheral tissues. J Biol Chem 275:20647–20651
Lin X, Koelsch G, Wu S, Downs D, Dashti A, Tang J (2000) Human aspartic protease memapsin 2 cleaves the beta-secretase site of beta-amyloid precursor protein. Proc Natl Acad Sci U S A A97:1456–1460
Solans A, Estivill X, de La Luna S (2000) A new aspartyl protease on 21q22.3, BACE2, is highly similar to Alzheimer’s amyloid precursor protein beta-secretase. Cytogenet Cell Genet 89:177–184
Liu X, Wang Z, Wu Y, Wang J, Song W (2013) BACE2 degradation mediated by the macroautophagy-lysosome pathway. Eur J Neurosci 37:1970–1977
Sun X, Wang Y, Qing H, Christensen MA, Liu Y, Zhou W, Tong Y, Xiao C, Huang Y, Zhang S, Liu X, Song W (2005) Distinct transcriptional regulation and function of the human BACE2 and BACE1 genes. FASEB J19:739–749
von Lindern M, van Baal S, Wiegant J, Raap A, Hagemeijer A, Grosveld G (1992) Can, a putative oncogene associated with myeloid leukemogenesis, may be activated by fusion of its 3’ half to different genes: characterization of the set gene. Mol Cell Biol 12:3346–3355
Asaka MN, Murano K, Nagata K (2008) Sp1-mediated transcription regulation of TAF-Ialpha gene encoding a histone chaperone. Biochem Biophys Res Commun 376:665–670
Nagata K, Kawase H, Handa H, Yano K, Yamasaki M, Ishimi Y, Okuda A, Kikuchi A, Matsumoto K (1995) Replication factor encoded by a putative oncogene, set, associated with myeloid leukemogenesis. Proc Natl Acad Sci U S A A92:4279–4283
Adachi Y, Pavlakis GN, Copeland TD (1994) Identification of in vivo phosphorylation sites of SET, a nuclear phosphoprotein encoded by the translocation breakpoint in acute undifferentiated leukemia. FEBS Lett 340:231–235
Nagata K, Saito S, Okuwaki M, Kawase H, Furuya A, Kusano A, Hanai N, Okuda A, Kikuchi A (1998) Cellular localization and expression of template-activating factor I in different cell types. Exp Cell Res 240:274–281
Li M, Makkinje A, Damuni Z (1996) Molecular identification of I1PP2A, a novel potent heat-stable inhibitor protein of protein phosphatase 2A. Biochemistry 35:6998–7002
Li M, Damuni Z (1998) I1PP2A and I2PP2A. Two potent protein phosphatase 2A-specific inhibitor proteins. Methods Mol Biol 93:59–66
Saito S, Miyaji-Yamaguchi M, Shimoyama T, Nagata K (1999) Functional domains of template-activating factor-I as a protein phosphatase 2A inhibitor. Biochem Biophys Res Commun 259:471–475
Seo SB, McNamara P, Heo S, Turner A, Lane WS, Chakravarti D (2001) Regulation of histone acetylation and transcription by INHAT, a human cellular complex containing the set oncoprotein. Cell 104:119–130
Madeira A, Pommet JM, Prochiantz A, Allinquant B (2005) SET protein (TAF1beta, I2PP2A) is involved in neuronal apoptosis induced by an amyloid precursor protein cytoplasmic subdomain. FASEB J19:1905–1907
Qu D, Zhang Y, Ma J, Guo K, Li R, Yin Y, Cao X, Park DS (2007) The nuclear localization of SET mediated by impalpha3/impbeta attenuates its cytosolic toxicity in neurons. J Neurochem 103:408–422
Wang X, Blanchard J, Kohlbrenner E, Clement N, Linden RM, Radu A, Grundke-Iqbal I, Iqbal K (2010) The carboxy-terminal fragment of inhibitor-2 of protein phosphatase-2A induces Alzheimer disease pathology and cognitive impairment. FASEB J24:4420–4432
Arnaud L, Chen S, Liu F, Li B, Khatoon S, Grundke-Iqbal I, Iqbal K (2011) Mechanism of inhibition of PP2A activity and abnormal hyperphosphorylation of tau by I2(PP2A)/SET. FEBS Lett 585:2653–2659
Bolognin S, Blanchard J, Wang X, Basurto-Islas G, Tung YC, Kohlbrenner E, Grundke-Iqbal I, Iqbal K (2012) An experimental rat model of sporadic Alzheimerֹ’s disease and rescue of cognitive impairment with a neurotrophic peptide. Acta Neuropathol 123:133–151
Yu G, Yan T, Feng Y, Liu X, Xia Y, Luo H, Wang JZ, Wang X (2013) Ser9 phosphorylation causes cytoplasmic detention of I2PP2A/SET in Alzheimer disease. Neurobiol Aging 34:1748–1758
Qing H, Zhou W, Christensen MA, Sun X, Tong Y, Song W (2004) Degradation of BACE by the ubiquitin-proteasome pathway. FASEB J18:1571–1573
Sun X, He G, Qing H, Zhou W, Dobie F, Cai F, Staufenbiel M, Huang LE, Song W (2006) Hypoxia facilitates Alzheimer’s disease pathogenesis by up-regulating BACE1 gene expression. Proc Natl Acad Sci U S A A103:18727–18732
Zigman WB, Schupf N, Sersen E, Silverman W (1996) Prevalence of dementia in adults with and without Down syndrome. Am J Ment Retard 100:403–412
Visser FE, Aldenkamp AP, van Huffelen AC, Kuilman M, Overweg J, van Wijk J (1997) Prospective study of the prevalence of Alzheimer-type dementia in institutionalized individuals with Down syndrome. Am J Ment Retard 101:400–412
Nieuwenhuis-Mark RE (2009) Diagnosing Alzheimer’s dementia in Down syndrome: problems and possible solutions. Res Dev Disabil 30:827–838
Rumble B, Retallack R, Hilbich C, Simms G, Multhaup G, Martins R, Hockey A, Montgomery P, Beyreuther K, Masters CL (1989) Amyloid A4 protein and its precursor in Down’s syndrome and Alzheimer’s disease. N Engl J Med 320:1446–1452
Beyreuther K, Pollwein P, Multhaup G, Monning U, Konig G, Dyrks T, Schubert W, Masters CL (1993) Regulation and expression of the Alzheimer’s beta/A4 amyloid protein precursor in health, disease, and Down's syndrome. Ann N Y Acad Sci 695:91–102
Cheon MS, Dierssen M, Kim SH, Lubec G (2008) Protein expression of BACE1, BACE2 and APP in Down syndrome brains. Amino Acids 35:339–343
Salehi A, Delcroix JD, Belichenko PV, Zhan K, Wu C, Valletta JS, Takimoto-Kimura R, Kleschevnikov AM, Sambamurti K, Chung PP, Xia W, Villar A, Campbell WA, Kulnane LS, Nixon RA, Lamb BT, Epstein CJ, Stokin GB, Goldstein LS, Mobley WC (2006) Increased App expression in a mouse model of Down's syndrome disrupts NGF transport and causes cholinergic neuron degeneration. Neuron 51:29–42
Wisniewski KE, Dalton AJ, McLachlan C, Wen GY, Wisniewski HM (1985) Alzheimer’s disease in Down’s syndrome: clinicopathologic studies. Neurology 35:957–961
Rovelet-Lecrux A, Hannequin D, Raux G, Le Meur N, Laquerriere A, Vital A, Dumanchin C, Feuillette S, Brice A, Vercelletto M, Dubas F, Frebourg T, Campion D (2006) APP locus duplication causes autosomal dominant early-onset Alzheimer disease with cerebral amyloid angiopathy. Nat Genet 38:24–26
Sleegers K, Brouwers N, Gijselinck I, Theuns J, Goossens D, Wauters J, Del-Favero J, Cruts M, van Duijn CM, Van Broeckhoven C (2006) APP duplication is sufficient to cause early onset Alzheimer’s dementia with cerebral amyloid angiopathy. Brain 129:2977–2983
Yili Wu PTTL, Weihong Song (2014) Aberrant Expression of RCAN1 in Alzheimer’s pathogenesis: a new molecular mechanism and a novel drug target. Mol Neurobiol
Sun X, Bromley-Brits K, Song W (2012) Regulation of beta-site APP-cleaving enzyme 1 gene expression and its role in Alzheimer’s disease. J Neurochem 120(Suppl 1):62–70
Li M, Makkinje A, Damuni Z (1996) The myeloid leukemia-associated protein SET is a potent inhibitor of protein phosphatase 2A. J Biol Chem 271:11059–11062
Neviani P, Santhanam R, Trotta R, Notari M, Blaser BW, Liu S, Mao H, Chang JS, Galietta A, Uttam A, Roy DC, Valtieri M, Bruner-Klisovic R, Caligiuri MA, Bloomfield CD, Marcucci G, Perrotti D (2005) The tumor suppressor PP2A is functionally inactivated in blast crisis CML through the inhibitory activity of the BCR/ABL-regulated SET protein. Cancer Cell 8:355–368
Gong CX, Lidsky T, Wegiel J, Zuck L, Grundke-Iqbal I, Iqbal K (2000) Phosphorylation of microtubule-associated protein tau is regulated by protein phosphatase 2A in mammalian brain. Implications for neurofibrillary degeneration in Alzheimer’s disease. J Biol Chem 275:5535–5544
Kins S, Crameri A, Evans DR, Hemmings BA, Nitsch RM, Gotz J (2001) Reduced protein phosphatase 2A activity induces hyperphosphorylation and altered compartmentalization of tau in transgenic mice. J Biol Chem 276:38193–38200
Drewes G, Mandelkow EM, Baumann K, Goris J, Merlevede W, Mandelkow E (1993) Dephosphorylation of tau protein and Alzheimer paired helical filaments by calcineurin and phosphatase-2A. FEBS Lett 336:425–432
Goedert M, Jakes R, Qi Z, Wang JH, Cohen P (1995) Protein phosphatase 2A is the major enzyme in brain that dephosphorylates tau protein phosphorylated by proline-directed protein kinases or cyclic AMP-dependent protein kinase. J Neurochem 65:2804–2807
Hanger DP, Brion JP, Gallo JM, Cairns NJ, Luthert PJ, Anderton BH (1991) Tau in Alzheimer’s disease and Down’s syndrome is insoluble and abnormally phosphorylated. Biochem J 275(Pt 1):99–104
Barnes NY, Li L, Yoshikawa K, Schwartz LM, Oppenheim RW, Milligan CE (1998) Increased production of amyloid precursor protein provides a substrate for caspase-3 in dying motoneurons. J Neurosci 18:5869–5880
LeBlanc A, Liu H, Goodyer C, Bergeron C, Hammond J (1999) Caspase-6 role in apoptosis of human neurons, amyloidogenesis, and Alzheimer’s disease. J Biol Chem 274:23426–23436
Pellegrini L, Passer BJ, Tabaton M, Ganjei JK, D’Adamio L (1999) Alternative, non-secretase processing of Alzheimer's beta-amyloid precursor protein during apoptosis by caspase-6 and -8. J Biol Chem 274:21011–21016
Weidemann A, Paliga K, Durrwang U, Reinhard FB, Schuckert O, Evin G, Masters CL (1999) Proteolytic processing of the Alzheimer’s disease amyloid precursor protein within its cytoplasmic domain by caspase-like proteases. J Biol Chem 274:5823–5829
Lu DC, Rabizadeh S, Chandra S, Shayya RF, Ellerby LM, Ye X, Salvesen GS, Koo EH, Bredesen DE (2000) A second cytotoxic proteolytic peptide derived from amyloid beta-protein precursor. Nat Med 6:397–404
Acknowledgments
We thank CHiBI Proteomics Core Facility of University of British Columbia for mass spectrometric analysis of the 2D gel dots. We thank Dr. Suzanne C. Perry for the discussions. This work is supported by the Canadian Institutes for Health Research (CIHR) (MOP-97825 and TAD-117948), the National Natural Science Foundation of China (NSFC) (NSFC81161120498), and the Jack Brown and Family Alzheimer’s Research Foundation. WS is the holder of the Tier 1 Canada Research Chair in Alzheimer’s disease. XZ is the recipient of the Arthur and June Willms Fellowship. XD is supported by the Chinese Scholarship Council awards.
Conflict of Interest
The authors declare no conflict of interest.
Author information
Authors and Affiliations
Corresponding author
Additional information
X. Zhang, Y. Wu and X. Duan contributed equally to this work.
Rights and permissions
About this article

Cite this article
Zhang, X., Wu, Y., Duan, X. et al. Upregulation of SET Expression by BACE1 and its Implications in Down Syndrome. Mol Neurobiol 51, 781–790 (2015). https://doi.org/10.1007/s12035-014-8782-x
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s12035-014-8782-x