
Abstract
Gaining a basic understanding of the inhibitory molecules and the intracellular signaling involved in axon development and repulsion after neural lesions is of clear biomedical interest. In recent years, numerous studies have described new molecules and intracellular mechanisms that impair axonal outgrowth after injury. In this scenario, the role of glycogen synthase kinase 3 beta (GSK3β) in the axonal responses that occur after central nervous system (CNS) lesions began to be elucidated. GSK3β function in the nervous tissue is associated with neural development, neuron polarization, and, more recently, neurodegeneration. In fact, GSK3β has been considered as a putative therapeutic target for promoting functional recovery in injured or degenerative CNS. In this review, we summarize current understanding of the role of GSK3β during neuronal development and regeneration. In particular, we discuss GSK3β activity levels and their possible impact on cytoskeleton dynamics during both processes.
Similar content being viewed by others
Avoid common mistakes on your manuscript.
Introduction
It is currently believed that the glycogen synthase kinase 3 (GSK3) family of intracellular kinases are regulators of the physiology of developing and adult central nervous system (CNS) neurons. In fact, absence or misregulation of GSK3 functions has been observed in several CNS diseases such as Alzheimer’s disease (e.g., Ref. [1]), mood disorders and schizophrenia (e.g., Refs. [2, 3]), and anxiety or behavioral abnormalities associated with serotonin deficiency [4, 5]. The relevance of this family of kinases is demonstrated by emerging roles in neuronal physiology identified in recent years (see Refs. [6, 7] for review). For example, GSK3 kinases control gene transcription, axonal transport, and cytoskeletal dynamics in growth cones (see Ref. [8] for a recent review). Indeed, GSK3 kinases serve as key molecules in the coordination of cytoskeletal elements by controlling microtubule dynamics and assembly via regulation of several microtubule-binding proteins (MBPs). Thus, changes in GSK3 participation in the control of the neuronal cytoskeleton have been associated with neurodegeneration. In addition, several studies have implicated the modulation of the GSK3 activity and the upstream or downstream regulators in axonal regeneration after lesion (e.g., Refs. [9–13]). However, the role of GSK3 in neuronal shape maintenance (as a regulator of neuronal polarization) and plasticity is unclear due to different reports with contradictory findings. In this review, we would like to present an overview of the published roles of GSK3 beta (GSK3β) signaling in neuronal growth and regeneration in order to clarify these discrepancies. We will focus on the most relevant data in both neuron development and axon regeneration after injury. In the review, we will use the term GSK3β to refer directly to the kinase and GSK3 when the reported effects are not clearly associated specifically with GSK3β. Due to the large amount of information concerning GSK3 family of kinases and their various roles in different processes, we refer the reader to the reviews by Beurel et al. [14] and Lee and Kim [15] for the particular roles of GSK3 kinases in the immune system and in glucose metabolism, respectively, and to the review by Coen and Goedert [16] for the therapeutic potential of GSK3 inhibitors in several diseases.
GSK3β, a Fine-Tuned Kinase of the GSK3 Family in Neural Tissue
The GSK3 proteins are serine/threonine (Ser/Thr) kinases originally identified as key enzymes in glycogen metabolism [17, 18]. Two isoforms, GSK3α and GSK3β, are encoded by different genes with 95 % identity in their kinase domains and 85 % sequence homology [19, 20]. GSK3α and GSK3β are expressed in the central nervous system and participate in several non-redundant functions [21]. GSK3 kinases are implicated in multiple processes during neural development and in adult stages including neurogenesis, neuroprotection, migration, neuronal polarization, and axonal growth and maintenance, as well as intracellular transport. Phosphorylation of most GSK3 substrates requires prior proline phosphorylation by a priming kinase [e.g. Casein kinase I-II, cyclin-dependent kinase 5 (Cdk5) or dual-specificity tyrosine phosphorylation-regulated kinase (DYRK)], but unprimed substrates have also been reported for some proteins (e.g., Tau, phosphorylated at Ser396/404) [22]. GSK3β is the most commonly expressed isoform in the nervous system [23] and its spliced variant, GSK3β2, is the most enriched isoform in neurons of humans and rodents [24]. GSK3β expression is widespread throughout all regions of the developing and adult brain, although it is greatest in the hippocampus, thalamus, cortex, and Purkinje cells of the cerebellum in the adult (e.g., Ref. [25]). At the cellular level, GSK3β is present throughout the cell body and processes (dendrites and axon) of post-mitotic neurons (e.g., Ref. [25]).
Neuronal GSK3β activity is under the control of numerous mechanisms and signaling pathways. In fact, GSK3β is regulated both positively and negatively by several kinases including protein kinase B (Akt), p38 mitogen-activated protein kinase (MAPK), and protein tyrosine phosphatase (PTPase; Fig. 1). Thus, whereas the phosphorylation of tyrosine 216 residue (Tyr216) leads to GSK3β activation, phosphorylation of Serine 9 residue (Ser9) reduces its activity [26, 27] (Fig. 1). In some cases, phosphorylation of Ser9 is the result of the combined action of two kinases (i.e., ERK1/2 and p90RSK [28]). In contrast to inhibitory regulation by Ser9 mainly associated in the literature with PKB/Akt activity [29], GSK3β activity can also be facilitated by auto-phosphorylation [30], by alterations in intracellular calcium levels [31] and by other tyrosine kinases like proline-rich tyrosine kinase 2 (Pyk2) through Tyr216 phosphorylation [32] (see Fig. 1 for a general overview of its regulation). In addition, the regulation of the GSK3β activity may also include protein–protein interaction (e.g., with Frat1) [33, 34].

A general overview of GSK3β regulation. GSK3β is subject to multiple regulatory mechanisms. Phosphorylation of Ser9 is probably the most important regulatory mechanism. Several kinases are capable of mediating this modification, including p70 S6 kinase, extracellular signal-regulated kinases (ERKs), p90Rsk (also called MAPKAP kinase-1), Akt (PKB), certain isoforms of protein kinase C (PKC), and cyclic AMP-dependent protein kinase (PKA). Stimulation of pGSK3β (Tyr216) could be mediated by alterations in intracellular calcium levels and a calcium-dependent tyrosine kinase, proline-rich tyrosine kinase 2 (PYK2), or Fyn, a member of the Src tyrosine family. pGSK3β (Tyr216) is also subject to the regulation of mitogen-activated protein kinase (MEK1/2), PTPase, and even autophosphorylation. Moreover, the protein phosphatases 1 and 2A (PP1 and PP2A) also regulate GSK3β phosphorylation levels. However, control of GSK3β activity is through its main targets (MAPs or transcription factors), finally leading to control of neurogenesis, neuronal polarization, and axonal outgrowth. See references in text
GSK3β activity has been shown to be a key regulator in signaling pathways in neurons triggered by many extracellular molecules such as Wnt, neural growth factor (NGF), fibroblast growth factor (FGF), epidermal growth factor (EGF), and sonic hedgehog (Shh) [20, 35–37]. These signaling mechanisms have been largely characterized as regulators of proliferation, differentiation, and maturation of neural progenitors (e.g., Refs. [6, 38, 39]). Indeed, GSK3β exhibits crosstalk with several signaling pathways and is considered to have the potential to integrate these pathways into the control of neuronal physiology, particularly at the level of microtubular cytoskeleton dynamics.
GSK3β Activity in Neuronal Polarization and Plasticity
In developing neurons, the involvement of GSK3β in neuronal polarization coordinating mammalian target of rapamycin (mTOR), phosphoinositide 3-kinase (PI3K), Par3/6, protein kinase A (PKA)-LKB1, and Rho-GTPase signaling pathways has been reported (see also Ref. [7] for a recent review). Today, it is well known that neuron polarization begins with the rapid extension of a single neurite to produce an axon, while the remaining neurites differentiate into dendrites. This is followed by the selective localization of molecules that are responsible for differential growth and specific functions of the axon and dendrites [40]. During this process, polarized neurons develop axonal and dendritic trees by elongation and branching. In these processes, GSK3β activity plays a role at two levels: (i) controlling the phosphorylation of different proteins associated with the microtubular cytoskeleton and (ii) modulating local protein synthesis and degradation [41] Indeed, GSK3β substrates include MBPs such as adenomatous polyposis coli (APC), collapsin response mediator proteins 2 and 4 (CRMP2 and CRMP4), microtubule-associated proteins 1B (MAP1B) and Tau, and cytoplasmic linker proteins (CLIP-associate protein), as well as other transcription factors and kinases putatively involved in cytoskeleton modifications (Fig. 1). In fact, functions of GSK3 during polarization are not exclusive since it runs in parallel with other kinases localized in growing neurites also acting on cytoskeleton polymerization (e.g., microtubule affinity-regulating kinase (MARK) [42], serine/threonine kinase LKB1, and SAD kinases [43]).
The activation and modulation of GSK3 yields numerous outcomes during neuronal polarization that may trigger both axon growth promotion and inhibition depending on the cellular model and particular experimental condition. For example, several studies have suggested that reduced GSK3β activity is required for initial axon formation and elongation (e.g., Refs. [44–48]) and that increased GSK3β activity leads to neurite retraction during polarization (e.g., Ref. [49]). Yoshimura et al. [44] reported that neurotrophic treatment of cultured neurons leads to axon elongation and branching by inhibiting GSK3β via PI3K/Akt, thereby reducing phosphorylation of CRMP2 and decreasing microtubule polymerization (Fig. 3). In fact, it has been reported that both GSK3α and GSK3β play roles in axon formation [48]. However, a second group of studies reported opposite results indicating that elevated activity of GSK3 leads to increased axon elongation. As an example of this, it has been described how high expression and phosphorylation of particular GSK3β targets (e.g., CMRP2 or β-adducin) increased neurite extension in cultured cortical neurons and SH-SY5Y neuroblastoma cells [50–52]. Similarly, it has been described how pharmacological and shRNA blockage of GSK3 activity induces neurite retraction [53, 54]. Although well developed, most of these results have been derived in cultured neurons of neural cells under different experimental conditions.
In an interesting study, Garrido et al. [48] determined a “critical period” of GSK3 activity in polarizing neurons of 24 h. Thus, inhibition of GSK3 activity after this period does not compromise the elongation process but increases axonal branching. In fact, two proteins, Katanin (a severing microtubular protein) and KIF2 (a microtubule depolymerase that modulates axon branching), are potential GSK3 substrates, suggesting that inactivation of GSK3 may regulate their functions to control axon branching [55]. These results defined a dual participation of GSK3 kinases during polarization at the level of initiation of axon growth from undifferentiated neurites and the branching modeling of the growing axon.
However, these challenging questions forced researchers to move forward with in vivo analysis of the function of GSK3 in mouse models. In particular, recent results obtained using conditional GSK3β expressing mice are of particular interest and reinforce some of the previous results. Indeed, functions of GSK3β during neurite extension and branching in newborn neurons in the hippocampal dentate gyrus have been reported [56]. GSK3β overexpression under the CaMK-II promoter in newborn granule cells of the dentate gyrus causes dramatic changes in neurite extension and synaptic connectivity (Fig. 2). These effects can be reversed by down-regulating GSK3β in the same cells [56]. In fact, GSK3β-overexpressing granule cells displayed a relevant dendritic tree remodeling in the adult hippocampus after maturation, mimicking the remodeling observed in Alzheimer´s disease. In this respect, general or prolonged inhibition of GSK3β might induce multiple points throughout the axon/neurite capable of supporting efficient microtubule formation, resulting in more branching instead of axonal elongation (e.g., Refs. [44, 46, 48]; Fig. 2). These effects have been reported in vivo by Llorens-Martin and co-workers [56] who found an enlarged highly branched proximal dendritic tree and a drastic decrease in distal branching of the granule cells in these mice as previously shown in vitro (see above).

GSK3β activity in neurite growth and branching during development and in the adult. GSK3β plays a key role in neuronal polarization and neurite outgrowth in developing neurons. The suppression of GSK3β activity in cultured neurons induces axon formation and elongation. Once the axon has been determined in developing neurons, a compartmentalization in the neuron takes place that is maintained by the differential spatiotemporal regulation of GSK3β. Moreover, Llorens-Martin et al. recently demonstrated that GSK3β overexpression in newly generated granule neurons induces relevant branching in proximal dendrites rather than elongation. Then further GSK3β inhibition leads to a morphological rescue phenotype. Thus, it appears that GSK3β activity above or below a baseline level of activity leads to alterations in the balance between elongation and branching
These results make it clear that the differences between specific domains of the growing neurite and differentiating axon may arise because developing neurons are able to spatially regulate GSK3β activity in different cellular domains along growing neurites. This may also come about from a particular distribution of receptors and/or intracellular partners modulating local activity of GSK3β toward specific domains of neuronal cytoskeleton. Particularly relevant is a putative scenario in which second messengers may act locally regulating kinases involved in neuronal polarization. Two examples of this are worth citing: Barnes et al. [43] determined that cAMP levels act through phosphorylated LKB1 and GSK3β to promote axon initiation, whereas cGMP-mediated signaling suppresses axon formation via reciprocal down-regulation of cAMP/PKA-dependent phosphorylation of LKB1 and GSK-3β [43]. Second, Jiang et al. [46] demonstrated that phosphatase and tensin homolog (PTEN), a phosphatase that converts PIP3 to PIP2, showed low activity in the axon compared to dendrites in developing neurons. This low activity of PTEN attenuates PIP3-dependent local activation of Akt in neurites other than the growing axon, leading to GSK3β inhibition and promotion of axonal growth and enhancement of neuronal polarization (Fig. 2). In support of this notion, it has recently been described how SIRT-1, a deacetylase that is mainly located in the axon and growth cones of developing neurons, inhibits GSK3 activity by deacetylating and activating Akt, leading to axon regrowth [57]. These actions are particularly relevant when considering axon formation and growth under the effects of some long-range axonal wiring cues during neural development since most of them act through cAMP levels [32, 58], in contrast with local control of neurite/axon formation by the balanced control of cGMP/cAMP levels. This is in line with a current notion that suggests that the neuronal growth cone may also sense different concentrations of some extracellular factors that modulate different intracellular mechanisms including GSK3 activity. This new idea implies that a single cue is able to trigger different spatial neuronal responses depending on their extracellular concentration. An example can be found in the growth cone responses to secreted semaphorins (e.g., Sema3A). Sema3A acts through Neuropilin-1 and Plexins, impairing neurite extension by modulating GSK3β activity [59–61]. However, a recent study by Manns et al. [62] demonstrated that while mTOR inhibition did not influence GSK3β activation, GSK3β inhibition promoted mTOR activity in cultured dorsal root ganglion (DRG) neurons exposed to different concentrations of Sema3A. Furthermore, regarding GSK3β activity and its reported spatiotemporal regulation, what is clear is that a simple analysis of the overall phosphorylation status of the kinase in the neuron does not correlate fully with the observed response. For example, this has been demonstrated for Netrin-1, a chemoattractive molecule for commissural axons that is able to guide axons by small changes in local concentration [63]. In fact, when analyzed after treatment, Netrin-1 acts by increasing the phosphorylation of GSK3β at Ser9 and Thr216 and by modulating MAP1B phosphorylation [64], which further mediates chemoattraction in vitro as well as in vivo (e.g., Refs. [64, 65]). Parallel to the spatiotemporal regulation, the specific experimental conditions may lead to an increase in or blocking of axonal growth depending on the substrates involved and the extent of GSK3β inhibition [54, 66]. In fact, today, we cannot rule out the scenario that hypothesizes that a decrease in GSK3β activity after intense treatment or local influence might reduce GSK3β activity to baseline levels, which would then promote axonal growth or branching. Furthermore, although examples have been reported for specific cell types (e.g., DRG neurons, [62] and hippocampal neurons [32]), we still do not know whether the mechanisms that regulate GSK3β also occur in all subsets of developmental neurons in the same manner.
A Brief Overview of the Molecular Factors that Impair Axon Regrowth After Adult CNS Lesion
In the adult mammalian central nervous system (CNS), axons have a limited capacity for regrowth after lesion (e.g., Refs. [67–69]). Several intrinsic and extrinsic neural processes converge in the absence of axonal regeneration including: (i) low potential to recapitulate the intrinsic developmental program in adult CNS neurons, (ii) low renewal of damaged neurons with endogenous stem cells, which impairs putative autonomous cell therapy in most CNS regions, and (iii) the overexpression of numerous inhibitory molecules by reactive cells after adult CNS lesions. These reactive cells form the meningoglial scar, a dense meshwork of astrocytes, oligodendrocyte precursors, microglia, and infiltrated meningeal cells (e.g., Refs. [70–72]). Indeed, this scar is considered both a physical and a biochemical barrier to axon regeneration and functional recovery, although some reports suggest that it may have beneficial effects shortly after the lesion (see Ref. [70] for review). Today, it is generally accepted that the molecular environment around the meningoglial scar is, in combination with the loss of intrinsic growth capacity of the injured axons, a determinant of the ability or failure of adult axons to regenerate (e.g., Refs. [73–77]). As indicated, the scar produces inhibitory and nonpermissive factors such as semaphorins, ephrins, tenascins, chondroitin sulfate proteoglycans (CSPGs), and myelin-associated inhibitors (MAIs) [71]. In this review we will focus on semaphorins, CSPGs, and MAIs as examples of inhibitory signals that may modulate GSK3 and especially GSK3β activity.
Semaphorins and Their Receptors
Although initially characterized as repulsive axonal guidance cues, semaphorins are now regarded as relevant contributors to morphogenesis and homeostasis for a wide range of tissue types (e.g., see Ref. [78] for a detailed description of semaphorins and their receptors). Semaphorin-mediated long- and short-range repulsive, and attractive, guidance has a profound influence on neuronal physiology (see Refs. [79, 80] for review). In fact, in recent years, several members of the superfamily of semaphorins have been associated with neural lesions, and some of these directly with inhibitory processes after injury. For example, Sema3A, Sema3C, Sema6B, and Sema6C are overexpressed after telencephalic lesions in several reactive as well as nonreactive cells [59, 81–84]. Sema6C and Sema3A lead to GSK3β-dependent axonal growth cone collapse [82, 85]. Moreover, it has also been reported that three other semaphorins, Sema4D, Sema7A, and Sema6A, are overexpressed after spinal cord injury [86–88] and may act though the same signaling pathways as Sema6C and Sema3A (GSK3β activity modulation) leading to inhibitory processes.
Chondroitin Sulfate Proteoglycans and Their Receptors
Proteoglycans (PGs) are highly glycosylated proteins synthesized by reactive astrocytes [89, 90], meningeal cells [91, 92], NG2 cells [93], and macrophages after lesion [94]. They are made up of a core protein and long unbranched glycosaminoglycan (GAG) chains such as chondroitin sulfate (CS) and heparan sulfate (HS). CSPGs are involved in several neuronal processes like migration, axon guidance, and the promotion or inhibition of neuritic growth (e.g., see Ref. [95]) during CNS development. However, CSPGs are also overexpressed in many types of CNS lesion (e.g., Refs. [96–101]). The modes of action induced by CSPGs have not yet been fully determined. Most of their inhibitory effect depends on GAG chains (e.g., Ref. [102]), although for particular CSPGs, such as NG2, the protein core also displays inhibitory properties [103]. Two proteins, the transmembrane protein tyrosine phosphatase 3 (PTPσ) [104–107] and the leukocyte common antigen-related phosphatase (LAR), are CSPG receptors that may modulate GSK3 activity [108]. In fact, from a molecular point of view CSPGs initially diminish Akt and GSK3β-Ser9 phosphorylation in cell lines and cultured neurons [10].
MAIs and Their Receptors
It is well known that MAIs are highly regulated after injury by glial cells, oligodendrocytes, and, in the case of MAIs (Nogo-A), lesioned neurons (e.g., Refs. [11, 109–111]). Three MAIs have been studied in detail: Nogo-A, the myelin-associated glycoprotein (MAG), and the GPI–linked oligodendrocyte-myelin glycoprotein (OMgp) [112–114]. These molecules impair axon regrowth mainly by acting on a common neuronal receptor complex, the Nogo receptor complex. This receptor is formed by the GPI-anchored protein Nogo receptor 1 (NgR1) and three putative co-receptors: (p75 (NTR), TAJ/TROY and Lingo-1) [112–114]. In addition, new ligands have recently been reported as acting through the Nogo receptor complex (e.g., Refs. [115–118]) and new receptors for MAIs have also been described [119–125]. The participation of these ligands and receptors and their associated intracellular signaling mechanisms in axon regeneration is under constant evaluation (e.g., Refs. [72, 126]). Most studies have reported MAI and receptor functions in the CNS, although emerging roles in PNS regeneration have also been described [127]. The effects of MAIs in GSK3 activity are discussed below, since dual effects have been described.
Modulation of GSK3β Activity by Lesion-Associated Molecules Through Akt/GSK3β and Akt/mTOR Pathways
From several studies we know that PI3K/Akt signaling is one of the most studied targets of inhibitory molecules after neural lesions. Two pathways can be activated by PI3K/Akt: i) the Akt/GSK3β pathway and ii) the Akt/mTOR pathway. The modulation of these pathways by inhibitory molecules has been reported in several studies. Concerning Akt/GSK3β, some studies reported that secreted semaphorins activated GSK3 (e.g., Sema3F [128] or Sema3A [85, 129, 130]). Uchida et al. report that the sequential phosphorylation of CRMP2 by Cdk5 and GSK3β mediates growth cone collapse in response to Sema3A in mouse DRG neurons. Interestingly, they also report the formation of a complex involving GSK3β, Axin-1, and β-catenin which may play an important role in Sema3A signaling involving the endocytic pathway [85].
With respect to CSPG and the Akt/GSK3β pathway, Fisher et al. [108] found a reduction in Akt phosphorylation after CSPG treatment in cultured cerebellar granule neurons (CGNs). Then, reduced Akt activity correlated with enhanced GSK3β activity. In addition, using pharmacological treatment, Gao et al. [131] also showed that treatment with Amphotericin B, a polyene fungal antibiotic [132], induces an increase in the regenerative process through Akt activation and GSK3β inhibition after CSPG stimulation in neurons. Interestingly, a recent study reported that NgR1 and 3 are also functional receptors for CSPGs, since they interact with CSPG GAG chains and might trigger a signaling pathway in which the Rho family is involved [123]. Hence, there may be collaboration between the NgR family and LAR family of receptors since an additive effect on the number and length of regenerating axons following CNS injury was observed in NgR1-3/RPTPσ -/- knockout mice [123]. Based on this evidence we may suppose that CSPGs affect GSK3 and particularly GSK3β activity both through known [e.g., NgR1, NgR3, integrins, paired immunoglobuline-like receptor B (PirB), PTPσ, LAR] and unknown receptors.
In the Akt/mTOR mechanism, PTEN has been shown to attenuate the regrowth of injured CNS axons by suppressing mTOR [133]. Indeed, PTEN deletion in injured mature retinal ganglion neurons (RGCs) increases axonal regeneration by over-activating Akt and inhibiting GSK3β activity [133, 134]. In line with these results, Liu and co-workers also show that PTEN deletion leads to a regenerative response of corticospinal neurons in the spinal cord [12]. The marked increase in compensatory sprouting of intact corticospinal tract (CST) axons and regenerative growth of injured CST axons after PTEN deletion suggests that these two forms of regrowth have similar underlying mechanisms [12]. Unfortunately, however, the chronic loss of PTEN triggers unwanted effects, since long-term knock-out or knock-down in particular tissues may lead to neoplasias, such as glial tumors, which promote astrocyte hypertrophy and proliferation (see Ref. [135]).
MAIs and GSK3 Activity After Lesion: a Scenario Requiring Clarification
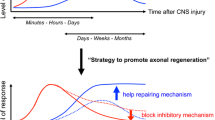
Some published data have directly associated GSK3β activity with MAIs. However, as observed above for neuronal polarization, there are some discrepancies in the literature concerning MAIs and GSK3 activity. In fact, several studies have shown increased GSK3β activity by stimulation with MAG or myelin extracts [10, 11, 136], whereas others found decreased GSK3β activity after stimulation with myelin extracts and the Nogo-P4 inhibitory peptide (residues 31–55 of the NogoA protein) [137] or OMgp [9] (Fig. 3). As an example of the first group of studies, Dill and co-workers demonstrated that the acute stimulation of PC12 cells and CGNs by MAG increased Akt and GSK3β-Ser9 phosphorylation [10]. Similar inhibition of GSK3 was described by Shen et al. [136] in N2a cells (see also Fig. 3 as example of the results published in Ref. 9). More relevantly, the published data attribute to GSK3β the main role of inhibiting axon regrowth after long-term exposure to inhibitory cues (see above), which may conflict with MAIs activities (in some studies). However, we should consider both the temporal and the GSK3 activity after lesion. For example, for some cultured cells, two different phases of kinase activation after MAIs exposure have been reported [11]. Thus, in contrast to GSK3β, ERK1/2 activity increased shortly after stimuli with myelin (Fig. 3). In a second phase, ERK1/2 decreased and GSK3β increased in parallel. As indicated above, ERK1/2 activity has been reported to inhibit GSK3β in hepatocellular carcinoma cells [28]. In fact, these data suggest an acute effect of ERK1/2 after stimulation with inhibitory molecules delaying the activation of GSK3β [11]. ERK1/2 activation by MAIs is mediated by fast EGF receptor (EGFR) transactivation [138]. Taking into account that, as indicated above, fast inhibition of GSK3 induces axon formation and growth (see above), it is reasonable to consider that ERK1/2 and GSK3 may act in a coordinated way to overcome from the very outset MAIs-mediated axon growth inhibition. Unfortunately, these neuronal responses are not followed up by fast regeneration and the lesioned axons remain collapsed. However, chronic inhibition of GSK3 activity by meningoglial scar molecules, such as CSPG, or secreted semaphorins impairing axonal growth may contribute to the axonal sprouting observed in chronic lesions [139, 140].

Proposed roles of GSK3β in axonal outgrowth inhibition. Alabed et al. demonstrate a scenario in which activation of the Nogo receptor complex or other receptors by myelin debris or other inhibitory molecules present at the injury site (e.g., CSPGs) triggers a downstream signaling pathway that leads to the phosphorylation of the GSK3β Ser9 epitope (1) and RhoA/ROCK pathway activation (2). This Ser9 phosphorylation results in changes in GSK3β activity, which switches from baseline activity to inactive status (1). The decreased activity may also direct CRMP4 dephosphorylation (3) and binding with RhoA (4). All these effects lead to modifications of the Actin or Microtubular cytoskeleton in the axon (5) and axonal outgrowth inhibition. Grey arrow represents the pathway direction without inhibitory stimulus that may lead to increased CRMP4 phosphorylation and axonal outgrowth stimulation. In these effects, the use of CRMP4-RhoA interfering peptide (C4RIP) (6) may promote increased neurite length, and the use of inhibitors (e.g., CT99021, SB216763, and SB415286), or inhibitory constructs like GSK3βs9A against GSK3β may lead to an inhibitory axonal outgrowth effect (7). Moreover, we and others (see Refs. [11, 138]) indicate that ERK1/2 is activated through a transactivational mechanism together with the RhoA pathway after inhibitory stimulus (8). In contrast current data suggest that ERK1/2 activity leads to an initial decrease in GSK3β activity that becomes stimulated in a second phase (9). Therefore, GSK3β may act directly on the microtubule binding proteins (e.g., MAP1B and Tau) (10) and through CRMP2 (11), contributing to microtubule destabilization. In parallel, RhoA (12) may facilitate CRMP2 phosphorylation by activation through GSK3β also acting on the F-Actin
GSK3β Inhibition as a Strategy to Overcome Axon Regeneration: Lessons from In Vivo and In Vitro Models
The first experiments aimed to enhance axon regeneration by inhibiting GSK3β activity were carried out using LiCl as inhibitor [141–144]. LiCl seems to be a nonspecific GSK3β inhibitor [145] and it may affect other kinases and signaling pathways after treatment (see also Ref. [146]). In this respect, other highly selective GSK3β inhibitors, such as SB-415286 and SB-216763, have been developed to avoid these indirect effects [147–149]. Moreover, other groups of molecules, such as aminopyrimidines (purine analogues) like CT99021, seem to inhibit GSK3β in a nanomolar concentration range [150, 151]. Another approach was to combine treatment with LiCl and chondroitinase ABC [152] to induce ganglion cell axon regeneration in the retina and the rubrospinal tract after lesion [10, 152]. Therefore, the question concerning treatment with LiCl is whether these results are dependent on the effect of LiCl on GSK3β or on other kinases and/or factors that promote cell survival (see also Refs. [152–155]; Fig. 3). To determine the functional effects of GSK3β inhibition after in vivo spinal cord lesion, Dill et al. [10] compared systemic treatment of lesioned mice with LiCl or SB-415286 induced sprouting and axonal growth of the CST and raphe spinal axons, specifically in the dorsal transection and contusion models, which correlated with an improvement in behavioral recovery. Using in vitro models, our previous studies in organotypic slice co-cultures revealed that GSK3β remains active for several days post-lesion (Fig. 3) [59]. Treatment of the axotomized slices with GSK3β inhibitors (SB-415286 or SB-216763) promoted highly significant axonal growth and sprouting across the lesion [11]. Taken together, these data point to GSK3β activity as a therapeutic target to improve axon regeneration. However, our studies developed using NgR1 (NgR1 o/o) knockout slices also support the role of other inhibitory molecules, such as semaphorins and CSPGs and their receptors, in GSK3β activation in lesioned neurons (e.g., Refs. [59, 108, 156], see above for details; Fig. 3). In contrast to other reports, in our experiments MAIs and Sema3A stimulation enhanced Tau phosphorylation mediated by GSK3β, leading to cytoskeleton instability, neurite retraction, and growth cone collapse (e.g., Refs. [157, 158]; Fig. 3).
Recently, some groups have developed transgenic mice in an attempt to improve axon regeneration after injury and to better understand the role of GSK3 kinases in this scenario. However, deletion of the GSK3β gene in mice is lethal [159, 160]. Only GSK3β heterozygous mice survive, but they show a wide range of neurological defects [161, 162]. In contrast, it seems that mice lacking GSK3α are viable and relatively normal [163]. Thus, new studies using the recently developed conditional mice (see above) and the GSK3α knockouts will determine the potential of GSK3 and particularly GSK3β as targets after spinal cord lesion and its associated muscle dystrophy.
Perspectives
A complete understanding of the role of GSK3β after neural lesions requires additional experiments to be fully revealed, since discrepancies exist in the literature. However, recent biochemical results support histological data indicating that, after lesion, axons might try to regenerate shortly after lesion inhibiting GSK3β through increased ERK1/2 activity. However, due to the inhibitory environment, there is a progressive increase in GSK3β activity after injury that affects neurite outgrowth, but this might be involved in the axonal sprouting/branching observed in chronic lesions. However, due to the different experimental models, it would be of great interest to study the status and participation of the regulatory pathways of GSK3β activity after spinal cord lesion in newly generated mice with controlled expression of GSK3β in neurons. In fact, it seems reasonable to consider the possibility that some of the discrepancies observed between the different in vitro and in vivo studies may be associated with different experimental conditions or different lesion models, respectively. However, we also cannot rule out the presence of differentially regulated GSK3β in different domains of the developing and regenerating neuron that may affect the observed results. Indeed, we feel that analysis of the phosphorylation status of this kinase as a whole does not help to reveal its specific role in particular conditions. In this scenario, mice that may avoid unwanted effects of prolonged deletion of PI3K/Akt/mTOR or GSK3β are essential in helping unravel these challenging questions.
Abbreviations
- APC:
-
Adenomatous polyposis coli
- BDNF:
-
Brain-derived neurothrophic factor
- Cdk5:
-
Cyclin-dependent kinase 5
- CGN:
-
Cerebellar granule neuron
- CLASP2:
-
Cytoplasmic linker proteins (CLIP-associate protein)
- CRMP:
-
Collapsing response mediator protein
- CSPG:
-
Chondroitin sulfate proteoglycan
- CST:
-
Corticospinal tract
- DRG:
-
Dorsal root ganglion
- DS:
-
Dermatan
- EGF:
-
Epidermal growth factor
- FGF:
-
Fibroblast growth factor
- GAG:
-
Glycosaminoglycan
- GSK3β:
-
Glycogen synthase kinase 3 beta
- HS:
-
Heparan sulfate
- ILK:
-
Integrin-linked kinase
- KS:
-
Keratan sulfate
- MAIs:
-
Myelin-associated inhibitors
- MAG:
-
Myelin-associated glycoprotein
- MAPs:
-
Microtubule-associated proteins
- MAPK:
-
Mitogen-activated protein kinase
- MTOC:
-
Microtubule organization center
- NgR1:
-
Nogo receptor 1
- NGF:
-
Neural growth factor
- Np:
-
Neuropilin
- NT-3:
-
Neurotrophine-3
- Olfm1:
-
Olfactomedin1
- OMgp:
-
Oligodendrocyte myelin glycoprotein
- PirB:
-
Paired immunoglobuline-like receptor B
- PI3K:
-
Phosphoinositide 3-kinase
- PKA:
-
Protein kinase A
- Plex:
-
Plexin
- PTEN:
-
Phosphatase and tensin homolog
- PTPase:
-
Protein tyrosine phosphatase
- PTPσ:
-
Protein phosphatase 3
- TOR:
-
Target of rapamycin
- VEGFr:
-
Vascular endothelial growth factor receptor
- ZAK1:
-
Zipper sterile-motif kinase
References
Hernandez F, de Barreda EG, Fuster-Matanzo A, Goni-Oliver P, Lucas JJ, Avila J (2009) The role of GSK3 in Alzheimer disease. Brain Res Bull 80(4–5):248–250
Emamian ES (2012) AKT/GSK3 signaling pathway and schizophrenia. Front Mol Neurosci 5:33
Cole AR (2013) Glycogen synthase kinase 3 substrates in mood disorders and schizophrenia. FEBS J. doi:10.1111/febs.12407
Latapy C, Rioux V, Guitton MJ, Beaulieu JM (2012) Selective deletion of forebrain glycogen synthase kinase 3beta reveals a central role in serotonin-sensitive anxiety and social behaviour. Philos Trans R Soc Lond B Biol Sci 367(1601):2460–2474
Beaulieu JM, Zhang X, Rodriguiz RM, Sotnikova TD, Cools MJ, Wetsel WC, Gainetdinov RR, Caron MG (2008) Role of GSK3 beta in behavioral abnormalities induced by serotonin deficiency. Proc Natl Acad Sci U S A 105(4):1333–1338
Hur EM, Zhou FQ (2010) GSK3 signalling in neural development. Nat Rev Neurosci 11(8):539–551
Kim YT, Hur EM, Snider WD, Zhou FQ (2011) Role of GSK3 signaling in neuronal morphogenesis. Front Mol Neurosci 4:48
Liu CM, Hur EM, Zhou FQ (2012) Coordinating gene expression and axon assembly to control axon growth: potential role of GSK3 signaling. Front Mol Neurosci 5:3
Alabed YZ, Pool M, Ong Tone S, Sutherland C, Fournier AE (2010) GSK3 beta regulates myelin-dependent axon outgrowth inhibition through CRMP4. J Neurosci 30(16):5635–5643
Dill J, Wang H, Zhou F, Li S (2008) Inactivation of glycogen synthase kinase 3 promotes axonal growth and recovery in the CNS. J Neurosci 28(36):8914–8928
Seira O, Gavin R, Gil V, Llorens F, Rangel A, Soriano E, del Rio JA (2010) Neurites regrowth of cortical neurons by GSK3beta inhibition independently of Nogo receptor 1. J Neurochem 113(6):1644–1658
Liu K, Lu Y, Lee JK, Samara R, Willenberg R, Sears-Kraxberger I, Tedeschi A, Park KK, Jin D, Cai B, Xu B, Connolly L, Steward O, Zheng B, He Z (2010) PTEN deletion enhances the regenerative ability of adult corticospinal neurons. Nat Neurosci 13(9):1075–1081
Arevalo MA, Rodriguez-Tebar A (2006) Activation of casein kinase II and inhibition of phosphatase and tensin homologue deleted on chromosome 10 phosphatase by nerve growth factor/p75NTR inhibit glycogen synthase kinase-3beta and stimulate axonal growth. Mol Biol Cell 17(8):3369–3377
Beurel E, Michalek SM, Jope RS (2010) Innate and adaptive immune responses regulated by glycogen synthase kinase-3 (GSK3). Trends Immunol 31(1):24–31
Lee J, Kim MS (2007) The role of GSK3 in glucose homeostasis and the development of insulin resistance. Diabetes Res Clin Pract 77(Suppl 1):S49–S57
Cohen P, Goedert M (2004) GSK3 inhibitors: development and therapeutic potential. Nat Rev Drug Discov 3(6):479–487
Woodgett JR, Cohen P (1984) Multisite phosphorylation of glycogen synthase. Molecular basis for the substrate specificity of glycogen synthase kinase-3 and casein kinase-II (glycogen synthase kinase-5). Biochim Biophys Acta 788(3):339–347
Wang Y, Roach PJ (1993) Inactivation of rabbit muscle glycogen synthase by glycogen synthase kinase-3. Dominant role of the phosphorylation of Ser-640 (site-3a). J Biol Chem 268(32):23876–23880
Ali A, Hoeflich KP, Woodgett JR (2001) Glycogen synthase kinase-3: properties, functions, and regulation. Chem Rev 101(8):2527–2540
Cohen P, Frame S (2001) The renaissance of GSK3. Nat Rev Mol Cell Biol 2(10):769–776
Maurin H, Lechat B, Dewachter I, Ris L, Louis JV, Borghgraef P, Devijver H, Jaworski T, Van Leuven F (2013) Neurological characterization of mice deficient in GSK3alpha highlight pleiotropic physiological functions in cognition and pathological activity as Tau kinase. Mol Brain 6:27
Cho JH, Johnson GV (2003) Glycogen synthase kinase 3beta phosphorylates tau at both primed and unprimed sites. Differential impact on microtubule binding. J Biol Chem 278(1):187–193
Lau KF, Miller CC, Anderton BH, Shaw PC (1999) Expression analysis of glycogen synthase kinase-3 in human tissues. J Pept Res 54(1):85–91
Mukai F, Ishiguro K, Sano Y, Fujita SC (2002) Alternative splicing isoform of tau protein kinase I/glycogen synthase kinase 3beta. J Neurochem 81(5):1073–1083
Takahashi M, Tomizawa K, Kato R, Sato K, Uchida T, Fujita SC, Imahori K (1994) Localization and developmental changes of tau protein kinase I/glycogen synthase kinase-3 beta in rat brain. J Neurochem 63(1):245–255
Wang QM, Fiol CJ, DePaoli-Roach AA, Roach PJ (1994) Glycogen synthase kinase-3 beta is a dual specificity kinase differentially regulated by tyrosine and serine/threonine phosphorylation. J Biol Chem 269(20):14566–14574
Grimes CA, Jope RS (2001) The multifaceted roles of glycogen synthase kinase 3beta in cellular signaling. Prog Neurobiol 65(4):391–426
Ding Q, Xia W, Liu JC, Yang JY, Lee DF, Xia J, Bartholomeusz G, Li Y, Pan Y, Li Z, Bargou RC, Qin J, Lai CC, Tsai FJ, Tsai CH, Hung MC (2005) Erk associates with and primes GSK-3beta for its inactivation resulting in upregulation of beta-catenin. Mol Cell 19(2):159–170
Cross DA, Alessi DR, Cohen P, Andjelkovich M, Hemmings BA (1995) Inhibition of glycogen synthase kinase-3 by insulin mediated by protein kinase B. Nature 378(6559):785–789
Bhat RV, Shanley J, Correll MP, Fieles WE, Keith RA, Scott CW, Lee CM (2000) Regulation and localization of tyrosine216 phosphorylation of glycogen synthase kinase-3beta in cellular and animal models of neuronal degeneration. Proc Natl Acad Sci U S A 97(20):11074–11079
Lebel M, Patenaude C, Allyson J, Massicotte G, Cyr M (2009) Dopamine D1 receptor activation induces tau phosphorylation via cdk5 and GSK3 signaling pathways. Neuropharmacology 57(4):392–402
Shelly M, Lim BK, Cancedda L, Heilshorn SC, Gao H, Poo MM (2010) Local and long-range reciprocal regulation of cAMP and cGMP in axon/dendrite formation. Science 327(5965):547–552
Li L, Yuan H, Weaver CD, Mao J, Farr GH 3rd, Sussman DJ, Jonkers J, Kimelman D, Wu D (1999) Axin and Frat1 interact with dvl and GSK, bridging Dvl to GSK in Wnt-mediated regulation of LEF-1. EMBO J 18(15):4233–4240
Freemantle SJ, Portland HB, Ewings K, Dmitrovsky F, DiPetrillo K, Spinella MJ, Dmitrovsky E (2002) Characterization and tissue-specific expression of human GSK-3-binding proteins FRAT1 and FRAT2. Gene 291(1–2):17–27
Zhou BP, Deng J, Xia W, Xu J, Li YM, Gunduz M, Hung MC (2004) Dual regulation of Snail by GSK-3beta-mediated phosphorylation in control of epithelial-mesenchymal transition. Nat Cell Biol 6(10):931–940
Doble BW, Woodgett JR (2003) GSK-3: tricks of the trade for a multi-tasking kinase. J Cell Sci 116(Pt 7):1175–1186
Jope RS, Johnson GV (2004) The glamour and gloom of glycogen synthase kinase-3. Trends Biochem Sci 29(2):95–102
Chenn A, Walsh CA (2002) Regulation of cerebral cortical size by control of cell cycle exit in neural precursors. Science 297(5580):365–369
Machold R, Hayashi S, Rutlin M, Muzumdar MD, Nery S, Corbin JG, Gritli-Linde A, Dellovade T, Porter JA, Rubin LL, Dudek H, McMahon AP, Fishell G (2003) Sonic hedgehog is required for progenitor cell maintenance in telencephalic stem cell niches. Neuron 39(6):937–950
Tahirovic S, Bradke F (2009) Neuronal polarity. Cold Spring Harb Perspect Biol 1(3):a001644
Horiguchi K, Hanada T, Fukui Y, Chishti AH (2006) Transport of PIP3 by GAKIN, a kinesin-3 family protein, regulates neuronal cell polarity. J Cell Biol 174(3):425–436
Biernat J, Wu YZ, Timm T, Zheng-Fischhofer Q, Mandelkow E, Meijer L, Mandelkow EM (2002) Protein kinase MARK/PAR-1 is required for neurite outgrowth and establishment of neuronal polarity. Mol Biol Cell 13(11):4013–4028
Barnes AP, Lilley BN, Pan YA, Plummer LJ, Powell AW, Raines AN, Sanes JR, Polleux F (2007) LKB1 and SAD kinases define a pathway required for the polarization of cortical neurons. Cell 129(3):549–563
Yoshimura T, Kawano Y, Arimura N, Kawabata S, Kikuchi A, Kaibuchi K (2005) GSK-3beta regulates phosphorylation of CRMP-2 and neuronal polarity. Cell 120(1):137–149
Zhou FQ, Zhou J, Dedhar S, Wu YH, Snider WD (2004) NGF-induced axon growth is mediated by localized inactivation of GSK-3beta and functions of the microtubule plus end binding protein APC. Neuron 42(6):897–912
Jiang H, Guo W, Liang X, Rao Y (2005) Both the establishment and the maintenance of neuronal polarity require active mechanisms: critical roles of GSK-3beta and its upstream regulators. Cell 120(1):123–135
Jones DM, Tucker BA, Rahimtula M, Mearow KM (2003) The synergistic effects of NGF and IGF-1 on neurite growth in adult sensory neurons: convergence on the PI 3-kinase signaling pathway. J Neurochem 86(5):1116–1128
Garrido JJ, Simon D, Varea O, Wandosell F (2007) GSK3 alpha and GSK3 beta are necessary for axon formation. FEBS Lett 581(8):1579–1586
Sanchez S, Sayas CL, Lim F, Diaz-Nido J, Avila J, Wandosell F (2001) The inhibition of phosphatidylinositol-3-kinase induces neurite retraction and activates GSK3. J Neurochem 78(3):468–481
Farghaian H, Turnley AM, Sutherland C, Cole AR (2011) Bioinformatic prediction and confirmation of beta-adducin as a novel substrate of glycogen synthase kinase 3. J Biol Chem 286(28):25274–25283
Cole AR, Knebel A, Morrice NA, Robertson LA, Irving AJ, Connolly CN, Sutherland C (2004) GSK-3 phosphorylation of the Alzheimer epitope within collapsing response mediator proteins regulates axon elongation in primary neurons. J Biol Chem 279(48):50176–50180
Naska S, Park KJ, Hannigan GE, Dedhar S, Miller FD, Kaplan DR (2006) An essential role for the integrin-linked kinase-glycogen synthase kinase-3 beta pathway during dendrite initiation and growth. J Neurosci 26(51):13344–13356
Owen R, Gordon-Weeks PR (2003) Inhibition of glycogen synthase kinase 3beta in sensory neurons in culture alters filopodia dynamics and microtubule distribution in growth cones. Mol Cell Neurosci 23(4):626–637
Kim WY, Zhou FQ, Zhou J, Yokota Y, Wang YM, Yoshimura T, Kaibuchi K, Woodgett JR, Anton ES, Snider WD (2006) Essential roles for GSK-3s and GSK-3-primed substrates in neurotrophin-induced and hippocampal axon growth. Neuron 52(6):981–996
Taelman VF, Dobrowolski R, Plouhinec JL, Fuentealba LC, Vorwald PP, Gumper I, Sabatini DD, De Robertis EM (2010) Wnt signaling requires sequestration of glycogen synthase kinase 3 inside multivesicular endosomes. Cell 143(7):1136–1148
Llorens-Martin M, Fuster-Matanzo A, Teixeira CM, Jurado-Arjona J, Ulloa F, Defelipe J, Rabano A, Hernandez F, Soriano E, Avila J (2013) GSK-3beta overexpression causes reversible alterations on postsynaptic densities and dendritic morphology of hippocampal granule neurons in vivo. Mol Psychiatry 18(4):451–460. doi:10.1038/mp.2013.4
Moon MH, Jeong JK, Lee YJ, Seol JW, Jackson CJ, Park SY (2013) SIRT1, a class III histone deacetylase, regulates TNF-alpha-induced inflammation in human chondrocytes. Osteoarthritis Cartilage 21(3):470–480
Lohof AM, Quillan M, Dan Y, Poo MM (1992) Asymmetric modulation of cytosolic cAMP activity induces growth cone turning. J Neurosci 12(4):1253–1261
Montolio M, Messeguer J, Masip I, Guijarro P, Gavin R, Antonio Del Rio J, Messeguer A, Soriano E (2009) A semaphorin 3A inhibitor blocks axonal chemorepulsion and enhances axon regeneration. Chem Biol 16(7):691–701
Eickholt BJ, Walsh FS, Doherty P (2002) An inactive pool of GSK-3 at the leading edge of growth cones is implicated in Semaphorin 3A signaling. J Cell Biol 157(2):211–217
Chadborn NH, Ahmed AI, Holt MR, Prinjha R, Dunn GA, Jones GE, Eickholt BJ (2006) PTEN couples Sema3A signalling to growth cone collapse. J Cell Sci 119(Pt 5):951–957
Manns RP, Cook GM, Holt CE, Keynes RJ (2012) Differing semaphorin 3A concentrations trigger distinct signaling mechanisms in growth cone collapse. J Neurosci 32(25):8554–8559
Bhattacharjee N, Li N, Keenan TM, Folch A (2010) A neuron-benign microfluidic gradient generator for studying the response of mammalian neurons towards axon guidance factors. Integr Biol (Camb) 2(11–12):669–679
Del Rio JA, Gonzalez-Billault C, Urena JM, Jimenez EM, Barallobre MJ, Pascual M, Pujadas L, Simo S, La Torre A, Wandosell F, Avila J, Soriano E (2004) MAP1B is required for Netrin 1 signaling in neuronal migration and axonal guidance. Curr Biol 14(10):840–850
Barallobre MJ, Pascual M, Del Rio JA, Soriano E (2005) The Netrin family of guidance factors: emphasis on Netrin-1 signalling. Brain Res Brain Res Rev 49(1):22–47
Hur EM, Saijilafu LBD, Kim SJ, Xu WL, Zhou FQ (2011) GSK3 controls axon growth via CLASP-mediated regulation of growth cone microtubules. Genes Dev 25(18):1968–1981
Schwab ME, Bartholdi D (1996) Degeneration and regeneration of axons in the lesioned spinal cord. Physiol Rev 76(2):319–370
Horner PJ, Gage FH (2000) Regenerating the damaged central nervous system. Nature 407(6807):963–970
Llorens F, Gil V, del Rio JA (2011) Emerging functions of myelin-associated proteins during development, neuronal plasticity, and neurodegeneration. FASEB J 25(2):463–475
Rolls A, Shechter R, Schwartz M (2009) The bright side of the glial scar in CNS repair. Nat Rev Neurosci 10(3):235–241
Fitch MT, Silver J (2008) CNS injury, glial scars, and inflammation: inhibitory extracellular matrices and regeneration failure. Exp Neurol 209(2):294–301
Silver J, Miller JH (2004) Regeneration beyond the glial scar. Nat Rev Neurosci 5(2):146–156
Afshari FT, Kappagantula S, Fawcett JW (2009) Extrinsic and intrinsic factors controlling axonal regeneration after spinal cord injury. Expert Rev Mol Med 11:e37
Busch SA, Silver J (2007) The role of extracellular matrix in CNS regeneration. Curr Opin Neurobiol 17(1):120–127
Tan AM, Zhang W, Levine JM (2005) NG2: a component of the glial scar that inhibits axon growth. J Anat 207(6):717–725
Pasterkamp RJ, Verhaagen J (2006) Semaphorins in axon regeneration: developmental guidance molecules gone wrong? Philos Trans R Soc Lond B Biol Sci 361(1473):1499–1511
Low K, Culbertson M, Bradke F, Tessier-Lavigne M, Tuszynski MH (2008) Netrin-1 is a novel myelin-associated inhibitor to axon growth. J Neurosci 28(5):1099–1108
Kolodkin AL, Tessier-Lavigne M (2011) Mechanisms and molecules of neuronal wiring: a primer. Cold Spring Harb Perspect Biol 3(6). doi:10.1101/cshperspect.a001727
Mann F, Rougon G (2007) Mechanisms of axon guidance: membrane dynamics and axonal transport in semaphorin signalling. J Neurochem 102(2):316–323
Pasterkamp RJ (2012) Getting neural circuits into shape with semaphorins. Nat Rev Neurosci 13(9):605–618
Kury P, Abankwa D, Kruse F, Greiner-Petter R, Muller HW (2004) Gene expression profiling reveals multiple novel intrinsic and extrinsic factors associated with axonal regeneration failure. Eur J Neurosci 19(1):32–42
Burgaya F, Fontana X, Martinez A, Montolio M, Mingorance A, Simo S, del Rio JA, Soriano E (2006) Semaphorin 6C leads to GSK-3-dependent growth cone collapse and redistributes after entorhino-hippocampal axotomy. Mol Cell Neurosci 33(3):321–334
Oschipok LW, Teh J, McPhail LT, Tetzlaff W (2008) Expression of Semaphorin3C in axotomized rodent facial and rubrospinal neurons. Neurosci Lett 434(1):113–118
Niclou SP, Franssen EH, Ehlert EM, Taniguchi M, Verhaagen J (2003) Meningeal cell-derived semaphorin 3A inhibits neurite outgrowth. Mol Cell Neurosci 24(4):902–912
Hida T, Yamashita N, Usui H, Nakamura F, Sasaki Y, Kikuchi A, Goshima Y (2012) GSK3beta/axin-1/beta-catenin complex is involved in semaphorin3A signaling. J Neurosci 32(35):11905–11918
Moreau-Fauvarque C, Kumanogoh A, Camand E, Jaillard C, Barbin G, Boquet I, Love C, Jones EY, Kikutani H, Lubetzki C, Dusart I, Chedotal A (2003) The transmembrane semaphorin Sema4D/CD100, an inhibitor of axonal growth, is expressed on oligodendrocytes and upregulated after CNS lesion. J Neurosci 23(27):9229–9239
Pasterkamp RJ, Kolk SM, Hellemons AJ, Kolodkin AL (2007) Expression patterns of semaphorin7A and plexinC1 during rat neural development suggest roles in axon guidance and neuronal migration. BMC Dev Biol 7:98
Shim SO, Cafferty WB, Schmidt EC, Kim BG, Fujisawa H, Strittmatter SM (2012) PlexinA2 limits recovery from corticospinal axotomy by mediating oligodendrocyte-derived Sema6A growth inhibition. Mol Cell Neurosci 50(2):193–200
Gallo V, Bertolotto A (1990) Extracellular matrix of cultured glial cells: selective expression of chondroitin 4-sulfate by type-2 astrocytes and their progenitors. Exp Cell Res 187(2):211–223
Gallo V, Bertolotto A, Levi G (1987) The proteoglycan chondroitin sulfate is present in a subpopulation of cultured astrocytes and in their precursors. Dev Biol 123(1):282–285
Hirsch S, Bahr M (1999) Immunocytochemical characterization of reactive optic nerve astrocytes and meningeal cells. Glia 26(1):36–46
Junghans U, Koops A, Westmeyer A, Kappler J, Meyer HE, Muller HW (1995) Purification of a meningeal cell-derived chondroitin sulphate proteoglycan with neurotrophic activity for brain neurons and its identification as biglycan. Eur J Neurosci 7(11):2341–2350
Levine JM (1994) Increased expression of the NG2 chondroitin-sulfate proteoglycan after brain injury. J Neurosci 14(8):4716–4730
Jones LL, Tuszynski MH (2002) Spinal cord injury elicits expression of keratan sulfate proteoglycans by macrophages, reactive microglia, and oligodendrocyte progenitors. J Neurosci 22(11):4611–4624
Zimmer G, Schanuel SM, Burger S, Weth F, Steinecke A, Bolz J, Lent R (2010) Chondroitin sulfate acts in concert with semaphorin 3A to guide tangential migration of cortical interneurons in the ventral telencephalon. Cereb Cortex 20(10):2411–2422
Lips K, Stichel CC, Muller HW (1995) Restricted appearance of tenascin and chondroitin sulphate proteoglycans after transection and sprouting of adult rat postcommissural fornix. J Neurocytol 24(6):449–464
McKeon RJ, Jurynec MJ, Buck CR (1999) The chondroitin sulfate proteoglycans neurocan and phosphacan are expressed by reactive astrocytes in the chronic CNS glial scar. J Neurosci 19(24):10778–10788
Haas CA, Rauch U, Thon N, Merten T, Deller T (1999) Entorhinal cortex lesion in adult rats induces the expression of the neuronal chondroitin sulfate proteoglycan neurocan in reactive astrocytes. J Neurosci 19(22):9953–9963
McKeon RJ, Schreiber RC, Rudge JS, Silver J (1991) Reduction of neurite outgrowth in a model of glial scarring following CNS injury is correlated with the expression of inhibitory molecules on reactive astrocytes. J Neurosci 11(11):3398–3411
Asher RA, Morgenstern DA, Shearer MC, Adcock KH, Pesheva P, Fawcett JW (2002) Versican is upregulated in CNS injury and is a product of oligodendrocyte lineage cells. J Neurosci 22(6):2225–2236
Bovolenta P, Wandosell F, Nieto-Sampedro M (1993) Characterization of a neurite outgrowth inhibitor expressed after CNS injury. Eur J Neurosci 5(5):454–465
Oohira A, Matsui F, Katoh-Semba R (1991) Inhibitory effects of brain chondroitin sulfate proteoglycans on neurite outgrowth from PC12D cells. J Neurosci 11(3):822–827
Ughrin YM, Chen ZJ, Levine JM (2003) Multiple regions of the NG2 proteoglycan inhibit neurite growth and induce growth cone collapse. J Neurosci 23(1):175–186
Shen Y, Tenney AP, Busch SA, Horn KP, Cuascut FX, Liu K, He Z, Silver J, Flanagan JG (2009) PTPsigma is a receptor for chondroitin sulfate proteoglycan, an inhibitor of neural regeneration. Science 326(5952):592–596
Coles CH, Shen Y, Tenney AP, Siebold C, Sutton GC, Lu W, Gallagher JT, Jones EY, Flanagan JG, Aricescu AR (2011) Proteoglycan-specific molecular switch for RPTPsigma clustering and neuronal extension. Science 332(6028):484–488
Duan Y, Giger RJ (2010) A new role for RPTPsigma in spinal cord injury: signaling chondroitin sulfate proteoglycan inhibition. Sci Signal 3(110):pe6
Fry EJ, Chagnon MJ, Lopez-Vales R, Tremblay ML, David S (2010) Corticospinal tract regeneration after spinal cord injury in receptor protein tyrosine phosphatase sigma deficient mice. Glia 58(4):423–433
Fisher D, Xing B, Dill J, Li H, Hoang HH, Zhao Z, Yang XL, Bachoo R, Cannon S, Longo FM, Sheng M, Silver J, Li S (2011) Leukocyte common antigen-related phosphatase is a functional receptor for chondroitin sulfate proteoglycan axon growth inhibitors. J Neurosci 31(40):14051–14066
Mingorance A, Fontana X, Sole M, Burgaya F, Urena JM, Teng FY, Tang BL, Hunt D, Anderson PN, Bethea JR, Schwab ME, Soriano E, del Rio JA (2004) Regulation of Nogo and Nogo receptor during the development of the entorhino-hippocampal pathway and after adult hippocampal lesions. Mol Cell Neurosci 26(1):34–49
Hunt D, Coffin RS, Anderson PN (2002) The Nogo receptor, its ligands and axonal regeneration in the spinal cord; a review. J Neurocytol 31(2):93–120
Meier S, Brauer AU, Heimrich B, Schwab ME, Nitsch R, Savaskan NE (2003) Molecular analysis of Nogo expression in the hippocampus during development and following lesion and seizure. FASEB J 17(9):1153–1155
Cao Z, Gao Y, Deng K, Williams G, Doherty P, Walsh FS (2010) Receptors for myelin inhibitors: structures and therapeutic opportunities. Mol Cell Neurosci 43(1):1–14
Giger RJ, Venkatesh K, Chivatakarn O, Raiker SJ, Robak L, Hofer T, Lee H, Rader C (2008) Mechanisms of CNS myelin inhibition: evidence for distinct and neuronal cell type specific receptor systems. Restor Neurol Neurosci 26(2–3):97–115
Fawcett JW, Schwab ME, Montani L, Brazda N, Muller HW (2012) Defeating inhibition of regeneration by scar and myelin components. Handb Clin Neurol 109:503–522
Thomas R, Favell K, Morante-Redolat J, Pool M, Kent C, Wright M, Daignault K, Ferraro GB, Montcalm S, Durocher Y, Fournier A, Perez-Tur J, Barker PA (2010) LGI1 is a Nogo receptor 1 ligand that antagonizes myelin-based growth inhibition. J Neurosci 30(19):6607–6612
Zhang L, Zheng S, Wu H, Wu Y, Liu S, Fan M, Zhang J (2009) Identification of BLyS (B lymphocyte stimulator), a non-myelin-associated protein, as a functional ligand for Nogo-66 receptor. J Neurosci 29(19):6348–6352
Lee H, Raiker SJ, Venkatesh K, Geary R, Robak LA, Zhang Y, Yeh HH, Shrager P, Giger RJ (2008) Synaptic function for the Nogo-66 receptor NgR1: regulation of dendritic spine morphology and activity-dependent synaptic strength. J Neurosci 28(11):2753–2765
Nakaya N, Sultana A, Lee HS, Tomarev SI (2012) Olfactomedin 1 interacts with the Nogo A receptor complex to regulate axon growth. J Biol Chem 287(44):37171–37184
Sato Y, Iketani M, Kurihara Y, Yamaguchi M, Yamashita N, Nakamura F, Arie Y, Kawasaki T, Hirata T, Abe T, Kiyonari H, Strittmatter SM, Goshima Y, Takei K (2011) Cartilage acidic protein-1B (LOTUS), an endogenous Nogo receptor antagonist for axon tract formation. Science 333(6043):769–773
Worter V, Schweigreiter R, Kinzel B, Mueller M, Barske C, Bock G, Frentzel S, Bandtlow CE (2009) Inhibitory activity of myelin-associated glycoprotein on sensory neurons is largely independent of NgR1 and NgR2 and resides within Ig-Like domains 4 and 5. PLoS One 4(4):e5218
Hu F, Strittmatter SM (2008) The N-terminal domain of Nogo-A inhibits cell adhesion and axonal outgrowth by an integrin-specific mechanism. J Neurosci 28(5):1262–1269
Venkatesh K, Chivatakarn O, Lee H, Joshi PS, Kantor DB, Newman BA, Mage R, Rader C, Giger RJ (2005) The Nogo-66 receptor homolog NgR2 is a sialic acid-dependent receptor selective for myelin-associated glycoprotein. J Neurosci 25(4):808–822
Dickendesher TL, Baldwin KT, Mironova YA, Koriyama Y, Raiker SJ, Askew KL, Wood A, Geoffroy CG, Zheng B, Liepmann CD, Katagiri Y, Benowitz LI, Geller HM, Giger RJ (2012) NgR1 and NgR3 are receptors for chondroitin sulfate proteoglycans. Nat Neurosci 15(5):703–712
Grunewald E, Kinnell HL, Porteous DJ, Thomson PA (2009) GPR50 interacts with neuronal NOGO-A and affects neurite outgrowth. Mol Cell Neurosci 42(4):363–371
Atwal JK, Pinkston-Gosse J, Syken J, Stawicki S, Wu Y, Shatz C, Tessier-Lavigne M (2008) PirB is a functional receptor for myelin inhibitors of axonal regeneration. Science 322(5903):967–970
Chivatakarn O, Kaneko S, He Z, Tessier-Lavigne M, Giger RJ (2007) The Nogo-66 receptor NgR1 is required only for the acute growth cone-collapsing but not the chronic growth-inhibitory actions of myelin inhibitors. J Neurosci 27(27):7117–7124
O'Brien GS, Martin SM, Sollner C, Wright GJ, Becker CG, Portera-Cailliau C, Sagasti A (2009) Developmentally regulated impediments to skin reinnervation by injured peripheral sensory axon terminals. Curr Biol 19(24):2086–2090
Atwal JK, Singh KK, Tessier-Lavigne M, Miller FD, Kaplan DR (2003) Semaphorin 3F antagonizes neurotrophin-induced phosphatidylinositol 3-kinase and mitogen-activated protein kinase kinase signaling: a mechanism for growth cone collapse. J Neurosci 23(20):7602–7609
Uchida Y, Ohshima T, Sasaki Y, Suzuki H, Yanai S, Yamashita N, Nakamura F, Takei K, Ihara Y, Mikoshiba K, Kolattukudy P, Honnorat J, Goshima Y (2005) Semaphorin3A signalling is mediated via sequential Cdk5 and GSK3beta phosphorylation of CRMP2: implication of common phosphorylating mechanism underlying axon guidance and Alzheimer's disease. Genes Cells 10(2):165–179
Uchida Y, Ohshima T, Yamashita N, Ogawara M, Sasaki Y, Nakamura F, Goshima Y (2009) Semaphorin3A signaling mediated by Fyn-dependent tyrosine phosphorylation of collapsing response mediator protein 2 at tyrosine 32. J Biol Chem 284(40):27393–27401
Gao Y, Deng K, Cao Z, Graziani EI, Gilbert AM, Koehn FE, Wood A, Doherty P, Walsh FS (2010) Amphotericin B, identified from a natural product screen, antagonizes CNS inhibitors to promote axon growth via activation of an Akt pathway in neurons. J Neurochem 113(5):1331–1342
Brajtburg J, Powderly WG, Kobayashi GS, Medoff G (1990) Amphotericin B: current understanding of mechanisms of action. Antimicrob Agents Chemother 34(2):183–188
Park KK, Liu K, Hu Y, Smith PD, Wang C, Cai B, Xu B, Connolly L, Kramvis I, Sahin M, He Z (2008) Promoting axon regeneration in the adult CNS by modulation of the PTEN/mTOR pathway. Science 322(5903):963–966
Park KK, Liu K, Hu Y, Kanter JL, He Z (2010) PTEN/mTOR and axon regeneration. Exp Neurol 223(1):45–50
Zhao T, Qi Y, Li Y, Xu K (2012) PI3 Kinase regulation of neural regeneration and muscle hypertrophy after spinal cord injury. Mol Biol Rep 39(4):3541–3547
Shen JY, Yi XX, Xiong NX, Wang HJ, Duan XW, Zhao HY (2011) GSK-3beta activation mediates Nogo-66-induced inhibition of neurite outgrowth in N2a cells. Neurosci Lett 505(2):165–170
GrandPre T, Nakamura F, Vartanian T, Strittmatter SM (2000) Identification of the Nogo inhibitor of axon regeneration as a Reticulon protein. Nature 403(6768):439–444
Koprivica V, Cho KS, Park JB, Yiu G, Atwal J, Gore B, Kim JA, Lin E, Tessier-Lavigne M, Chen DF, He Z (2005) EGFR activation mediates inhibition of axon regeneration by myelin and chondroitin sulfate proteoglycans. Science 310(5745):106–110
Navarro X, Vivo M, Valero-Cabre A (2007) Neural plasticity after peripheral nerve injury and regeneration. Prog Neurobiol 82(4):163–201
Schwab ME (1990) Myelin-associated inhibitors of neurite growth and regeneration in the CNS. Trends Neurosci 13(11):452–456
Williams R, Ryves WJ, Dalton EC, Eickholt B, Shaltiel G, Agam G, Harwood AJ (2004) A molecular cell biology of lithium. Biochem Soc Trans 32(Pt 5):799–802
Bhat RV, Budd Haeberlein SL, Avila J (2004) Glycogen synthase kinase 3: a drug target for CNS therapies. J Neurochem 89(6):1313–1317
Takahashi M, Yasutake K, Tomizawa K (1999) Lithium inhibits neurite growth and tau protein kinase I/glycogen synthase kinase-3beta-dependent phosphorylation of juvenile tau in cultured hippocampal neurons. J Neurochem 73(5):2073–2083
Goold RG, Owen R, Gordon-Weeks PR (1999) Glycogen synthase kinase 3beta phosphorylation of microtubule-associated protein 1B regulates the stability of microtubules in growth cones. J Cell Sci 112(Pt 19):3373–3384
Pardo R, Andreolotti AG, Ramos B, Picatoste F, Claro E (2003) Opposed effects of lithium on the MEK-ERK pathway in neural cells: inhibition in astrocytes and stimulation in neurons by GSK3 independent mechanisms. J Neurochem 87(2):417–426
Freland L, Beaulieu JM (2012) Inhibition of GSK3 by lithium, from single molecules to signaling networks. Front Mol Neurosci 5:14
Coghlan MP, Culbert AA, Cross DA, Corcoran SL, Yates JW, Pearce NJ, Rausch OL, Murphy GJ, Carter PS, Roxbee Cox L, Mills D, Brown MJ, Haigh D, Ward RW, Smith DG, Murray KJ, Reith AD, Holder JC (2000) Selective small molecule inhibitors of glycogen synthase kinase-3 modulate glycogen metabolism and gene transcription. Chem Biol 7(10):793–803
Smith DG, Buffet M, Fenwick AE, Haigh D, Ife RJ, Saunders M, Slingsby BP, Stacey R, Ward RW (2001) 3-Anilino-4-arylmaleimides: potent and selective inhibitors of glycogen synthase kinase-3 (GSK-3). Bioorg Med Chem Lett 11(5):635–639
Meijer L, Flajolet M, Greengard P (2004) Pharmacological inhibitors of glycogen synthase kinase 3. Trends Pharmacol Sci 25(9):471–480
Bain J, Plater L, Elliott M, Shpiro N, Hastie CJ, McLauchlan H, Klevernic I, Arthur JS, Alessi DR, Cohen P (2007) The selectivity of protein kinase inhibitors: a further update. Biochem J 408(3):297–315
Ring DB, Johnson KW, Henriksen EJ, Nuss JM, Goff D, Kinnick TR, Ma ST, Reeder JW, Samuels I, Slabiak T, Wagman AS, Hammond ME, Harrison SD (2003) Selective glycogen synthase kinase 3 inhibitors potentiate insulin activation of glucose transport and utilization in vitro and in vivo. Diabetes 52(3):588–595
Yick LW, So KF, Cheung PT, Wu WT (2004) Lithium chloride reinforces the regeneration-promoting effect of chondroitinase ABC on rubrospinal neurons after spinal cord injury. J Neurotrauma 21(7):932–943
Feinstein DL (1998) Potentiation of astroglial nitric oxide synthase type-2 expression by lithium chloride. J Neurochem 71(2):883–886
Victoratos P, Yiangou M, Avramidis N, Hadjipetrou L (1997) Regulation of cytokine gene expression by adjuvants in vivo. Clin Exp Immunol 109(3):569–578
Wada A, Yokoo H, Yanagita T, Kobayashi H (2005) Lithium: potential therapeutics against acute brain injuries and chronic neurodegenerative diseases. J Pharmacol Sci 99(4):307–321
Ahmed A, Eickholt BJ (2007) Intracellular kinases in semaphorin signaling. Adv Exp Med Biol 600:24–37
Cuchillo-Ibanez I, Seereeram A, Byers HL, Leung KY, Ward MA, Anderton BH, Hanger DP (2008) Phosphorylation of tau regulates its axonal transport by controlling its binding to kinesin. FASEB J 22(9):3186–3195
Nuydens R, Dispersyn G, de Jong M, van den Kieboom G, Borgers M, Geerts H (1997) Aberrant tau phosphorylation and neurite retraction during NGF deprivation in PC12 cells. Biochem Biophys Res Commun 240(3):687–691
Schneider MR, Lahm H, Wu M, Hoeflich A, Wolf E (2000) Transgenic mouse models for studying the functions of insulin-like growth factor-binding proteins. FASEB J 14(5):629–640
Liu KJ, Arron JR, Stankunas K, Crabtree GR, Longaker MT (2007) Chemical rescue of cleft palate and midline defects in conditional GSK-3beta mice. Nature 446(7131):79–82
Kimura T, Yamashita S, Nakao S, Park JM, Murayama M, Mizoroki T, Yoshiike Y, Sahara N, Takashima A (2008) GSK-3beta is required for memory reconsolidation in adult brain. PLoS One 3(10):e3540
O'Brien WT, Harper AD, Jove F, Woodgett JR, Maretto S, Piccolo S, Klein PS (2004) Glycogen synthase kinase-3beta haploinsufficiency mimics the behavioral and molecular effects of lithium. J Neurosci 24(30):6791–6798
MacAulay K, Doble BW, Patel S, Hansotia T, Sinclair EM, Drucker DJ, Nagy A, Woodgett JR (2007) Glycogen synthase kinase 3alpha-specific regulation of murine hepatic glycogen metabolism. Cell Metab 6(4):329–337
Acknowledgments
The authors thank V. Gil and S. Nocentini for critical reading of the manuscript and Tom Yohannan for linguistic advice. This research was supported by the Seventh Framework Programme of the European Commission, grant agreement 222887, FP7-PRIORITY, and DEMTEST (Joint Programming of Neurodegenerative Diseases), the Spanish Ministry of Science and Innovation (BFU2012-32617), the Generalitat de Catalunya (SGR2009-366), La Caixa Obra Social Foundation (LCOSF), and the Instituto Salud Carlos III (PI11/03028). OS was supported by MICINN, IBEC (Strategic Research Initiative program founded by LCOSF) and ISCIII (Ciberned).
Author information
Authors and Affiliations
Corresponding authors
Rights and permissions
About this article
Cite this article
Seira, O., del Río, J.A. Glycogen Synthase Kinase 3 Beta (GSK3β) at the Tip of Neuronal Development and Regeneration. Mol Neurobiol 49, 931–944 (2014). https://doi.org/10.1007/s12035-013-8571-y
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s12035-013-8571-y