
Abstract
The performance of tree species is influenced by environmental factors and growth stages. To evaluate the practical performance of transgenic tree species, it is insufficient to grow small, young trees under controlled conditions, such as in a growth chamber. Three transgenic Eucalyptus globulus lines, carrying the choline oxidase gene, were investigated for their salt tolerance and expression of the transgene at the young plantlet stage in a special netted-house. To clarify the characteristics at the young as well during the later stages, salt tolerance and the properties of the transgenic lines at large juvenile and adult stages were evaluated in the special netted-house. All transgenic lines showed high glycinebetaine content, particularly in young leaves. Trees of the transgenic line 107-1 showed low damage because of salinity stress based on the results from the chlorophyll analysis and malondialdehyde content, and they survived the high-salt-shock treatment at the large juvenile and adult stages. Only this line showed salt tolerance at all stages in the special netted-house. In this evaluation in the special netted-house, the tolerant line among young plantlets might perform better at all stages. Since evaluation in these special netted-house mimics field evaluation, line 107-1 is a potential tolerant line.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
Forest plantations are effective for addressing environmental problems such as desertification and global warming. The genus Eucalyptus has been used for plantations in many semi-tropical areas because of its fast growth rate and adaptability to rigorous environments [4]. Eucalyptus is also an important material for the production of fuel wood and pulp [21]. However, in some dry lands, plantations of Eucalyptus are limited by salinity [21]. To alleviate this problem, introduction of salt tolerance-inducing genes is considered as an effective method to improve the salt tolerance of plants. Recently, some studies have shown that transgenes enhance salt tolerance in woody plants [7, 8, 17, 28–30]. One of these transgenes is the choline oxidase gene (codA), which is isolated from the soil bacteria Arthrobacter globiformis [9]. The codA gene increases the accumulation of glycinebetaine by catalyzing choline in plants [6]. Glycinebetaine is a compatible solute that protects cells from salt stress by maintaining an osmotic balance and by stabilizing the three-dimensional conformation of protein complexes and the membrane [24]. The accumulation of glycinebetaine enhances salt tolerance in plants [12]. Therefore, the codA gene was introduced into Eucalyptus globulus, a species with low salt tolerance in the genus Eucalyptus [1, 18].
However, field studies of transgenic woody plants are not very advanced. In addition, the practical application of transgenic woody plants has not been established because of the lack of adequate research on transgenic woody plants grown under natural conditions [14]. Most transgenic woody plants conferring salt tolerance have demonstrated salt tolerance only under controlled conditions such as in vitro or in a growth chamber or growth room. These conditions are not same as natural environmental conditions, which are influenced by climatic factors such as temperature, light intensity, humidity, and wind. Therefore, field evaluation of salt tolerance in transgenic plants is necessary to determine their performance under natural environmental conditions. However, before field trials in Japan, an environmental biosafety risk assessment of transgenic plants in a semi-confined condition such as a special netted-house must be performed [22, 27]. On the other hand, trees such as E. globulus are perennial plants. Changes in the physiological traits might occur with growth [10]. In particular, leaf anatomy and morphology in E. globulus undergo dramatic development from the seedling stage to the adult tree stage, that is, 1–3-years old [2]. During this developmental stage, the chlorophyll content and features of the stomatal apparatus change to improve the adaptability of abiotic stresses [2, 20]. To investigate the salt tolerance of the transgenic E. globulus lines, the salt tolerance of these plants should be evaluated along with their growth.
In our previous study, the transgenic E. globulus lines 107-1, 1-9-1, and 2-1-1, which have the codA gene, showed salt tolerance at the young plantlet (ca. 10-cm height) stage with transgene expression in a special netted-house [30]. Here, to determine a tolerant line from these three transgenic lines, salt tolerance of the trees in several growth stages was evaluated in a special netted-house.
Materials and Methods
Plant Materials
Plant materials of E. globulus were propagated with cutting in vitro. Three transgenic lines (107-1, 1-9-1, and 2-1-1) carrying the choline oxidase gene (codA) [9], driven by the Eucalyptus ubiquitin promoter [30], that were isolated from the soil bacteria A. globiformis and five non-transgenic lines (No.1, M001, No.8-7, No.8-11, and No.8-20) were used as the plant material. The inside of border sequence on pGW23codA was introduced to E. globulus mediated by Agrobacterium [17]. Each unit except codA cassette was NPTII gene driven by nos promoter and GUS gene driven by GSP promoter from E. globulus. For first screening, the transformant shoots were grown on MS medium containing 100 mg/L kanamycin. Around 250 resistant buds were obtained from 10,000 explants. After 1 year cultivation, existence of NPTII gene in grown 100 shoots was confirmed by Southern blot analysis. Expression levels of codA gene and rooting ability were evaluated in easily propagated lines with less than five copies. Furthermore, after 1 year cultivation, 23 lines were obtained as rooted young plantlets. The No.1 and M001 lines were established as elite trees, and the M001 line has been shown to be salt tolerant [21]. Although transformation system in E globulus has been established [17], there are some difficulties in rooting from shoot buds and in acclimatization of small plantlets. We tried to fix the control lines among each experiment. But, it was difficult to prepare the plantlet stably due to the above reason.
The rooted young plantlets were transplanted in vitro into soil-filled quadrangular pyramid pots [30] and acclimatized in the growth room for 3 weeks; then, they were grown in a special netted-house for 2 months. The special netted-house is a special green house for the cultivation of genetically modified (GM) plants in Japan [30]. In this experiment, a special tall netted-house (10-m height) was used. The young plantlets (ca. 10 cm in height) were transplanted into a small plastic pot (φ, 14 cm; 4 L) with pre-mixed soil (Kato Inc., Kanuma, Japan). The small juvenile trees grew to ~40 cm in height within 5 months. Small juvenile trees were transferred to bigger pots as the trees grew. The 2-year-old juvenile trees in the mid-sized pots (φ, 21 cm; 15 L) were grown to ~3 m in height, but only had juvenile leaves. Adult trees (2.5-year-old) grown in large-sized pots (φ, 42 cm; 28 L) to ~4.5 m in height had adult leaves only on the treetop region. All lines were grown using tap water in the special netted-house until salinity test. All transgenic lines showed same growth profile as non-transformant except No.1 and M001 lines. Although emergence timing of adult leaves was slightly different in each line, there was no remarkable morphological difference among lines.
The internal temperature of the special netted-house was controlled by opening and closing of windows and by the gas heater. Specifically, opening and closing of roof windows was automatically activated at a threshold of 25 °C. Side windows were opened during the summer season except a strong wind. The gas heater was automatically activated when temperature fell below 15 °C [31].
Salinity Stress Tolerance Assay
Small juvenile trees, 8-month old (ca. 40 cm in height), were irrigated with 200 mL of 200 mM NaCl every 2 days for 5 weeks for the three transgenic lines (107-1, 1-9-1, and 2-1-1) and the three non-transgenic lines (No.1, M001, and No.8-7). Five trees in each line were evaluated. Salinity damage was observed at 3 and 5 weeks after starting salt-water irrigation.
Large juvenile trees, 2-year old (ca. 3 m in height), were irrigated with 2.5 L of 1.3 M NaCl once (salt-shock treatment) for the transgenic line 107-1 and the non-transgenic line No.8-8. Since salinity damage did not appear immediately, irrigation with 2 L of freshwater was performed every day. Some trees began to show some damage within 1 month. Five trees in each line were evaluated.
Adult trees (2.5 years) of the transgenic line 107-1 and the non-transgenic lines No.8-7 and No.8-20 (ca. 4.5 m in height) were treated with salinity stress in the same manner as the large juvenile trees. Starting the day after salt-water irrigation, the trees were irrigated with 2 L of fresh water every day. After 1 month, trees of the lines No.8-20 and No.107-1 did not show any salinity damage. Irrigation with salt water was repeated followed by irrigation with fresh water. Six trees from the transgenic lines and three trees from each non-transgenic lines were evaluated.
RNA Extraction and RT-PCR
Total RNA was prepared from new leaves of each line (No.1, 107-1, 1-9-1, and 2-1-1). cDNA was synthesized from the total RNA using a RNA PCR kit (TAKARA bio, Kyoto, Japan). The RT-PCR was performed using the GC-RICH PCR System (Roche, Mannheim, Germany). Specific primers for codA were the forward primer 5′-AGGAGAACTTCACCCTGCTAAC-3′ and the reverse primer 5′-AGCCGTAGTGCATCATCAGGT-3′, and the primers for E. globulusubiqitin were the forward primer 5′-CCAAGATCCAGGACAAGGAA-3′ and the reverse primer 5′-CCTCGAGCTTAATGGTCTCG-3′.
Determination of Glycinebetaine and Chlorophyll Contents
The glycinebetaine content of each line was detected from the frozen leaves by capillary electrophoresis-mass spectrometry (CE-MS) analysis [30]. The chlorophyll content was measured on the third leaf from the treetop using the chlorophyll meter SPAD-502 (Konica-Minolta, Tokyo, Japan).
Determination of Malondialdehyde Content
Malondialdehyde (MDA) content was determined by a previously described method [5]. The frozen forth leaf from the tree top was homogenized in a pre-chilled mortar with ice-cold 5 % (w/v) trichloroacetic acid. An equal volume of the extract and 0.67 % (w/v) thiobarbituric acid were mixed and heated at 100 °C for 30 min. After centrifugation (10,000×g for 1 min at 4 °C), the absorbance of the supernatants at 532 nm was read, and the values corresponding to non-specific absorption (600 nm) were subtracted. The MDA concentration was calculated by means of its molar extinction coefficient (155 mM/cm). Three trees were analyzed from each line.
Statistical Analysis
ANOVA and Tukey’s test were used by means of the SYSTAT version 11 software.
Results
Confirmation of Transgene Expression and Evaluation of Salt Tolerance of Small Juvenile Trees
The expression of the transgene was confirmed in the three transgenic lines (107-1, 1-9-1, and 2-1-1) by RT-PCR in the 8-month-old trees (ca. 40 cm in height) (Fig. 1). Furthermore, salinity stress tolerance was evaluated in these transgenic lines and three non-transgenic lines (No.1, M001, and No.8-7) in the juvenile trees. One of the non-transgenic lines M001 has been shown to be salt tolerant [21]. The trees were irrigated with salt water (200 mM NaCl) every 2 days for 5 weeks. After 3 weeks of salt-water irrigation, some trees in the lines 1-9-1, 2-1-1, and No.8-7 had started to wilt, but trees in the lines 107-1, No.1, and M001 did not show any damage by salinity stress. After 5 weeks of irrigation, one tree in the line No.107-1 and line M001 as well as two trees in the line No.1 showed salt damage on the treetop (yellow arrow in Fig. 2), but all the trees in these three lines survived (Fig. 2). On the other hand, two trees in line 2-1-1 and line No.8-7 as well as four trees in the line 1-9-1 had wilted completely (red arrow in Fig. 2). Sodium contents of leaves in E. globulus were increased with treatment of 1 % NaCl [15]. Sodium ion might be regarded as one of the causative substances to give some damage to E. globulus with NaCl treatment.

Expression analysis of the codA gene. The expression of the codA transgene in the transgenic lines (107-1, 1-9-1, and 2-1-1) was analyzed by RT-PCR. No.1 is a non-transgenic line. The expression of ubiquitin was used as the internal control

Evaluation of salt tolerance of 8-month-old trees (ca. 40 cm in height). Three individual non-transgenic lines (No.1, M001, and No.8-7) and three transgenic lines (107-1, 1-9-1, and 2-1-1) were cultivated in special netted-houses and irrigated with 200 mM NaCl for 5 weeks. The yellow arrows indicate the salt-damaged treetops. The red arrows indicate the wilted trees following the salt stress. The trees of line M001 possess high-salt tolerance compared to the other non-transgenic lines. The scale bar indicates 10 cm
Glycinebetaine Content
The glycinebetaine content was analyzed before and after salinity stress in the young and mature leaves from the 8-month-old trees. Young leaves from trees in all the lines had higher glycinebetaine content. In the young leaves before exposure to salinity stress, glycinebetaine levels were almost the same (<0.6 nmol/mg DW) except in the leaves from the line 107-1, which were 3-times higher than that in the other lines (Fig. 3). After exposure to the stress, the glycinebetaine content in all non-transgenic lines was almost the same as that before exposure to the stress, but the levels in the transgenic lines increased to >1.5 nmol/mg DW (Fig. 3). On the other hand, in mature leaves, the glycinebetaine content in all the lines increased with salt stress, but the levels did not reach >1.0 nmol/mg DW (Fig. 3).

Glycinebetaine content in leaves. Glycinebetaine levels were detected in the young and mature leaves from three non-transgenic lines (NT) No.1, M001, and No.8-7 (white) and three transgenic lines (codA) 107-1, 1-9-1, and 2-1-1 (black) before and after irrigation with 200 mM NaCl. Young leaves were defined as the leaf during expansion in the treetop region. Adult leaves were defined as the leaf that expanded completely in subalternate branch. DW denotes the dry weight. Roman letters on the graph bars indicate significant differences at 0.05 confidence level (Tukey’s multiple comparison test). The white bars are non-transgenic genotypes. The black bars represent core-type transgenic genotypes. The number in the bar indicates the number of individuals that have been measured in each genotype. The error bar indicates standard error
Damage Evaluation by Salinity Stress Treatment
The chlorophyll content in Eucalyptus leaves is reduced by salinity stress [3, 19]. Changes in the chlorophyll content before and after exposure to salinity stress is an indicator of the extent of salt tolerance. When evaluating salt tolerance of small juvenile trees, young treetop regions showed clear damage. Thus, chlorophyll content in the top leaves of the surviving trees was measured. The SPAD values of all lines were 25–30 before stress treatment, but the values decreased after salt treatment. The chlorophyll content was reduced to 58–72 % and 78–88 % in the non-transgenic and transgenic lines, respectively (Table 1). Reduction in chlorophyll content seemed to be diminished in the transgenic lines. No significant difference was noted in the chlorophyll content of the line 107-1 before and after the stress treatment (Table 1).
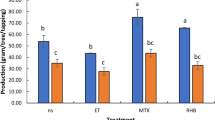
On the other hand, salinity stress has been shown to induce the accumulation of reactive oxygen species (ROS) in plant cells [5, 11]. The ROS generates peroxidants of membrane lipids such as MDA [5]. Since MDA content is also regarded as an indicator of salt stress-induced damage to the plasma membrane [13], MDA content in the top leaves of the surviving trees was analyzed after the stress. MDA content in the leaves of the transgenic lines 107-1 and 2-1-1 was about 120 μmol/g FW, while that of the non-transgenic lines No.1 and No.8-7 was >160 μmol/g FW (Fig. 4). The MDA content in the leaves of line M001 that possess salt tolerance was the lowest among the non-transgenic lines (147.7 μmol/g FW). Tukey’s multiple comparison test indicated that MDA content was lower in the leaves of the transgenic lines 107-1 and 2-1-1 than in the leaves of the non-transgenic lines No.1 and No.8-7.

MDA content in the top leaves of 8-month-old trees. The MDA content in trees of each line was analyzed in the salinity stress survivors. Roman letters on the graph bars indicate significant differences at the 0.05 confidence level (Tukey’s multiple comparison test). The white bars represent the non-transgenic lines. The dark gray bars represent the transgenic lines. FW denotes fresh weight. The error bar indicates standard error. The data in each line was derived from three trees for all lines, except line 1-9-1, which only had one tree
Evaluation of Salt Tolerance in Large Juvenile Trees
Since the transgenic line 107-1 showed the highest salt tolerance among the transgenic lines in the salinity tests for young plantlets [30] and small juvenile trees (Fig. 2), the salt tolerance of the 2-year-old large juvenile trees (ca. 3 m in height) of this line and the non-transgenic line No.8-8 was evaluated. It has been reported that the evaluation of salt tolerance in the trees over 1 m in height is useful for salt-shock irrigation, which is single-step irrigation using a high concentration of NaCl [22]. The stress treatment was performed with a single irrigation of 1.3 M NaCl. Fresh-water irrigation was performed from the next day onward. All the leaves from both lines, except from the treetop region of line 107-1, wilted after 1 month. The leaves in the treetop region of line 107-1 remained pale green at this time (Fig. 5a), but those in the region gradually became wilted. Although the entire tree of both lines seemed to be wilted, only the trees of the line 107-1 started to grow lateral buds at the basal part of the tree after continuous irrigation with fresh water to both lines (Fig. 5b). However, lateral buds were not observed on the trees from the non-transgenic line No.8-8.

Salinity test for juvenile trees. Two-year-old trees of the non-transgenic line No.8-8 and the transgenic line 107-1 were evaluated for salinity tolerance (a, b). The stress treatment was performed with a single irrigation with 1.3 M NaCl. Fresh-water irrigation was then performed every day. a Overview of plants the first month after salt-water irrigation. The trees of both lines wilted, but the leaves on the treetop region of line 107-1 were still pale green. Later, the trees of both the lines seem to be completely wilted, but only the trees of line 107-1 began to grow many lateral buds at their basal part. b Overview of plants the third month after salt-water irrigation. The lateral buds at the basal part grew only on the trees of line 107-1. Five trees of line 107-1 and four trees of line No.8-8 were tested
Evaluation of Adult Tree Salt Tolerance
Eucalyptus globulus trees over 2 years of age began to form adult leaves. The 2.5-year-old trees, with adult leaves on just the treetop region, were used for evaluating the salt tolerance of adult trees. Two non-transgenic lines (No.8-11 and No.8-20) and the transgenic line 107-1 were used. Although there were some developmental differences depending on the lines and trees, a part of the tree from each line formed an adult leaf. Just before salt treatment, the glycinebetaine content in each line was determined. The glycinebetaine content in trees of the non-transgenic lines was almost the same for all trees (ca. 0.5 nmol/g FW) except that the levels in the trees from line 107-1 were around 3-times higher (Fig. 6).

Determination of glycinebetaine levels in the leaves of adult trees. The glycinebetaine content in the leaves of trees of the transgenic line 107-1 and the non-transgenic lines No.8-11 and No.8-20 were analyzed. Roman letters on the graph bars indicate significant differences at the 0.05 confidence level (Tukey’s multiple comparison test). The white bars and black bar represent non-transgenic and transgenic lines, respectively. The error bar indicates standard error. Three trees in each line were analyzed
The stress treatment was performed in the same manner as that for large juvenile trees. For line No.8-20, one tree started wilting, and others showed some salt damage on the treetop region (Fig. 7a, b). On the other hand, the salt damage did not appear in all trees of the line 107-1 and line No.8-11 (Fig. 7a, b). It was difficult to evaluate the salt stress tolerance of line 107-1 by single irrigation of salt water, and therefore it was repeated. One month after the second salt-water irrigation, many leaves in the trees of line No.8-20 wilted and fell off. Top leaves on the trees of line No.8-11 started wilting, and the head of the trees bent downward (Fig. 7c, d). On the other hand, the trees of the transgenic line 107-1 showed some salt stress-induced damage in the bottom region, but did not show obvious salt stress-induced damage in the upper region (Fig. 7c, d).


Salinity test for adult trees. The 2.5-year-old trees of the non-transgenic lines No.8-11 and No.8-20 and the transgenic line 107-1 were evaluated for salinity tolerance (a–d). a Overview of plants 1 month after salt-water irrigation. b Treetop region of each tree in (c). Trees of the lines No.8-20 and 107-1 did not show apparent salt stress-induced damage; therefore, salt-water irrigation was repeated. c Overview of plants 1 month after the second salt-water irrigation. d Treetop region of each tree in E. Many leaves in the trees of line No.8-20 wilted and fell off. Top leaves on the trees of line No.8-11 started wilting, and the head of the trees bent downward. On the other hand, the trees of the transgenic line 107-1 showed some salt stress-induced damage in the bottom region, but did not show obvious salt stress-induced damage in the upper region. The red scale bar indicates 10 cm, and the yellow scale bar indicates 1 m
Discussion
Salt Tolerance and Glycinebetaine Content in Small Juvenile Trees
At the growth stage, i.e., 8 months, the transgenic line 107-1 and the non-transgenic lines No.1 and M001 were categorized as “salt-tolerant” by 200 mM NaCl irrigation for 5 weeks (Fig. 2). Although the transgenic lines 1-9-1 and 2-1-1 were shown to have higher tolerance than line No.1 in a previous evaluation of young plantlets [30], they belonged to the salt-sensitive group in our study. It has been reported that the level of salt tolerance alters depending on the growth stage [16, 23, 32]. In spite of transgene expression in all the transgenic lines (Fig. 1), the glycinebetaine content in young leaf before stress treatment in those two transgenic lines (1-9-1 and 2-1-1) was almost the same as that in the non-transgenic lines (Fig. 3). It is possible that the glycinebetaine content did not reach a level high enough to reflect the salt tolerance at this stage. Although those two transgenic lines showed severe salt damage, the glycinebetaine content after the stress treatment was increased to over 1.5 nmol/mg DW in young leaf (Fig. 3). The accumulation of glycinebetaine after stress might not reflect salt tolerance. On the other hand, the trees of the lines 107-1, No.1, and M001 showed only minor salt stress-induced damage (Fig. 2). The glycinebetaine content before stress treatment in the trees of the line 107-1 was around 3-times higher than that in the trees of other lines (Fig. 3). It is expected that the salt tolerance of line 107-1 is enhanced, but it is difficult to make a definitive conclusion from this evaluation alone. Stronger stress treatment may be necessary to evaluate the performance of line 107-1. While the non-transgenic line M001 has been identified as a salt-tolerant line [21], the glycinebetaine levels were not high in the trees of this line. As such, the stress tolerance of the trees of line M001 might not be derived from the accumulation of glycinebetaine.
Salt Tolerance in Large Juvenile and Adult Trees
At the age of 2 and 2.5 years, the trees of the transgenic line 107-1 had higher salt tolerance than the trees of the non-transgenic lines with irrigation of 1.3 M NaCl (Fig. 5). Sensitive lines showed uniform wilting over the entire tree or began wilting from the treetop region (Figs. 5a, 7b, d). In fact, trees of the tolerant line hardly showed salt stress-induced damage in the treetop region (Figs. 5a, 7b, d). E. globulus might possess some protective systems for the treetop region during stress conditions. In small juvenile trees, the glycinebetaine content in young leaves were more than double the levels in mature leaves in all lines (Fig. 3). These young leaves and mature leaves were obtained from treetop and lower portion in small juvenile trees, respectively. Abundance of glycinebetaine in the treetops might reflect that protective system under stress conditions.
Performance Evaluation of Tolerant Lines in a Special Netted-House
At the age of 2 years, since the trees of the line 107-1 seemed to completely wilt at once, a single irrigation with 1.3 M NaCl was assumed to be a slightly stronger stress for the evaluation of large juvenile trees. Fortunately, all the trees of the line 107-1 grew lateral buds at the basal part of tree with the irrigation of fresh water (Fig. 5b). At the growth stage of 2.5 years, as line 107-1 and line No.8-11 did not show any salt damage (Fig. 7b), a single irrigation with 1.3 M NaCl was assumed to be a slightly weaker stress for the evaluation of adult trees. These data indicate that the salt tolerance of E. globulus increases depending on the growth stage. Furthermore, since the special netted-house conditions are easily affected by alterations in the weather as well as seasons, similar to the field conditions, each evaluation was not performed under the same conditions. Therefore, it is difficult to determine the appropriate conditions for evaluating the performance of tolerant lines. The evaluation of tolerant lines in the special netted-house mimics field evaluation. The evaluation in the special netted-house is good for screening and evaluation of practical tolerant lines.
Association Among Damage Parameters, Glycinebetaine Content, and Salt Tolerance
Alterations in the chlorophyll content and MDA generation by stress treatment were evaluated. Both parameters are considered good indicators to evaluate salt damage [3, 13, 19]. In fact, reduction in the chlorophyll content and MDA generation were suppressed in the trees of the transgenic lines compared with the non-transgenic lines (Table 1; Fig. 4). We did not identify a clear association between salt tolerance and the levels of those parameters (Table 1; Figs. 2, 4). Glycinebetaine content showed a similar pattern. Except for the mature leaves before stress, the content was higher in the trees of the transgenic lines than in those of the non-transgenic lines. It is expected that the accumulation of glycinebetaine in the transgenic lines is caused by the expression of the codA gene (Fig. 1). However, there was no association between glycinebetaine content and the levels of salt tolerance. On the other hand, the line 107-1 was the strongest transgenic line (Figs. 2, 5) [30]: reduction in chlorophyll levels and MDA generation following the stress treatment were suppressed in the trees of the line 107-1 (Table 1; Fig. 4). Furthermore, glycinebetaine content was the highest, except in mature leaves. These data indicate that protection of treetop leaves is an important protective mechanism in the trees against salt treatment. In spite of driven codA gene by ubiquitin promoter, glycinebetaine contents were altered in leaf position (young or mature) before and after salt treatment (Fig. 3). CodA synthesizes glycinebetaine from choline. Salt stress stimulates choline biosynthesis from ethanolamine in spinach [25]. Furthermore, nitrate influx and synthesis of both ribulose-1,5-bisphosphate carboxylase/oxygenase small and large subunits in E. globulus were higher in top leaves than in middle and low leaves [26]. The treetop region is biologically important for the maintenance of individual trees and might activate various biochemical reactions. For these reasons, glycinebetaine contents might increase especially in young leaves after salt stress treatment. It is likely that the accumulation of glycinebetaine in the treetop leaves before stress confers salt tolerance with mitigation of salinity stress-induced damage, such as decreasing chlorophyll levels and MDA generation. However, suppression of decreasing chlorophyll levels or MDA generation is not directly associated with conferring salt tolerance. Influence of saline circumstance except saline wind is started from roots. Passive absorption of some ions and adversity of water resorption occurred in roots. But, our evaluation focused on the damage of aerial parts in trees. From the study in Arabidopsis, overexpressor of codA gene had elongated roots under 100 mM NaCl condition and seemed to get enough water to grow by increased glycinebetaine [6]. Actually, 107-1 showed high salinity stress tolerance from the aerial parts evaluation. Root systems of all trees were established without saline condition since those were grown using tap water in special netted-house until salinity test. Further physiological evaluation in root might be necessary for our transgenic lines under salt stress treatment.
Practical Utilization
Although trees of all three transgenic lines showed salt tolerance under controlled conditions [17], the transgenic line 107-1 was the most tolerant line among the transgenic lines at the young plantlet stage, as identified from our evaluation of salt tolerance in a special netted-house [30]. In addition, the transgenic line 107-1 showed salt tolerance at the large juvenile stage (Fig. 2) as well as later stages (Figs. 5, 7). Since evaluation in a special netted-house mimics field evaluation, line 107-1 is a candidate practical tolerant line. To evaluate the practical utility of line 107-1, it is necessary to evaluate its salt tolerance performance in the field. Fortunately, 107-1 was selected as salt-tolerant line via evaluation in special netted-house. Although transformation system in E. globulus was already established, there are some difficulties in rooting from shoot buds and in acclimatization of small plantlets. It took over 3 years from starting generation of transformants to the selection of some candidate lines in vitro [17].
On the other hand, the non-transgenic line M001 also showed high-salt tolerance. Since the strength of salt tolerance in the lines 107-1 and M001 were not compared in this study, a field trial should be done to this end. Glycinebetaine content was not high in the line M001 (Fig. 3). It is expected that the salt tolerance of the line M001 is not derived from the accumulation of glycinebetaine. The tolerance of line 107-1 indicated that the codA gene could confer resistance to salt stress in E. globulus. However, it is difficult to introduce a transgene into E. globulus without involving a seedling. It is suggested that introducing the codA gene in high-salt-tolerating lines such as line M001 may result in a super-salt-tolerant transgenic tree after applying gene improvement techniques. It might be necessary that the transformation system improve at the points of plant materials (without seedling), the rooting efficiently and the acclimatization of small plantlets. The results of this study provide valuable information that may ultimately lead to the practical use of transgenic plants in addressing issues related to food supply, natural resources, and environment.
References
Adams, M., Richter, A., Hill, A. K., & Colmer, T. D. (2005). Salt tolerance in Eucalyptus spp.: identity and response of putative osmolytes. Plant, Cell and Environment, 28, 772–787.
Barry, K. M., Newnham, G. J., & Stone, C. (2009). Estimation of chlorophyll content in Eucalyptus globulus foliage with the leaf reflectance model PROSPECT. Agricultural and Forest Meteorology, 149, 1209–1213.
Corney, H. J., Sasse, J. M., & Ades, P. K. (2003). Assessment of salt tolerance in eucalypts using chlorophyll fluorescence attributes. New Forests, 26, 233–246.
Eldridge, K., Davidson, J., Harwood, C., & Wyk, G. V. (1993). Eucalypt domestication and breeding. London: Oxford University Press.
Ghanem, M. E., Albacete, A., Martinez-Andujar, C., Acosta, M., Romero-Aranda, R., Dodd, I. C., et al. (2008). Hormonal changes during salinity-induced leaf senescence in tomato (Solanum lycopersicum L.). Journal of Experimental Botany, 59, 3039–3050.
Hayashi, H., Alia, Mustardy, L., Deshnium, P., Ida, M., & Murata, N. (1997). Transformation of Arabidopsis thaliana with the codA gene for choline oxidase; accumulation of glycinebetaine and enhanced tolerance to salt and cold stress. Plant Journal, 12, 133–142.
Hibino, T. (2009). “Post-genomics” research in Eucalyptus in the near future. Plant Biotechnology, 26, 109–113.
Hu, L., Lu, H., Liu, Q. L., Chen, X. M., & Jiang, X. N. (2005). Overexpression of mtlD gene in transgenic Populus tomentosa improves salt tolerance through accumulation of mannitol. Tree Physiology, 23, 1273–1281.
Ikuta, S., Imamura, S., Misaki, H., & Horiuchi, Y. (1977). Purification and characterization of choline oxidase from Arthrobacter globiformis. Journal of Biochemistry, 82, 1741–1749.
James, S. A., Smith, W. K., & Vogelmann, T. C. (1999). Ontogenetic differences in mesophyll structure and chlorophyll distribution in Eucalyptus globulus ssp. globulus (Myrtaceae). American Journal of Botany, 86, 198–207.
Katsuhara, M., Otsuka, T., & Ezaki, B. (2005). Salt stress-induced lipid peroxidation is reduced by glutathione S-transferase, but this reduction of lipid peroxides is not enough for a recovery of root growth in Arabidopsis. Plant Science, 169, 369–373.
Khan, M. S., Yu, X., Kikuchi, A., Asahina, M., & Watanabe, K. N. (2009). Genetic engineering of glycine betaine biosynthesis to enhance abiotic stress tolerance in plants. Plant Biotechnology, 26, 125–134.
Kikuchi, A., Kawaoka, A., Shimazaki, T., Yu, X., Ebinuma, H., & Watanabe, K. N. (2006). Trait stability and environmental biosafety assessments on three transgenic Eucalyptus lines (Eucalyptus camldulensis Dehnh. codA 12–5B, codA 12–5C, codA 20-C) conferring salt tolerance. Breeding Research, 8, 17–26. (in Japanese).
Kikuchi, A., Watanabe, K. N., Tanaka, Y., & Kamada, H. (2008). Recent progress on environmental biosafety assessment of genetically modified trees and floricultural plants in Japan. Plant Biotechnology, 25, 9–15.
Loreto, F., & Delfine, S. (2000). Emission of isoprene from salt-stressed Eucalyptus globulus leaves. Plant Physiology, 123, 1605–1610.
Maeda, Y., Ukai, H., & Takenaga, H. (2002). Salt tolerance of five temperate grasses at germination and different growth stages. Journal of the Japanese Society of Revegetation Technology, 27, 499–506.
Matsunaga, E., Nanto, K., Oishi, M., Ebinuma, H., Morishita, Y., Sakurai, N., et al. (2012). Agrobacterium-mediated transformation of Eucalyptus globulus using explants with shoot apex with introduction of bacterial choline oxidase gene to enhance salt tolerance. Plant Cell Report, 31, 225–235.
McComb, J. (2007). Selection of salt-waterlogging tolerant genotypes. In Salt tolerance hybrid eucalypts. (pp. 12–15). Kingston: Rural Industries Research and Development Corporation.
Meneguzzo, S., Sgherri, C. L. M., Navari-Izzo, F., & Izzo, R. (1998). Stromal and thylakoid-bound ascorbate peroxidases in NaCl-treated wheat. Physiologia Plantarum, 104, 735–740.
Michelozzii, M., Johnson, J. D., & Warrag, E. I. (1995). Response of ethylene and chlorophyll in two eucalyptus clones during drought. New Forests, 9, 197–204.
Murakami, A. (2006). Development of salt tolerant Eucalyptus globulus selection. Japan Technical Association of the Pulp and Paper Industry Journal, 60, 69–74. (in Japanese).
Niknam, S. R., & McComb, J. (2000). Salt tolerance screening of selected Australian woody species—a review. Forest Ecology and Management, 139, 1–19.
Nukaya, A., Masui, M., & Ishida, A. (1984). Salt tolerance of muskmelons as affected by diluted sea water applied at different growth stages in nutrient solution culture. Journal of the Japanese Society for Horticultural Science, 53, 168–175.
Sakamoto, A., & Murata, N. (2001). The use of bacterial choline oxidase, a glycinebetaine-synthesizing enzyme, to create stress-resistant transgenic plants. Plant Physiology, 125, 180–188.
Summers, Peter S., & Weretilnyk, Elizabeth A. (1993). Choline synthesis in spinach in relation to salt stress. Plant Physiology, 103(1269–1), 276.
Suzuki, Y., Kihara-Doi, T., Kawazu, T., Miyake, C., & Makino, A. (2010). Differences in Rubisco content and its synthesis in leaves at different positions in Eucalyptus globulus seedlings. Plant Cell and Environment., 33, 1314–1323.
Tabei, Y. (1999). Environmental risk assessment of transgenic melon in Japan. Current Topics in Microbiology and Immunology, 16, 65–68.
Tang, W. (2002). Regeneration of transgenic loblolly pine expressing genes for salt tolerance. Journal of Forest Research, 13, 1–6.
Tang, W., Peng, X. X., & Newton, R. J. (2005). Enhanced tolerance to salt stress in transgenic loblolly pine simultaneously expressing two genes encoding mannitol-1-phosphate dehydrogenase and glucitol-6-phosphate dehydrogenase. Plant Physiology and Biochemistry, 43, 139–146.
Yu, X., Kikuchi, A., Matsunaga, E., Morishita, Y., Nanto, K., Sakurai, N., et al. (2009). Establishment of the evaluation system of salt tolerance on transgenic woody plants in the special netted-house. Plant Biotechnology, 26, 135–141.
Yu, X., Kikuchi, A., Shimazaki, T., Yamada, A., Ozeki, Y., Matsunaga, E., Ebinuma, H., Watanabe K. N. (2012). Assessment of the salt tolerance and environmental biosafety of Eucalyptus camaldulensis harboring a mangrin transgene. Journal of Plant Research (in press).
Zeng, L., Shannon, M. C., & Grieve, C. M. (2002). Evaluation of salt tolerance in rice genotypes by multiple agronomic parameters. Euphytica, 127, 235–245.
Acknowledgments
We would like to thank Dr. Norio Murata for providing the codA gene. This research was supported in part by the Japanese Society for the Promotion of Science (JSPS) (Grant-in-Aid for Scientific Research (A) No. 21248001) and a research grant from the New Energy and Industrial Technology Development Organization (NEDO) of Japan. The authors are very grateful for their support.
Author information
Authors and Affiliations
Corresponding author
Additional information
Xiang Yu, Akira Kikuchi has contributed equally as first author.
Rights and permissions
About this article
Cite this article
Yu, X., Kikuchi, A., Matsunaga, E. et al. The Choline Oxidase Gene codA Confers Salt Tolerance to Transgenic Eucalyptus globulus in a Semi-Confined Condition. Mol Biotechnol 54, 320–330 (2013). https://doi.org/10.1007/s12033-012-9575-y
Published:
Issue Date:
DOI: https://doi.org/10.1007/s12033-012-9575-y