
Abstract
The present study aimed to investigate the biological functions of miR-96 in the processes of proliferation and clonogenicity in the prostate cancer cells. miR-96 was identified to be markedly up-regulated in prostate cancer cell and cancer tissues compared with normal prostate cell and normal prostate tissues by microarray method and RT-PCR analysis. Down-regulation of miR-96 expression reduced the proliferation and colony formation ability of PC3 prostate cancer cells, while over-expression of miR-96 induced proliferation and colony formation ability of LNCaP prostate cancer cells. Forkhead box protein O1 (FOXO1) is key tumor suppressors and has been shown to play key roles in the regulation of diverse cellular processes, including cell proliferation, differentiation, cell cycle progression and apoptosis. The expression level of FOXO1 was strikingly up-regulated in PC3 cells after transfected with miR-96 inhibitor, and FOXO1 expression was down-regulated in LNCaP cells after transfected with miR-96 mimics. miR-96 may play a vital role in promoting cell proliferation in human prostate cancer cells. Inhibition of miR-96 caused expression increase of tumor suppressor gene FOXO1, thus manipulating miR-96 expression may be a promising approach in treatment of prostate cancer.
Similar content being viewed by others
Avoid common mistakes on your manuscript.
Introduction
Prostate cancer is one of the most common diagnosed malignant diseases in men and the leading causes of cancer deaths in males [1]. Although in the past decade, many studies were conducted to investigate the precise mechanism that mediated development and progression of prostate cancers, molecular mechanisms remain poorly understood [2]. Therefore, understanding of the mechanisms involved in the development and progression of cancers is critical to identify novel targets for effective therapeutic strategies.
MicroRNAs (miRNAs) are a class of endogenous small non-coding, single-stranded RNA with 20–22 nucleotides that can interfere with protein expression either by inducing translational repression or mRNA degradation [3]. miRNAs have been implicated in the control of many fundamental cellular activities, such as cell proliferation, differentiation, apoptosis and they played important roles in carcinogenesis [4, 5]. Accumulating evidence indicates that miRNAs are frequently dysregulated in several of human cancers and act as oncogenes or tumor suppressors [6, 7]. miR-96 is a member of the miR-183-96-182 polycistronic miRNA cluster that is located at chromosome 7q32.2 [8]. Recently, miR-96 was found to be up-regulated in prostate cancer [9]. However, functional analysis of miR-96 has not been carried out in prostate cancer cells in vitro, and the detailed role of miR-96 in prostate cancer remains poorly understood.
Herein, the expression of miR-96 was markedly up-regulated in prostate cancer cells and cancer tissues compared with normal prostate cells and normal prostate tissues, and miR-96 acts a significant regulatory role in prostate cancer cell proliferation by regulation of the expression of FOXO1. Our results showed that it might be a novel target for further studies of the therapeutics of prostate cancer.
Materials and methods
Patient samples
Human prostate cancer specimens and their corresponding non-tumorous prostate samples were obtained from 13 patients who had received radical prostatectomy at Department of Urology, Shanghai First People’s Hospital, School of Medicine, Shanghai Jiaotong University, China. Samples were snap frozen in liquid nitrogen and stored at −80 °C for future isolation of RNA. All experimental procedures were approved by the Institutional Review Board of Shanghai First People’s Hospital. The detail information about characteristics of patients was summarized in Table 1.
miRNA microarray
Six RNA samples were firstly extracted from three prostate cancer samples and three from their corresponding non-tumorous prostate samples and then labeled with FlashTag™ Biotin RNA Labeling Kit. The miRNA hybridization and washing steps were carried out following Affymetrix GeneChip® miRNA 3.0 Array’s instructions. Affymetrix GeneChip miRNA 3.0 Array is one high-density array comprised of 179,217 probes that represent 19,913 mature microRNA contained in miRBase V17. For human, this single array covered all annotated human miRNA-based miRBase, version 17. Limma (Linear Models for Microarray Data) is used to identify differentially expressed miRNA between prostate cancer samples and non-tumorous prostate samples. After Benjamini–Hochberg correction, only the miRNAs under 10 % FDR and with more than twofold change difference were considered as differentially expressed miRNAs. miRNAs with genomic distance <10 K were considered as from the same miRNA cluster.
Cell culture and transient transfections
Human PCa cell lines LNCaP and PC3 were obtained from Shanghai Cell Bank, Chinese Academy of Sciences, and cultivated in RPMI 1640 medium supplemented with 50 U/mL penicillin, 50 mg/mL streptomycin and 10 % fetal bovine serum (Gibco, Invitrogen) in a humidified atmosphere at 37 °C and 5 % CO2. The normal prostate cell lines RWPE-1 were purchased from American Type Culture Collection (Manassas, VA, USA), cultured in keratinocyte serum-free medium (K-SFM, Invitrogen, CA, USA) and supplemented with bovine pituitary extract and human recombinant epidermal growth factor. Human prostatic epithelial cell line BPH-1 was preserved in our laboratory and maintained in RPMI 1640 medium supplemented with 10 % FBS. Cells were transfected with miR-96 mimics, negative control and miR-96 inhibitor purchased from GenePharma Company (Shanghai, China) performing with Lipofectamine 2000 reagent (Invitrogen) according to the manufacturer’s instructions.
RNA isolation and real-time PCR
Total RNA from prostate cancer tissue samples and cell lines was extracted using Trizol reagent (Invitrogen). Reverse transcription and quantitative real-time PCR (qRT-PCR) was carried out using the PrimeScript Reverse Transcription System and SYBR Premix Ex Taq™ II kit (Takara, Dalian, China) according to the manufacturer’s instructions. The relative expression of mRNA compared with GAPDH was calculated using the 2−ΔCt method. The primers used are as the following: FOXO1 forward: 5′-TGTCCCTACACAGCAAGTTCA-3′; reverse: 5′-CACCCTCTGGATTGAGCATC-3′ and GAPDH forward: 5′-TCGACAGTCAGCCGCATCTTCTTT-3′, reverse: 5′-ACCAAATCCGTTGACTCCGACCTT-3′. Quantification of mature miR-96 was assayed using RT-PCR kit (Ribobio Co., Ltd, Guangdong, China) analysis according to the manufacturer’s protocol. U6 small nuclear RNA was used as a control to determine relative miRNA expression.
CCK-8 and EdU analysis
Cell proliferation was assessed by the Cell Counting Kit-8 (CCK-8) assay (Dojindo, Japan). The transfected cells of PC3, LNCaP were plated with 100 μL culture medium in 96-well plates at a density of 3,000 cells per well. After 48 and 72 h, 10 μL CCK-8 reagent (5 mg/mL) was added to each well and incubated at 37 °C for 2 h. Viable cells were evaluated by absorbance measurements at 450 nm. Each assay was performed in six replicates on three independent experiments. The impact of miR-96 expression on PC3 and LNCaP cell proliferation was also assessed by the EdU cell proliferation assay kit, according to the manufacturer’s instructions. The assays were performed as recommended by the manufacturer of EdU detection kits (Apollo488, RiboBio, China).
Plate clone formation assay
Twenty-four hours after transfection, cells were seeded into each well of 6-well plates at the density of 3,000/well and cultured in 1,640 containing 10 % FBS at 37 °C for 14 days. The colonies were washed with PBS, fixed with 10 % formaldehyde for 5 min and stained with 1.0 % crystal violet for 30 s. The colonies that had 50 cells or more per colony were counted.
Western blotting
Cells were washed twice with cold PBS and homogenized in ice-cold RIPA buffer (Beyotime Institute of Biotechnology, China) containing phosphatase and protease inhibitors. Total protein (50 μg) was separated by denaturing 10 % SDS-polyacrylamide gel electrophoresis and transferred electrophoretically onto PVDF membranes. Membranes were incubated with the primary antibodies for rabbit-FOXO1, rabbit-GAPDH (cell signaling technology) at 4 °C overnight and subsequently with horseradish peroxidase-conjugated secondary antibody. Signals were performed with ECL chemiluminescence kit (Boster, Wuhan, China) and exposured to X-ray films.
Statistical analysis
Statistical analyses were performed using the Statistical Package of the Social Sciences software (SPSS) version 17.0. Data were presented as mean ± standard deviation (SD) from at least three independent experiments. The Student’s t test was used to assess statistically significant differences. A P < 0.05 was considered statistically significant difference.
Results
miR-96 is up-regulated in prostate cancer tissues and cancer cell lines
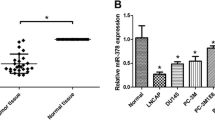
To investigate the expression profiles of miRNA in prostate cancer and non-tumorous prostate samples, global miRNA expression levels were measured using Affymetrix miRNA microarray assay. We identified 26 miRNAs with expression levels that differed significantly between cancer tissues and non-tumorous prostate samples and derived from miRNA clusters (See Materials and methods). Among them, we observed significantly increased expression of miR-96 in prostate cancer tissues (Fig. 1a). To confirm the up-regulation of miR-96 in prostate cancer tissues, we measured miR-96 expression levels in 10 cases of prostate cancer tissue samples by real-time PCR. Indeed, we observed that the expression of miR-96 is significantly increased in these prostate cancer tissues compared that in normal prostate tissues except a pair of sample (Fig. 1b). Besides, miR-96 expression was also elevated in prostate cancer PC3 cell lines compared with normal prostate epithelial cells (RWPE-1 and BPH-1). We observed that miR-96 is not significantly increased in LNCaP cells compared with the normal prostate cell line (Fig. 1c).

Up-regulation of miR-96 in human prostate cancer. a Comparison of differentially expressed miRNA genes in human prostate cancer and non-tumorous prostate tissues. Hierarchical clusters of significantly altered miRNAs (as determined by Limma) across different samples. Red denotes low expression levels, whereas green depicts high-expression levels. Each miRNA listed is significantly differentially expressed between the prostate cancer and non-tumorous prostate tissues and derived from miRNA clusters. b miR-96 is overexpressed in prostate cancers compared to normal prostate tissues. c Real-time PCR analysis of relative miR-96 expression levels in normal human prostate and prostate cancer cell lines. *P < 0.05 compared with RWPE-1 cell
Effect of miR-96 over-expression on cell proliferation and colony formation in LNCaP cell lines
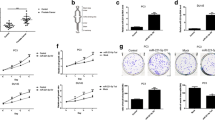
In order to confirm the biological function of miR-96, we transfected miR-96 mimics into LNCaP prostate cancer cell line and examined its effect on cell proliferation and colony formation. By using CCK-8 and plate clone formation assay, we observed significantly increased cell growth rate (Fig. 2a) and colony formation (Fig. 2b, c) in miRNA-96 mimic-transfected cells compared with miR-NC-transfected cells. These data suggest that up-regulation of miR-96 could augment the tumorigenicity of LNCaP prostate cancer cell in vitro. Furthermore, the effect of miR-96 on proliferation was also measured using the EdU incorporation assay, which is similar to BrdU incorporation assay. The percentage of EdU positively stained cells was determined to assess differences. More EdU-positive cells were found in miR-96 mimic-transfected LNCaP cells(42.7 ± 1.2 %) than in control cells (32.6 ± 1.2 %) (Fig. 2d, e). Above all, data indicate over-expression of miR-96 promoted proliferation of LNCaP cells.

Effect of miR-96 overexpression on prostate cancer cells. Cells of LNCaP were treated with miR-96 inhibitor or negative control. a CCK-8 assay was performed at 48 and 72 h after LNCaP cells were transfected with miR-96 mimics and negative control. b miR-96 increased prostate cancer cells LNCaP colonies formation. Representative images are shown, and colonies were counted after staining with 0.1 % crystal violet. c Quantification of crystal violet stained cell colonies. d EdU labeling showing proliferation of LNCaP cell transfected with miR-96 mimics and miRNA negative control. e Quantification of ErdU-incorporating cells after transfection with miR-96 mimics and negative control. The percentage of positive cells was derived from triplicate samples. *P < 0.05 compared with control group
Effect of miR-96 knock down on cell proliferation and colony formation in PC3 cell lines
As described above, we assessed the effect of miR-96 on the cell proliferation, colony formation and cell cycle of prostate cancer LNCaP cells. Next, we further examined the role of inhibiting miR-96 in prostate cancer cells. CCK-8 assay investigated the impact of inhibiting miR-96 on cell proliferation. The result indicated that down-regulation of miR-96 could reduce the PC3 cells growth rates compared with negative control (Fig. 3a). Ectopically expression of miR-96 inhibitor in PC3 cells also significantly reduced the cell colony formation (Fig. 3b, c). Fewer EdU-positive cells were found in miR-96 inhibitor-transfected PC3 cells(34.3 ± 3.1 %) than in control cells (40.9 ± 1.2 %) (Fig. 3d, e). These data indicate that down-regulation of miR-96 inhibited PC3 cell proliferation in vitro.

Inhibition of miR-96 suppressed proliferation of prostate cancer cells. a CCk-8 assays revealed that transfection with miR-96 inhibitor reduced PC3 cells growth. b Representative micrographs of crystal violet stained cell colonies. c Quantification of miR-96 down-expression significantly inhibits the colony formation ability of PC3 cells. Each bar represents the mean of three independent experiments. d Representative micrographs of EdU-incorporating cells after transfection with miR-96 inhibitor or NC. e Quantification of EdU-incorporating cells after transfection with miR-96 inhibitor or NC. Values represent the mean of three independent experiments. *P < 0.05 compared with control group
miR-96 regulate the expression of FOXO1
We transfected prostate cell lines LNCaP and PC3 with miR-96 mimic and miR-96 inhibitor, respectively, and examined the effects of miR-96 on expression of FOXO1 (Fig. 4). Western blot demonstrated that the protein levels of FOXO1 were decreased in miR-96 mimic-transfected LNCaP cells compared with cells transfected with the negative control. The effect of inhibiting miR-96 on expression of FOXO1 of prostate cancer cells was further examined. FOXO1 inhibition efficiency was also confirmed by Western blot analysis. Suppression of miR-96 could increase the protein expression level of FOXO1 in PC3 cells. These results indicated that FOXO1 might be a potential miR-96 target gene.

miR-96 regulated the expression of FOXO1 in PC3 and LNCaP cells. Western blotting analysis of FOXO1 expression in PC3 cell after transfection with miR-96 inhibitor and negative control, FOXO1 expression was up-regulated after PC3 cells transfection with miR-96 inhibitor, and Western blotting analysis showed that the FOXO1 expression was lower in LNCaP transfection with miR-96 mimic than in negative control
Discussion
miRNAs with high influence on target protein complexes play a role in prostate cancer progression and are promising diagnostic or prognostic biomarkers. In human prostate cancers, several miRNAs have been identified as oncogenes due to their overexpression in malignant tissue [10, 11], and their functional analyses have been performed [12]. Overexpression of miR-183-96-182 cluster has already been reported in prostate tissue and reduced zinc uptake, demonstrating this miRNA cluster as a regulator of zinc homeostasis [13]. The current study confirmed that miR-96 was increased in prostate cancer PC3 cells and prostate cancer tissues as compared with that in normal prostate epithelial cell RWPE-1 and normal prostate tissues. Recent evidence also showed that elevated levels of miR-96 are correlated with high Gleason score and cancer recurrence after radical prostatectomy [14]. Larne [15] discovered miR-96-5p, miR-183-5p, miR-145-5p and miR221-5p discriminatory miRNAs in prostate cancer were combined with a quota, denoted the miRNA index quote (miQ); ((miR-96-5p × miR-183-5p)/(miR-145-5p × miR221-5p)). miQ has potential to be used as a clinical tool for prostate cancer diagnosis and as a prognostic marker of disease progression. In contrast to above data, the latest report has not observed a significant correlation between the expression of miR-96, -145 and -221 and clinicopathologic parameters in prostate cancer [16]. Thus, implication of miR-96 in prostate cancer as a diagnostic tool in clinical practice, it is critical to do more research to understand its mechanisms and further characterize its function.
In this study, we found that ectopic expression of miR-96 could promote the proliferation and clonogenicity in LNCaP cells, while prostate cancer PC3 cells were transfected with inhibitor of miR-96, these effects was reduced. Collectively, these data suggest that up-regulation of miR-96 may act as a vital role in promoting carcinogenesis and progression of prostate cancer. To explore the mechanism by which miR-96 suppressed the proliferation and clonogenicity of prostate cancer cells, we used gain-of-function and loss-of-function approaches to studying miR-96 in prostate cancer cells, and the protein level of FOXO1 was significantly regulated. The FOXO subfamily of forkhead transcription factors characterized by the presence of a winged-helix DNA binding domain called a forkhead box [17]. Forkhead box O (FOXO) transcription factors FOXO1, FOXO3a, FOXO4 and FOXO6 are emerging as an important family of proteins that modulate the expression of genes involved in apoptosis, the cell cycle, DNA damage repair, oxidative stress, cell differentiation, glucose metabolism and other cellular functions [18], and FOXO proteins are regulated by multiple mechanisms. The levels of FOXO1 proteins are depressed in prostate cancer [19] and described as the tumor suppressor gene in prostate cancer, and expression of FOXO1 knockdown promoted cell proliferation or survival [20], but the underlying mechanisms are not well understood.
Overexpression of miR-96 was also found to in endometrial cancer that functions in concert to repress FOXO1 expression. FOXO1 expression was efficiently restored in the Ishikawa cell line upon simultaneous inhibition of miR-96 [21]. Our department has identified that reduced expression of FOXO1 in renal cell carcinoma and the expression of FOXO1 mRNA and miR-27a had inverse relation [22]. FOXO1 inhibits Runx2 transcriptional activity and prostate cancer cell migration and invasion [23]. Ectopic expression of FOXO1 can up-regulate the cell cycle inhibitors p21Cip1 and p27Kip1, and down-regulate the cell cycle regulator cyclin D1, consequently leading to G1/S cell cycle arrest [24]. Recent evidence suggests that the FOXO1 expression is regulated by miRNAs. For example, aberrant miR-182, miR-27a, miR-96, miR-153 and miR-183 regulate the expression of FOXO1 through directly targeting the FOXO1 3′-UTR [25–27]. Guo and their colleagues revealed that miR-96 expression was sufficient to significantly reduce FOXO1 expression in transitional cell carcinoma and miR-96 inhibitor-mediated T24 cell apoptosis [28]. We demonstrated that transfection of a miR-96 inhibitor in PC3 cells led to up-regulated FOXO1. Overexpression of miR-96 in LNCaP cells decreased FOXO1 protein expression. These data indicate that miR-96 may increase the proliferation of prostate cancer cell lines through regulating FOXO1.
In summary, our study identified that there is increased expression of miR-96 in prostate cancer and that miR-96 showed an important abilities in promoting carcinogenesis and progression of prostate cancer in vitro by affecting proliferation, clonogenicity. These findings suggest that miR-96 may act as oncogenic function in human prostate cancer, so inhibiting miR-96 may offer a novel gene therapy targets for treating human prostate cancer.
References
Jemal A, Siegel R, Ward E, Murray T, Xu J, Smigal C, et al. Cancer statistics, 2006. CA Cancer J Clin. 2006;56(2):106–30.
De Marzo AM, Platz EA, Sutcliffe S, Xu J, Gronberg H, Drake CG, et al. Inflammation in prostate carcinogenesis. Nat Rev Cancer. 2007;7(4):256–69. doi:10.1038/nrc2090.
Iorio MV, Croce CM. MicroRNAs in cancer: small molecules with a huge impact. J Clin Oncol. 2009;27(34):5848–56. doi:10.1200/jco.2009.24.0317.
Lu Z, Liu M, Stribinskis V, Klinge CM, Ramos KS, Colburn NH, et al. MicroRNA-21 promotes cell transformation by targeting the programmed cell death 4 gene. Oncogene. 2008;27(31):4373–9. doi:10.1038/onc.2008.72.
Bartel DP. MicroRNAs: target recognition and regulatory functions. Cell. 2009;136(2):215–33. doi:10.1016/j.cell.2009.01.002.
Shi XB, Tepper CG, deVere White RW. Cancerous miRNAs and their regulation. Cell Cycle. 2008;7(11):1529–38.
Esquela-Kerscher A, Slack FJ. Oncomirs—microRNAs with a role in cancer. Nat Rev Cancer. 2006;6(4):259–69. doi:10.1038/nrc1840.
Wang Y, Huang JW, Calses P, Kemp CJ, Taniguchi T. MiR-96 downregulates REV1 and RAD51 to promote cellular sensitivity to cisplatin and PARP inhibition. Cancer Res. 2012;72(16):4037–46. doi:10.1158/0008-5472.CAN-12-0103.
Jung M, Schaefer A, Steiner I, Kempkensteffen C, Stephan C, Erbersdobler A, et al. Robust microRNA stability in degraded RNA preparations from human tissue and cell samples. Clin Chem. 2010;56(6):998–1006. doi:10.1373/clinchem.2009.141580.
Pang Y, Young CYF, Yuan H. MicroRNAs and prostate cancer. Acta Biochim Biophys Sin. 2010;42(6):363–9. doi:10.1093/abbs/gmq038.
Fletcher CE, Dart DA, Sita-Lumsden A, Cheng H, Rennie PS, Bevan CL. Androgen-regulated processing of the oncomir MiR-27a, which targets Prohibitin in prostate cancer. Hum Mol Genet. 2012;. doi:10.1093/hmg/dds139.
Folini M, Gandellini P, Longoni N, Profumo V, Callari M, Pennati M, et al. miR-21: an oncomir on strike in prostate cancer. Mol Cancer. 2010;9:12. doi:10.1186/1476-4598-9-12.
Mihelich BL, Khramtsova EA, Arva N, Vaishnav A, Johnson DN, Giangreco AA, et al. miR-183-96-182 Cluster is overexpressed in prostate tissue and regulates zinc homeostasis in prostate cells. J Biol Chem. 2011;286(52):44503–11. doi:10.1074/jbc.M111.262915.
Schaefer A, Jung M, Mollenkopf HJ, Wagner I, Stephan C, Jentzmik F, et al. Diagnostic and prognostic implications of microRNA profiling in prostate carcinoma. Int J Cancer. 2010;126(5):1166–76. doi:10.1002/ijc.24827.
Larne O, Martens-Uzunova E, Hagman Z, Edsjo A, Lippolis G, den Berg MS, et al. miQ—a novel microRNA based diagnostic and prognostic tool for prostate cancer. Int J Cancer. 2013;132(12):2867–75. doi:10.1002/ijc.27973.
Kang SG, Ha YR, Kim SJ, Kang SH, Park HS, Lee JG, et al. Do microRNA 96, 145 and 221 expressions really aid in the prognosis of prostate carcinoma? Asian J Androl. 2012;14(5):752–7. doi:10.1038/aja.2012.68.
Greer EL, Brunet A. FOXO transcription factors at the interface between longevity and tumor suppression. Oncogene. 2005;24(50):7410–25. doi:10.1038/sj.onc.1209086.
Huang H, Tindall DJ. Dynamic FoxO transcription factors. J Cell Sci. 2007;120(Pt 15):2479–87. doi:10.1242/jcs.001222.
Li R, Erdamar S, Dai H, Wheeler TM, Frolov A, Scardino PT, et al. Forkhead protein FKHR and its phosphorylated form p-FKHR in human prostate cancer. Hum Pathol. 2007;38(10):1501–7. doi:10.1016/j.humpath.2007.02.016.
Dong XY, Chen C, Sun X, Guo P, Vessella RL, Wang RX, et al. FOXO1A is a candidate for the 13q14 tumor suppressor gene inhibiting androgen receptor signaling in prostate cancer. Cancer Res. 2006;66(14):6998–7006. doi:10.1158/0008-5472.CAN-06-0411.
Myatt SS, Wang J, Monteiro LJ, Christian M, Ho KK, Fusi L, et al. Definition of microRNAs that repress expression of the tumor suppressor gene FOXO1 in endometrial cancer. Cancer Res. 2010;70(1):367–77. doi:10.1158/0008-5472.can-09-1891.
Zhou L, Yin B, Liu Y, Hong Y, Zhang C, Fan J. Mechanism and function of decreased FOXO1 in renal cell carcinoma. J Surg Oncol. 2012;105(8):841–7. doi:10.1002/jso.23013.
Zhang H, Pan Y, Zheng L, Choe C, Lindgren B, Jensen ED, et al. FOXO1 inhibits Runx2 transcriptional activity and prostate cancer cell migration and invasion. Cancer Res. 2011;71(9):3257–67. doi:10.1158/0008-5472.can-10-2603.
Wu Z, Sun H, Zeng W, He J, Mao X. Upregulation of MircoRNA-370 induces proliferation in human prostate cancer cells by downregulating the transcription factor FOXO1. PLoS ONE. 2012;7(9):e45825. doi:10.1371/journal.pone.0045825.
Guttilla IK, White BA. Coordinate regulation of FOXO1 by miR-27a, miR-96, and miR-182 in breast cancer cells. J Biol Chem. 2009;284(35):23204–16. doi:10.1074/jbc.M109.031427.
Wu Z, He B, He J, Mao X. Upregulation of miR-153 promotes cell proliferation via downregulation of the PTEN tumor suppressor gene in human prostate cancer. Prostate. 2012;. doi:10.1002/pros.22600.
Myatt SS, Wang J, Monteiro LJ, Christian M, Ho KK, Fusi L, et al. Definition of microRNAs that repress expression of the tumor suppressor gene FOXO1 in endometrial cancer. Cancer Res. 2010;70(1):367–77. doi:10.1158/0008-5472.CAN-09-1891.
Guo Y, Liu H, Zhang H, Shang C, Song Y. miR-96 regulates FOXO1-mediated cell apoptosis in bladder cancer. Oncol Lett. 2012;4(3):561–5. doi:10.3892/ol.2012.775.
Acknowledgements
This study was supported by the grants from the National Natural Science Foundation of China (No. 81072114).
Conflict of interest
The authors have no conflict of interest to disclose.
Author information
Authors and Affiliations
Corresponding authors
Additional information
Junjie Yu and Yinxia Wu have contributed equally to this work.
Rights and permissions
About this article
Cite this article
Yu, JJ., Wu, YX., Zhao, FJ. et al. miR-96 promotes cell proliferation and clonogenicity by down-regulating of FOXO1 in prostate cancer cells. Med Oncol 31, 910 (2014). https://doi.org/10.1007/s12032-014-0910-y
Received:
Accepted:
Published:
DOI: https://doi.org/10.1007/s12032-014-0910-y