
Abstract
Excessive accumulation and deposition of amyloid-beta (Aβ) has been considered as a pivotal event in the pathogenesis of Alzheimer’s disease (AD). Neuronal apoptosis is one of the characteristics of AD, which is a possible mechanism underlying Aβ-induced neuronal neurotoxicity. Neuroglobin (Ngb) is a newly discovered vertebrate heme protein that exhibits neuroprotective functions against cell death associated with hypoxic and amyloid insult. However, until now, the exact mechanism of neuroglobin’s protective action has not been determined. To investigate the potential neuroprotective roles and mechanisms of Ngb, transgenic AD mice (APPswe/PSEN1dE9) and SH-SY5Y cells transfected with pAPPswe were enrolled into the study. In vivo, overexpression of Ngb via intracerebroventricular injection with pNgb attenuated memory, cognitive impairment, and plaque generations. In pAPPswe transfected SH-SY5Y cells, Ngb not only decreased the generation of Aβ42, but also attenuated mitochondrial dysfunction and apoptosis through suppressing the activation of caspase-3, caspase-9 by Akt activating phosphorylation, which were restrained by phosphatidylinositol 3-kinase inhibitor (LY294002). Our data indicate the anti-apoptotic property of Ngb may play a neuroprotective role against AD.
Similar content being viewed by others
Avoid common mistakes on your manuscript.
Introduction
AD is the most common chronic neurodegenerative disorder which affects elderly people, characterized by a progressive decline in cognitive function and learning in clinical setting. Usually, it is considered that senile plaques (SPs), formation of neurofibrillary tangles (NFTs) and neuron loss are three major pathological hallmarks of AD. And, neuronal apoptosis is the main manifestation of neuronal loss (Yuan and Yanker 2000). Excessive accumulation of Aβ peptide has been proposed as not only a common pathway in the pathogenesis of AD induced by various reasons, but also a key factor that induces neuronal apoptosis and leading to cognitive dysfunction (Golde et al. 2006), although the precise mechanism by which Aβ induces neuronal apoptosis is still unknown.
Caspases family is a type of specific apoptotic signal transduction molecular. Its activation is considered to be one of the most critical elements in the mechanisms of apoptosis. There are two main initiation pathways of apoptosis: the death receptor (extrinsic) pathway and the mitochondrial (intrinsic) pathway. The latter can be triggered by various cytotoxic stresses, cytochrome c in the presence of dATP initiates the oligomerization of Apaf-1 (apoptotic protease activating factor-1) into a large protein scaffold, the apoptosome, which recruits and processes pro-caspase 9 into an active initiator caspase 9. The initiator caspase 9 then activates the executioner caspases, such as caspase 3, which brings about the apoptotic destruction of the cell. It has been proved that caspases not only mediate the process of apoptosis of cortical and hippocampal neurons of AD (Yu et al. 2010), but also relate with Aβ (Hoi et al. 2010), Aβ precursor protein (APP) (Francois et al. 1999), Presenilin 1 (Hedskog et al. 2011), and NFTs (De Calignon et al. 2010), all of which are tightly related with the pathogenesis of AD. Therefore, how to inhibit the activity of caspases related with neuronal apoptosis may be a pivotal strategy for prevention and treatment of AD (Galimberti and Scarpini 2011).
Neuroglobin (Ngb) is a newly discovered oxygen-binding globin protein family by Burmester et al. (2000). Ngb is widely distributed in the human body, including cerebral cortex, hippocampus, thalamus, hypothalamus, and cerebellum and other organs with endocrine function. Previously, most studies were focused on the role played by Ngb in neuronal protection in response to cerebral hypoxia and ischemia injury in vivo, such as stroke (Jin et al. 2010). Recently, a few studies suggested that Ngb also plays a neuroprotection against AD (Fordel et al. 2006). However, the molecular mechanisms of the protection of Ngb against AD remains largely not elucidated (Wei et al. 2011; Richard et al. 2008).
It has been suggested that Ngb is an inducer of PI3K/Akt signaling pathway (Antao et al. 2010), which is an important method of treatment with degenerative disease in future (Zhao et al. 2011). Moreover, it is known that caspase 9 is a downstream target gene of PI3K/Akt pathway (Redlak and Miller 2011), and once its activated form (phosphorylated caspase 9) was inhibited, the apoptosis process would be avoided, so, we hypothesized that the protective role against Aβ-induced apoptosis provided by Ngb could arise from its intervention of caspase activity by activating PI3K/Akt signaling pathway.
Materials and Methods
Animals
Double-transgenic mice (APPswe/PS1dE9) used in this study were purchased from Nanjing Institute of Animal Model. They all expressed a chimeric human amyloid precursor protein containing the Swedish mutation (HuAPP695swe) and a mutant human presenilin 1 (PS1-dE9) gene. All mice were housed in plastic cages (three to four mice/cage) with free access to food and water and were maintained under controlled conditions of humidity (50 ± 10 %), light (12/12-h light/dark cycle), and temperature (23 °C ± 1).
All procedures were in accordance with the Guide for the Care and Use of Laboratory Animals adopted by the National Institutes of Health (USA) and approved by the Ethical Committee of Chongqing Medical University, China. Twelve male and 12 female APP/PS1 double transgenic mice, aged 13 months, weighing 30–35 g, were randomly split into three groups: intracerebroventricular injection with normal saline, saline + pCDNA3.1 (empty vector), pCDNA3.1 + pNgb, respectively.
Intracerebroventricular (ICV) Injection
All the mice were anesthetized by intraperitoneal injection of about 1 mL 10 % chloral hydrate (3.5 mL/kg) and held by the head on the stereotaxic instrument (Taimeng, Chengdu, China). A polyethylene cannula was implanted into the lateral ventricle (coordinates of 0.2 mm posterior, 1.3 mm lateral, and 2.5 mm ventral to the bregma) and secured to the skull with dental cement. About 5 μL of saline, saline + pCDNA3.1 (1 mg/mL), pCDNA3.1 (1 mg/mL) + pNgb were injected into lateral ventricle, respectively, once a week, continuously for 8 weeks (Zhu and Tang 2010).
Reference Memory Protocol by Morris Water Maze
In this task, the mice received five training sessions (one session/day) and a probe trial on the 6th day. Each session consisted of four trials with a 15-min interval. A trial began when the mice was placed in the water at one of the four starting positions, chosen at random, facing the wall. The order of the starting position varied in each trial and any given sequence was not repeated on acquisition phase days. The mice were given 60 s to locate the platform. If the animal did not succeed, it was gently guided to the platform and left on it for 10 s. Mice were dried and returned to their home cages after each trial. The latency to find the platform was measured in each trial and the mean latency for each training day was calculated. The probe consisted of a single trial, as described before, with the platform removed. Here, the latency to reach the original platform position, the number of crossings over that place and the time spent on the target, as well as in the opposite quadrants, were measured. Sessions were recorded with a video acquisition system. Videotapes were used by a trained observer using the software. Videos were subsequently placed in randomized order in a separate ANY-maze protocol to be scored by a trained observer blind to the experimental condition using a keyboard-based behavioral tracking system.
Cell Culture and Plasmid Transfection
To achieve high expression, a cDNA encoded with wild type Ngb was synthesized using a modification of recursive PCR strategy. First, vector GV144 was enzymed by XhoI/Bam HI (Gene Chemical Tech Com., Shanghai, China). Then, the primer of Ngb was designed, former: 5′-TCC GCT CGA GCT ATG GAG CGC CCG GAG CCC GA′-3; reverse: 5′-ATC GGG ATC CTT ACT CGC CAT CCC AGC CTC GA′-3. The PCR product was connected into the linear vector and identified with restriction enzyme site-Bam HI/XhoI. Finally, the positive RT-PCR products were cloned and sequenced. After verification, pNgb transformation was performed with a plasmid extraction kit (TianGen Biochemical Co. LTD, Beijing, China). Finally, the extracted pNgb was dissolved with EB fluid, and the concentration of pNgb was measured by a Fluorescence Spectrophotometer ND-1000. The human neuroblastoma cell line SH-SY5Y was cultured in Dulbercco modified Eagle medium (DMEM) supplemented with 10 % fetal bovine serum, 2 mM glutamine, 100 μg/mL Streptomycin, and 100 units/mL Penicillin. Cells were maintained at 37 °C in an incubator containing 5 % CO2. One day before transfection, cells were grown in 75-cm2 flasks to approximately 70 % confluence: cells of each flask was transfected with 8.0 μg pAPPswe (gifted by Prof. Weihong Song, The University of British Columbia, Vancouver, BC, Canada) or 4.0 μg pNgb (Genechem Com, Shanghai, China) using 30 μL of Lipofectamine2000 Reagent (Invitrogen, USA) with Opti-MEM® Medium to show high enough transfection efficiency according to the manufacturer’s instructions (Zhang et al. 2011). For LY294002 treatment, the concentration was 5.0 μM in the culture medium.
Cell Viability Assays
Cell viability was measured using the MTT reduction assay as described previously (Wang et al. 2014). After the experiments, MTT was added to each well and incubated at 37 °C for different time. Then, the insoluble formazan was dissolved with dimethyl sulfoxide; the reduction product was measured by reading absorbance at 540 nm in a Universal Microplate Reader (Gene Company, USA).
Measurement of Aβ by ELISA Assay
Conditioned media were collected from cells and mice brain tissues were harvested. Protein inhibitors and AEBSF were added to the media to prevent degradation of Aβ protein. The concentration of Aβ40/42 was detected by Aβ 1–42 Colorimetric ELISA kit according to the manufacturer’s instructions. For brain samples, the values of Aβ42 concentration were normalized with total brain protein concentrations and expressed in nanograms of Aβ42 per milligram total protein (nanogram per milligram of protein).
Mitochondrial Membrane Potential
The mitochondrial membrane potential was measured using JC-1 fluorescent dye kit (Beyotime Institute of Biotechnology, Beijing, China). According to the manufacturer’s instruction, the transfected SH-SY5Y cells were incubated with JC-1 dye for 30 min. After incubation, cells were rinsed with PBS. The emission signals at 590 and 529 nm elicited by excitation at 514 nm were measured with a spectrofluorometer. The ratio of the signal at 590 nm over that at 514 nm (red/green ratio) was then calculated.
Cell Apoptosis Assessment by Flow Cytometry
For cells apoptotic analysis, the SH-SY5Y cells were seeded in a 6-well culture plate at a concentration of 1 × 104 cells/well and cultured in an incubator with DMEM medium +10 % FBS. After different treatment, the cells were harvested, washed twice with ice-cold PBS and resuspended in the dark with AnnexinV-FITC and PI (KeyGEN Biotech, Nanjing, China) buffer for 15 min at room temperature. Cells were then analyzed with a flow cytometry. Cells were considered to be apoptotic when they were either Annexin V+/PI− (early apoptotic) or Annexin V+/PI+ (late apoptotic).
Immunohistochemistry and Image Analysis
The staining protocol employed a modified streptavidin-HRP immunohistochemistry procedure (CoWin Century Biotechnology, Inc, China). Briefly, after initial deparaffinization, endogenous peroxidase activitity was blocked with 0.3%H2O2. Deparaffinized sections were microwaved in 10 mmol/l citrate buffer (pH 6.0). The sections were pretreated with normal goat serum for 30 min at 37 °C and were incubated overnight with the rabbit anti-mouse Aβ1–42 monoclonal antibody (Bioss, Beijing, China, 1:500). The slides were treated with peroxidase-conjugated streptavidin and visualized by the diaminobenzidine (DAB) Kit (CoWin Century Biotechnology, Inc, China), and sections were counterstained with hematoxylin. For quantitative analysis, immunostaining in the cortex and hippocampus was defined as the percentage of Aβ1–42 positive area in relation to the total area, the image analysis was done as described previously (Lim et al. 2001).
Western Blot Assay
Tissues were lysed with PRO-PREPTM liquid (Beyon, China) in the presence of a protease inhibitor cocktail (Roche, Welwyn Garden, Hertfordshire, UK). Lysates were then spun at 13,000×g for 15 min at 4 °C. The total protein concentration was determined by the Bradford method employing a Universal Microplate Reader (Gene Company, Hong Kong) at 595 nm. Equal amount (50 μg) of total protein was separated by 10 % SDS-PAGE gels and transferred to polyvinylidene fluoride (PVDF) membranes (Millipore, Billerica, MS, USA). The membranes were blocked in 5 % solution (TBS-T containing non-fat milk) 1 h and were incubated with primary antibodies directed against neuroglobin (Bioworld Technology, USA) pAkt (Bioworld Technology), Akt (Bioworld Technology), caspase-3 (Bioworld Technology), caspase-9 (Bioworld Technology), and β-actin (Bioss Technology Limited Company, Peking, China), diluted in 0.1 % (v/v) non-fat dry milk powder and incubated overnight at 4 °C. After washing, bounded antibodies were detected by incubation for 1–2 h at room temperature with secondary peroxidase-conjugated anti-mouse or anti-rabbit antibodies (Zhong Shan Golden Bridge Bio-tech Company, China). Membranes were then developed using a commercial enhanced chemiluminescence system (Bio-Rad, USA) and quantitated using Quantitive One Image Analysis (Bio-Rad, USA).
Statistical Analysis
All the data were normally distributed, and presented as means ± SD; the homogeneity test for variance was performed using SPSS 11.5 software. The significance of differences between the groups was judged using one-way ANOVA followed by Bonferroni post hoc test, the difference of escape latency was analyzed with two-way repeated measures ANOVA followed by Bonferroni post hoc test. Results were considered statistically significant at P < 0.05.
Results
Ngb Improves the Memory and Cognition Function of the APP/PS1 Transgenic Mice
We tested spatial learning and memory of mice with Morris water maze 8 weeks after ICV. All mice from each group showed a day-to-day decrease trend in escape latency, after 4 days tests, APP/PS1 mice ICV with pNgb displayed significantly shorter escape latencies(38.5 ± 2.0 s) than the Saline group (44.5 ± 2.3 s) and the empty vector group (45.7 ± 1.8 s) (P < 0.01) (Fig. 1a). Furthermore, APP/PS1 mice treated with pNgb crossed the platform location more times after the platform was removed (Fig. 1b), spent significantly more time within target quadrant (Fig. 1d) and traveled longer distance in the target quadrant (Fig. 1d), compared to the mice ICV with Saline or empty vector, the pNgb group not only had crossed the quadrant more number of times, but also had a longer staying time and covered a longer distance in the quadrant, and there was a statistical significance (P < 0.05). We still tested mice on the visible platform task and found no differences among the three groups in their latency to find the visible platform (data not shown). The improved learning and memory function of the APP/PS1 mice ICV with pNgb was indicated by reduced escape latency, increased target quadrant occupation, and more frequency of platform crossings.

The Morris water maze test of mice. a Escape latencies of mice in the water maze during the five consecutive days test. Two-way repeated measures ANOVA followed by Bonferroni post hoc test, p < 0.0001, F = 354.1. APP/PS1 + pNgb versus APP/PS1 + saline, p < 0.001, t = 9.681. APP/PS1 + pNgb versus APP/PS1 + empty vector, p < 0.001, t = 4.572. b The numbers of crossing the quadrant of platform. c The distance traveled by the mice to the hidden platform in the Morris water maze d time spent in the target quadrant. All the data are reported as means ± SD (n = 8, *P < 0.05, **P < 0.01, ***P < 0.001. ns: non-significant)
Overexpression of Ngb Significantly Attenuates Aβ42 Deposition and Production in the APP/PS1 Transgenic Mice
Extracellular accumulation of amyloid beta is one of the main neuropathological features of AD. To determine the effect of Ngb on Aβ deposition, we analyzed Aβ burden in 13 months old APP/PS1 transgenetic mice treated with pNgb for 8 weeks. The transgenetic mice treated with Saline or with empty vector show large, round and compact Aβ plaques mainly distributed in the cortex (Fig. 2a) and hippocampus (Fig. 2b). The mice treated with pNgb showed decreased Aβ plaques of cortex (Fig.2a) and hippocampus (Fig. 2b) in size and number. Quantification of Aβ burden which was expressed as percentage of Aβ positive area per total area, showed a significantly decreased Aβ load in cortex (1.08 ± 0.12 %; Fig. 2c) and hippocampus (0.32 ± 0.09 %; Fig. 2d) of transgenetic animals treated with pNgb in comparison to cortex (1.40 ± 0.16 %) and hippocampus (0.54 ± 0.10 %) of transgenetic animals treated with Saline or cortex (1.41 ± 0.21 %) and hippocampus (0.53 ± 0.13 %) of transgenetic animals treated with Empty vector (Fig. 2c, d; P < 0.05).

Overexpression of Ngb significantly attenuates Aβ42 production and load in the APP/PS1 transgenic mice. Immunohistochemistry of Aβ plaques in cortex (a) of and hippocampus (b) of the mice in the three groups were studied with Aβ1–42 monoclonal antibody (*100). The amyloid burden in cortex (c) of and hippocampus (d) was expressed as the percentage of Aβ42-positive area in relation to the total area. The concentration of Aβ42 of mouse brain was detected by ELISA Assay (e), Aβ42 concentration was normalized to brain protein concentration which was expressed as ng/mg protein. All the data are shown as means ± SD. (n = 8, *P < 0.05, **P < 0.01. ns: non-significant. Scale bar, 200 μm)
To determine whether Aβ42 production was effected by pNgb, we measured cerebral levels of and Aβ42 in whole-brain homogenates using specific sandwich ELISAs. The concentration of Aβ42 in mice intracerebroventricularly treated with saline or empty vector was 105.9 ± 13.5 pg/mg protein, 107.4 ± 14.7 pg/mg protein, respectively (Fig. 2e; p > 0.05). While, the concentration of Aβ42 was significantly decreased to 79.5 ± 22.5 pg/mg protein in comparison to the mice treated with saline or empty vector (Fig. 2e; P < 0.01), which indicated that Ngb inhibited Aβ42 production.
Overexpression of Ngb Promotes pAPPswe-Transfected SH-SY5Y Cell Survival and Inhibits Apoptisis
To detect the effect of overexpression of Ngb on the cell viability, cellular responses were assessed by MTT assay. In SH-SY5Y cells only transfected with pAPPswe as control, the absorbance at 540 nm was 0.687 ± 0.098, and absorbance was 0.704 ± 0.053 in pAPPswe cells transfected with empty vector, these two groups showed no significant difference (p > 0.05; Fig. 3a). In pAPPswe cells transfected with pNgb for 48 h showed significant difference with absorbance 0.883 ± 0.074, compared to control or empty vector group (n = 6, #P < 0.01, vs. control, *P < 0.01, vs. Vector), which indicated that overexpression of Ngb can improve the cell survival of pAPPswe-transfected SH-SY5Y cell.

Effect of pNgb transfection on cell survival and apoptisis. a Cell viability was assessed by MTT assay (n = 6, ***P < 0.001). The absorbance at 540 nm in cells transfected only with pAPPswe as control showed no difference from the pAPPswe cells transfected with empty vector (p > 0.05). In pAPPswe, cells transfected with pNgb for 48 h showed significant difference in comparison to control or empty vector group(n = 6, ***P < 0.001, ns: non-significant). Data were expressed as optical density (OD) values. b The apoptotic behavior of the APPswe-transfected SH-SY5Y cells were analyzed by flow cytometry. The percentage of apoptotic cells were characterized as those that stained with Annexin-V and membrane integrity was simultaneously assessed by Propidium Iodide (PI) exclusion. Flow cytometry profile represents Annexin-V-FITC staining in x axis and PI in y axis. The ratios of the early apoptotic cells were calculated to be 0.78 ± 0.27 % in control group and 0.93 ± 0.33 % in empty vector group, which were higher than that in pNgb group (0.24 ± 0.11 %). Similarly, the ratios of the later apoptotic cells in control (4.43 ± 1.50 %) and empty vector group (4.04 ± 0.59 %) were higher than that of the pNgb group (2.53 ± 1.03 %). Experiments were repeated three times
Flow cytometric analysis was performed to study the influence of overexpression of Ngb on the apoptotic behavior of the APPswe-transfected SH-SY5Y cells (Fig. 3b). The percentage of apoptotic cells were characterized as those that stained with Annexin-V and membrane integrity was simultaneously assessed by propidium iodide (PI) exclusion. The ratios of the early apoptotic cells were calculated to be 0.78 ± 0.27 and 0.93 ± 0.33 % for the control and empty vector group, respectively, which were higher than that of pNgb group (0.24 ± 0.11 %). Similarly, the ratios of the later apoptotic cells in control (4.43 ± 1.50 %) and empty group (4.04 ± 0.59 %) were higher than that of the pNgb group (2.53 ± 1.03 %).
Overexpression of Ngb Improves Mitochondrial Membrane Potential in pAPPswe-Transfected SH-SY5Y Cell
The decrease of mitochondrial membrane potential is an early sign of cell apoptosis, and it can be easily detected through the transformation of JC-1 into green fluorescence from red fluorescence, which can be as an indicator of early sign of cell apoptosis. In the cells transfected only with pAPPswe, the JC-1 ratio of red to green fluorescence was 50.7 ± 3.5 %, while the ratio was 52.4 ± 4.2 % in the vector, and there was no significant difference between the two groups. In pAPPswe cells after transfection with pNgb, the ratio was 76.5 ± 5.9 %, and there was a significant increase compared to the pAPPswe group or empty vector (n = 4, *P < 0.01 vs. Vector, #P < 0.01 vs. control, Fig. 4). The data suggest that overexpression of Ngb improves mitochondrial membrane potential and attenuates mitochondrial dysfunction. The ratio of the signal at 590 nm over that at 514 nm (red/green ratio) was then calculated.

The effect of pNgb on the mitochondrial membrane potential assessed by JC-1 ratio. The mitochondrial membrane potential of pAPPswe transfected SH-SY5Y cells treated with pNgb. The mitochondrial membrane potential was decreased in pAPPswe transfected SH-SY5Y cells before transfection with empty vector or pNgb. Then, empty vector or pNgb (4.0 μg/106) was transfected into the cells for 48 h. After treated with pNgb, the JC-1 ratio of pAPPswe transfected SH-SY5Y cells was significantly increased (*P < 0.001) however in empty vector treated group had no difference in comparison to that of pAPPswe transfected SH-SY5Y cells (p > 0.05, ns: non-significant). SH-SY5Y cells before transfection with pAPPswe was used as control for JC-1 ratio (100 %)
Overexpression of Ngb Significantly Attenuates Aβ42 Production in pAPPswe Transfected SH-SY5Y Cells
In this study, Aβ42 deposition and production in the APP/PS1 transgenic miceAs were attenuated by overexpressed Ngb, and we also confirmed the experiment in cell lines. In media of pAPPswe-transfected SH-SY5Y cells (as vehicle group), the concentration of Aβ42 was 34.1 ± 3.5 pg/mL. And after transfection with vector, the concentration of Aβ42 in pAPPswe-transfected SH-SY5Y cells treated with empty vector was 33.5 ± 4.7 pg/mL, no significant difference between these two groups (p > 0.05, ns: not significant, Fig. 5). While, the concentration of Aβ42 in was significantly decreased by overexpression of Ngb to 21.3 ± 2.5 pg/mL compared with that in vehicle group or empty vector group (n = 3, ***P < 0.001).

The concentration of Aβ42 of pAPPswe transfected SH-SY5Y cells treated with pNgb. The pAPPswe transfected SH-SY5Y cells promoted the production of Aβ42, then, we transfected pNgb (4.0 μg/106) into pAPPswe transfected SH-SY5Y cells for 48 h and assessed the concentration of Aβ42 by ELISA. In pNgb-treated group, the concentration was significantly decreased. There was no difference between pAPPswe transfected SH-SY5Y cells (vehicle group) and empty vector group. The values represent mean ± SD (n = 3, **P < 0.01, ns: not significant)
Overexpression of Ngb Inhibits Caspase-3/9 Protein Expression Through Activating PI3K/Akt Pathway in Vitro and in Vivo
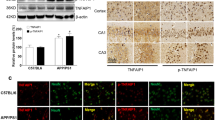
The overexpression of neuroglobin in vivo and vitro were verified by western blot. The SH-SY5Y cells were transfected with 1.0, 2.0, 4.0, and 8.0 μg pNgb, which showed the high expression of neuroglobin was found in the transfection with 4.0 μg pNgb (Fig. 6a). Also, when the APP/PS1 transgenetic mice intracerebroventricularly injected with pNgb showed stably high expression of neuroglobin (Fig. 6b), caspase activity was considered as an indicator of apoptosis. In the previous study, we foud that Ngb could improve mitochondrial potential of the cells, that is to say that Ngb could inhibit apoptosis induced by the generation and deposition of much more Aβ. So, we further investigated the effect of Ngb on the two important members of caspase family, caspase-3 or caspase-9, and explored its potential pathway. As shown in Fig. 6, Caspase-3 and caspase-9 expressions at protein level were inhibited in pAPPswe-transfected SH-SY5Y cells treated with pNgb, compared to the control and the vector group (**P < 0.01, Fig. 6c, d). Furthermore, we studied caspase-3 and caspase-9 expression in vivo. Expression of caspase-3 and caspase-9 at protein level was significantly decreased in the ICV with pNgb group, compared to saline group and the empty vector group (**P < 0.01, Fig. 6e, f). Phosphorylation of Akt was significantly induced in pAPPswe-transfected SH-SY5Y cells treated with pNgb (**P < 0.01, vs control or vector, Fig. 6c, d) without any induction of total Akt in all the groups (p > 0.05). To analyze the PI3K/Akt signaling pathway which could participate in the neuroprotective mechanism of Ngb, the cells were treated with PI3K inhibitor LY294002 and the expression of Akt (total Akt and pAkt) and expression of caspase-3and caspase-9 were analyzed. We found that PI3K inhibitor LY294002 could counteract the effect of Ngb on the expression of caspase-3 or caspase-9 and pAkt (##P < 0.01, vs pNgb, Fig. 6c, d).

Expression of caspase-3/9 was inhibited and phosphorylation of Akt was in enhanced by overexpression of Ngb. aThe pAPPswe-transfected SH-SY5Y cells were transfected with 1.0, 2.0, 4.0, and 8.0 μg pNgb, which showed the high expression of neuroglobin was found in the transfection with 4.0 μg pNgb. b the APP/PS1 transgenetic mice intracerebroventricularly injected with pNgb showed stably high expression of neuroglobin. c Representative pictures western blot of caspase-3/9, pAkt, Akt and β-actin of pAPPswe-transfected SH-SY5Y cells in control, empty vector, pNgb and pNgb + LY294002 group, respectively. d The relative optical density (OD) of caspase-3/9, pAkt, Akt at protein level was compared to β-actin, respectively. The data represent the mean ± S.D of a typical series of three experiments (*P < 0.01, vs control or vector; #P < 0.01, vs pNgb). Expressions of caspase-3 and caspase-9 in the brains of APPswe/PS1dE9 transgenic mice. e Representative pictures western blot of caspase-3, caspase-9, and β-actin in the brains of the mice in saline, empty vector and pNgb group, respectively. f The relative optical density (OD) of caspase-3, caspase-9 at protein level was compared to β-actin, respectively. The data represent the Mean ± SD (**P < 0.01, saline group vs pNgb group; ##P < 0.01, empty vector group vs pNgb group)
Discussion
It is well-known that deposition of Aβ within the cerebral cortices not only leads the formation of SPs, but also induces neuronal apoptosis, which are key pathogenesis of AD (Zara et al. 2011). Aβ is cleaved by β- and γ-secretase from APP in an abnormal way. The neurotoxicity of Aβ is manifested in many forms. Especially, neuron apoptosis induced by Aβ may be tightly linked with cognitive dysfunction (Song et al. 2015). Therefore, depressing the generation of Aβ may enable delayed occurrence of apoptosis or prevention of apoptosis altogether in AD.
Ngb is a newly discovered oxygen carrying globulin in human and murine brain. It is expressed in various regions of the brain and at particularly high levels in neurons, some endocrine cell types and in retinal rod cells. Previous studies have already documented that overexpression of Ngb can play its antiapoptotic neuroprotective roles against the brain from damage by AD (Brittain et al. 2010; Khan et al. 2007). However, its mechanisms remain mysterious and not fully elucidated. The aim of our work was to evaluate the effects of Ngb on the generation of Aβ and Aβ-induced damage, and to explore the potential mechanisms and the downstream signaling events in vitro and in vivo.
Human neuroblastoma cell line SH-SY5Y is one kind of tumor cells with low differentiation, and it has similarity with normal nerve cells at cell morphology, physiology and biochemical functions. In previous study, we have used SH-SY5Y cells as an experimental model to study the pathogenesis of AD and its preventive measures (Zhang et al. 2011). Therefore, we still inferred that SH-SY5Y cells transfected with pAPPswe to mimic the characteristic of AD as a valid model in the current in vitro experiments.
Previous studies have already demonstrated decreased Ngb levels with age in several human and rat brain regions, suggesting a possible relation between Ngb deficiency and age-related neurodegeneration (Sun et al. 2005; Szymanski et al. 2010). Furthermore, Sun et al. (Sun et al. 2013), measured Ngb levels by western blot and immunostained hippocampal sections for Ngb, cell-type protein markers, and Aβ in brain tissue obtained at autopsy from patients with AD. The results suggest that at least hippocampal Ngb is increased in early and moderately advanced AD, returning toward control levels in severe AD, suggesting the expression of Ngb was in a dynamic change with the changes of severity of AD. Therefore, 13-months-old double transgenic AD mice were enrolled, and the expression of Ngb was very low and they should be suitable for the study in vivo.
Firstly, in the study in vitro, Ngb overexpressed cells, derived from a neuroblastoma SH-SY5Y cell line transfected with pAPPswe, not only have significantly increased resistance to cell death and promoted survival rate, but also have remarkedly suppressed Aβ generation and counteracted the mitochondrial dysfunction and apoptosis induced by excessive Aβ. While, in vivo study results showed that after ICV with pNgb, overexpression of Ngb not only has improved the spatial memory and cognition functions of the mice, but also has significantly decreased the production of Aβ and Aβ-induced apoptosis cells. Then, the potential mechanisms of the effects of overexpression of Ngb were explored.
The exact mechanism by which Ngb achieves its neuroprotective activity in CNS disease is still controversial. A number of potential roles have been suggested for Ngb since its discovery in 2000. At first, researches took it for granted that Ngb could provide a reserve oxygen strore because it was a newly carrying oxygen globin and it was similar to other heme protein such as myoglin and hemoglobin. However, further studies have showed that Ngb undergoes rapid autoxidation and has a relatively low binding rate with oxygen (Schmidt et al. 2003; Brunori and Vallone 2006). Then, another viewpoint was proposed: Ngb was a nitric oxide (NO) scavenger (Zara et al. 2013; Singh et al. 2013). Recently, neuroglobin has been shown to inhibit Rac1-mediated actin assembly and aggregation of membrane microdomains which are involved in regulation of death-signal transduction pathways (Hajra and Liu 2004). In fact, there existed many reports about an anti-apoptotic effect of Ngb both in hypoxic ischemic diseases (Hajra and Liu 2004) and in neurodengenerative diseases (Ord et al. 2013). Our results in vitro and in vivo were also in accordance with the viewpoint.
The activation of caspases is considered to be one of the most critical elements in the mechanisms of apoptosis. Many studies have shown that caspases activation is tightly related with the pathogenesis of AD. Li et al. (2011) observed that the activity of caspase-2, 3, 6 was increased during the apoptosis in astrocytes and cerebellar granule cells induced by excess Aβ. And the inhibitor of caspase-2, caspase-3, caspase-6 have obviously played protective role against apoptosis, respectively. Francois et al. (1999) have proved that APP has three splice cites of caspase-3: (P4)DNVD 197(P1)/S, (P4) DYAD219 (P1)/G, (P4)VEVD720 (P1)/A. One peptide fragment C31 was from APP cleaved by caspases and involved in apoptosis induced by Aβ, and the apoptosis promoted caspases activation and APP cleavage. Raina et al. (2001) also found that the activation of caspase-3 was consistent with the cleavage of βAPP, elevated Aβ and the formation of SPs in the cerebral cortex and hippocampal neurons of AD. That is to say that the activation of caspases destroys the normal metabolism of intracellular APP, and causes overproduced Aβ and its deposition. Aβ-mediated neurotoxicity further promotes the cleavage of APP by caspases, which were forming a vicious cycle. Therefore, we hypothesized that inhibiting the activity of caspases may decrease the generation of Aβ and inhibit the cell apoptosis in AD.
In our study, the expression of caspase-3 and caspase-9 was high in pAPPswe-transfected SH-SY5Y cells; and overexpression of Ngb significantly inhibited the expression of caspase-3 and caspase-9, compared to the control and the vector groups. So, we concluded that the mechanism of Ngb playing a role of decreasing the generation of Aβ and Aβ-induced apoptosis may be through inhibiting the activity of caspase-3 and caspase-9.
Caspase-9 can activate caspase-3, which can specifically cleave DNA, and make poly ADP ribose polymerase (PARP) and DNA-dependent protein kinase (DNA-PK) involved in the course of DNA repair after damage to be inactivated. Then, the chromatin was condensed and the ribozyme was activated, and cell apoptosis occurred. And in turn, caspase-3 can activate caspase-9, and they were forming a positive feedback. The previous study showed that the application of PI3K inhibitor with high concentration would block Akt phosphorylation, reduce the phosphorylation of caspase-9, inhibit the process of apoptosis, and promote neural cell survival (Li et al. 2008). In the present study, our results that Ngb-induced caspase-3, caspase-9 and phosphorylation of Akt were prevented by PI3K/Akt inhibitor LY294002 indicate that the upstream signaling pathway responsible for the increased caspase-3 and caspase-9 by Ngb is also PI3K/Akt. Regulatory effect of activated PI3K/Akt on its downstream target gene is an important way to promote cell survival; especially it is playing a key role in cell survival mediated by growth factors, which provide a potential target for treating AD.
In conclusion, Ngb not only decreases Aβ generation, but also attenuates Aβ-induced mitochondrial dysfunction, apoptosis, and cell death. The potential mechanism may be through inhibiting caspase activity via activating PI3K/Akt pathway. Further exploration of Ngb anti-apoptotic properties may provide opportunities for novel pharmacological interventions aimed at preventing or palliating the consequences of AD.
References
Antao ST, Duang TT, Aran R, Witting PK (2010) Neuroglobin overexpression in cultured human neuronal cells protects against hydrogen peroxide insult via activating phosphoinositide-3 kinase and opening the mitochondrial K (ATP) channel. Antioxid Redox Signal 13(6):769–781
Brittain T, Skommer J, Raychaudhuri S, Birch N (2010) An antiapoptotic neuroprotective role for neuroglobin. Int J Mol Sci 11(6):2306–2321
Brunori M, Vallone B (2006) A globin for the brain. FASEB J 20:2192–2197
Burmester T, Weich B, Reinhardt S, Hankeln T (2000) A vertebrate globin expressed in the brain. Nature 407(6803):520–523
De Calignon A, Fox LM, Pitstick R et al (2010) Caspase activation precedes and leads to tangles. Nature 464(7292):1201–1204
Fordel E, Thus L, Martinet W et al (2006) Neuroglobin and cytoglobin overexpression protects human SH-SY5Y neuroblastoma cells against oxidative stress-induced cell death. Neurosci Lett 410(2):146–151
Francois G, Daigen X, George S et al (1999) Involvement of caspases inproteolytic cleavage of Alzheimer’ amyloid-beta precursor protein and amyloidogenin Abeta peptide formation. Cell 97(3):395–406
Galimberti D, Scarpini E (2011) Alzheimer’s disease: from pathogenesis to disease-modifying approaches. CNS Neurol Disord Drug Targets 10(2):163–174
Golde TE, Dickson D, Hutton M (2006) Filling the gaps in the Abeta cascade hypothesis of Alzheimer’s disease. Curr Alzheimers Res 3(5):421–430
Hajra KM, Liu JR (2004) Apoptosome dysfunction in human cancer. Apoptosis 9:691–704
Hedskog L, Petersen CA, Al S et al (2011) Gamma-secretase complexes containing caspase-cleaved presenlin-1 increase intracellular Aβ42/Aβ40 ratio. Cell Mol Med 15(10):2150–2163
Hoi CP, Ho YP, Baum L, Chow AH (2010) Neuroprotective effect of honokiol and magnolol compunds from Magnolia officinalis on beta-amyloid-induced toxicity in PC12 cells. Phytother Res 24(10):1538–1542
Jin K, Mao Y, Mao X, Xie L, Greenberg DA (2010) Neuroglobin expression in ischemic stroke. Stroke 41(3):557–559
Khan AA, Mao XO, Banwait S, Jin K, Greenberg DA (2007) Neuroglobin attenuates beta-amyloid neurotoxicity in vitro and transgenic Alzheimer phenotype in vivo. Proc Natl Acad Sci U S A 104(48):10114–10119
Li RC, Pouranfar F, Lee SK, Morris MW, Wang Y, Gozal D (2008) Neuroglobin protects PC12 cells against beta-amyloid-induced cell injury. Neurobiol Aging 29(12):1815–1822
Li C, Zhao R, Gao K et al (2011) Astrocytes: implication for neuroinflammatory pathogenesis of Alzheimer’s disease. Curr Alzheimers Res 80(1):67–80
Lim GP, Chu T, Yang F, Beech W, Frautschy SA, Cole GM (2001) The curry spice curcumin reduces oxidative damage and amyloid pathology in an Alzheimer transgenic mouse. J Neurosci 21(21):8370–8377
Ord EN, Shirley R, McClure JD et al (2013) Combined antiapoptotic and antioxidant approach to acute neuroprotection for stroke in hypertensive rats. J Cereb Blood Flow Metab 33(8):1215–1224
Raina AK, Hochman A, Zhu X et al (2001) Abortive apoptosis in Alzheimer’s disease. Acta Neuropathol 101(4):305–310
Redlak MJ, Miller TA (2011) Targeting PI3K/Akt/HSP90 signaling sensitizes gastric cancer cells to deoxycholate-induced apoptosis. Dig Dis Sci 56(2):323–329
Richard CL, Pouranfar F, Lee SK, Matthew WM, Yang W, Gozal D (2008) Neuroglobin protects PC12 cell against β-amyloid induced cell injury. Neurobiol Aging 29(12):1815–1822
Schmidt M, Giessl A, Laufs T, Hankeln T, Wolfrum U, Burmester T (2003) How does the eye breathe? Evidence for neuroglobin-mediated oxygen supply in the mammalian retina. J Biol Chem 278:1932–1935
Singh S, Zhuo M, Gorgun FM, Englander EW (2013) Overexpressed neuroglobin raises threshold for nitric oxide-induced impairment of mitochondrial respiratory activities and stress signaling in primary cortical neurons. Nitric Oxide 32:21–28
Song Z, Insel PS, Buckley S et al (2015) Brain amyloid-beta burden is associated with disruption of intrinsic functional connectivity within the medial temporal lobe in cognitively normal elderly. J Neurosci 35(7):3240–3247
Sun Y, Jin K, Mao XO et al (2005) Effect of aging on neuroglobin expression in rodent brain. Neurobiol Aging 26(2):275–278
Sun F, Mao X, Xie L, Greenberga DA, Jin K (2013) Neuroglobin protein is upregulated in Alzheimer’s disease. J Alzheimers Dis 36(4):659–663
Szymanski M, Wang R, Fallin MD, Bassett SS, Avramopoulos D (2010) Neuroglobin and Alzheimer’s dementia: genetic association and gene expression changes. Neurobiol Aging 31(11):1835–1842
Wang Y, Xu S, Cao Y et al (2014) Folate deficiency exacerbates apoptosis by inducing hypomethylation and resultant overexpresssion of DR4 together with altering DNMTs in Alzhemer’s disease. Int J Clin Exp Med 7(8):1945–57
Wei X, Yu Z, Cho KS et al (2011) Neuroglobin is an endogenousneuroprotectant for retinal ganglion cells against glaucomatousdamage. Am J Pathol 179:2988–1997
Yu W, Mechawar N, Krantic S, Quirion R (2010) Evidence for the involvement of apoptosis-inducing factor-mediated caspase-independent neuronal death in Alzheimer disease. Am J Pathol 176(5):2209–2218
Yuan JY, Yanker BA (2000) Apoptosis in the nervous system. Nature 407(6805):802–809
Zara S, Di Stefano A, Nasuti C et al (2011) NOS-mediated morphological and molecular modifications in rats infused with Aβ(1–40), as a model of Alzheimer’s disease, in response to a new lipophilic molecular combination codrug-1. Exp Gerontol 46(10):273–281
Zara S, Porzionato A, De Colli M et al (2013) Human carotid body neuroglobin, vascular endothelial growth factor and inducible nitric oxide synthase expression in heroin addiction. Histol Histopathol 28(7):903–911
Zhang X, Yin WK, Shi XD, Li Y (2011) Curcumin activates wnt/β-catenin signaling pathway through inhibiting the activity of gsk-3β in vitro. Eur J Pharm Sci 42(5):540–546
Zhao R, Zhang Z, Song Y, Wang D, Qi J, Wen S (2011) Implication of phosphatidylinositol-3kinas/Akt/glycogen synthase kinase-3beta pathway in ginsenoside Rb1’s attenuation of beta-amyloid-induced neurotoxicity and tau phosphorylation. J Ethnopharmacol 133(3):1109–1116
Zhu J, Tang W (2010) Cerebroprotection with recombinant neuroglobin plasmid in a rat model of focal cerebral ischemia. Neural Regen Res 5(1):52–57
Acknowledgments
This work was supported by the National Natural Science Foundation of China (NSFC: 81100948; NSFC: 81271426). We thank Deng Yu and Lin Xiao for their expert technical assistance in cell culture and immunocytochemistry.
Conflicts of Interest
The authors declare no conflicts of interest.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article

Cite this article
Li, Y., Dai, Yb., Sun, Jy. et al. Neuroglobin Attenuates Beta Amyloid-Induced Apoptosis Through Inhibiting Caspases Activity by Activating PI3K/Akt Signaling Pathway. J Mol Neurosci 58, 28–38 (2016). https://doi.org/10.1007/s12031-015-0645-z
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s12031-015-0645-z