
Abstract
VP1 protein is the immunodominant capsid protein of enterovirus 71 (EV71) which is responsible for large outbreaks of hand, foot and mouth disease. It has been reported that glucocorticoid-induced tumor necrosis factor receptor-related protein (GITR) and its ligand (GITRL) are involved in modulating both innate and adaptive immune responses. In this study, a DNA vaccine vector encoding EV71 VP1 gene and mGITRL gene (pIRES/VP1/mGITRL) was constructed. And female Balb/c mice were immunized intramuscularly with the DNA vaccine. Compared with the groups immunized with pIRES, pIRES/VP1, pIRES/mGITRL and PBS, the inoculation of pIRES/VP1/mGITRL induced a higher levels of EV71 VP1-specific antibody and specific antibody-forming cells. However, significantly higher levels of CD4+Th1, Th2 and CD8+IFN-γ+T cells were found in the pIRES/VP1/mGITRL group compared with control groups. Our results demonstrate that a novel DNA vaccine, expressing VP1 and mGITRL, could effectively elicit both humoral and cell-mediated immune responses against EV71 VP1 in mice. Thus, the mGITRL may be used as molecular adjuvant for EV71 DNA vaccine.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
Enterovirus 71 (EV71), which belongs to the enterovirus genus of the picornavirus family which also includes coxsackievirus A16 (CA16), poliovirus and echovirus, is recognized as the main pathogen of hand, foot and mouth disease (HFMD) [1, 2]. It was first isolated from California in 1969 and described in 1974 [3–5]. Since then, several outbreaks of EV71 infection were reported, including in Singapore with five deaths (2000) [2], in Tai wan with 78 deaths (1998) [6, 7] and in Malaysia with 29 deaths (1997) [8–10]. Moreover in China, a large outbreak occurred in 2008 with 126 deaths and in 2009 with 255 deaths [11, 12]. EV71 has been associated with a series of clinical diseases, including hand, foot and mouth disease, aseptic meningitis, poliomyelitis-like paralysis and possibly death [13]. Due to its tremendous impact on healthcare systems, development of EV71 vaccines is a national priority in some Asian countries.
EV71 is a small, non-enveloped, a positive-stranded RNA genome virus. It is a icosahedral capsid consisting of 60 copies each of four viral coat proteins VP1–VP4 [14]. The capsid protein VP1 is variable and contains many neutralization epitopes [15]; thus, the VP1 protein is considered to be immunodominant capsid protein and an ideal target for vaccine development [16, 17]. As the immunodominant antigen, previous study has verified that the VP1 protein play a critical role in protective efficacy.
In recent years, there have been many efforts to develop vaccine and drug candidates to defend against or treat the diseases caused by EV71 infection. Until now, DNA vaccine or nucleic acid vaccine as a third-generation vaccine has been investigated and has several advantages compared with conventional vaccine. It can express target gene of interest for a long time and induce protective immune response. Although these progresses, DNA vaccination alone often generates weak immune responses, especially in the large animals and human. Thus, molecular adjuvant containing gene encoding co-stimulatory molecule in an expressing construct has been considered as a potential facilitator and could be used to enhance immune effects of DNA vaccine [18].
Glucocorticoid-induced tumor necrosis factor receptor-related protein ligand (GITRL) is a type II transmembrane protein mainly expressed on dendritic cells (DCs) and macrophages [18–20]. The GITR/GITRL system provides a costimulatory signal to enhance CD4+ and CD8+ T cell proliferation and cytokines production [21, 22]. In addition, the functional interaction of GITR and GITRL abrogates the suppressive effects of Tregs and enhances autoreactive and alloreactive T cell responses as well as autoimmunity and graft-versus-host disease [23–27].
In this study, we attempted to use mGITRL as a molecular adjuvant to construct a new DNA vaccine encoding the EV71 VP1 gene and mGITRL gene (pIRES/VP1/mGITRL) and evaluated their roles in enhancing both humoral and cellular responses. The results showed that both humoral and cellular responses have been significantly enhanced in animals after inoculating with pIRES/VP1/mGITRL plasmid. Therefore, VP1 plus mGITRL DNA vaccine may be a potential candidate vaccine improving the protective efficacy of EV71 DNA vaccine against EV71 infection.
Materials and methods
Animals
Female Balb/c mice aged 6–8 weeks were purchased from Yangzhou University Comparative Medicine Centre and maintained with pathogen-free food and water under a clear air condition. All animal experiments performed in this study were approved by the Jiangsu University Animal Ethics and Experimentation Committee.
Plasmid constructions
The VP1 gene encoding the capsid protein VP1 of EV71 was amplified from plasmid pET-32a-VP1 with specific primers. The PCR product was cloned into multiple cloning sites (MCS) A of pIRES and designated as pIRES/VP1. The gene mGITRL encoding for the mouse GITRL protein was amplified from plasmid pMD18-T-mGITRL with specific primers. The PCR product was cloned into multiple cloning sites (MCS) B of pIRES or pIRES/VP1 and designated as pIRES/mGITRL or pIRES/VP1/mGITRL. Primers were as follows: VP1, forward primer, 5’-ATACTCGAGATGGGAGATAGGGTGGCAGA (Xho I)-3’, reverse primer, 5’-ATAACGCGTCTAGAGAGTGGTGATCGCTG (Mlu I)-3’; mGITRL, forward primer, 5’-ATCTCTAGAATGGAGGAAATGCCTTTG (Xba I)-3’, reverse primer, 5’-ATCGTCGACCTAAGAGATGAATGGTAG(Sal I)-3’.
Mice immunization
For immunization, female Balb/c mice (6 weeks old) were randomly divided into five groups with six animals per group (Table 1). The mice were immunized intramuscularly (i.m.) with pIRES, pIRES/VP1, pIRES/mGITRL, pIRES/VP1/mGITRL or phosphate-buffered saline on days 0, 14, 28. Before immunization, each mouse was injected with 100 μl of 0.25 % bupivacaine (Shanghai Zhaohui Pharmaceutical Co., Ltd, Shanghai, China) for 3 days consecutively. Mice were bled before immunization and again on day 7 after the third immunization.
Detection of anti-VP1 antibody by enzyme-linked immunosorbent assay (ELISA)
To detect the presence of anti-VP1 antibody in the sera of immunized mice, ELISA was performed. The 96-well plates were coated with 5 µg/ml VP1 protein per well in coating buffer at 4 °C overnight. Following blocking each well with 10 % NBS in PBS at 37 °C for 2 h, the serum samples were serially twofold diluted (1:200) and incubated at 37 °C for 1 h. Each well was washed and incubated with HRP-conjugated goat anti-mouse IgG (1:5000) at 37 °C for 1 h. 3,3,5,5-Tetramethylbenzidine substrate solution was added, and then, 30 µl H2SO4 (1 mol/L) was added to stop the reaction. After stopped by H2SO4, the plates were read absorbance value at 450 nm with a microplate reader (Bio-Rad, USA).
Neutralization antibody assay
Serum samples were collected and incubated at 56 °C for 30 min to inactivate the complement. Serum samples were serially diluted twofold with an initial serum dilution of 1:3 and mixed with equal volume (50 µl) of 100-TCID50 EV71 at 37 °C for 60 min. The mixtures were then added to RD cells seeded the day before. After 3-day incubation, the neutralization antibody titer was read as the highest dilution that prevented virus-induced cytopathic effects (CPE).
Detection of VP1-specific antibody-forming cells by enzyme-linked immunospot (ELISPOT) assay
To assess the number of VP1-specific antibody-forming cells in spleens from immunized mice, ELISPOT assay was performed. The 96-well MultiScreen Filter Plates (Millipore, Billerica, MA) were coated with 5 µg/ml VP1 protein per well at 4 °C for overnight. Single-cell suspensions from the spleens at 2 × 106 cells/well were added to 96-well plates for 5 h in a 5 % CO2 incubator. After three washings, each well was incubated with HRP-conjugated goat anti-mouse IgG (1:5000) at room temperature for 1 h. 3-Amino-9-ethylcarbazole dissolved in 0.1 M sodium acetate buffer containing H2O2 was added to develop spot. Counts of VP1-specific antibody-forming cells were administered by a stereomicroscope (Olympus, Japan).
Flow cytometry analysis
The mice of each group were sacrificed on day 7 after the third immunization, and splenocytes were isolated from the spleens of Balb/c mice. Single-cell suspensions from the spleens at 1 × 106 cells were stimulated in the 24-well plates with PMA (Sigma-Aldrich, USA, 50 ng/ml) and ionomycin (Enzo, Switzerland, 1 μg/ml) in the presence of monensin (Enzo, Switzerland, 2 μg/ml) for 5 h and then stained with PE-Cy5-conjugated anti-CD3mAb (eBiosciences, San Diego, CA, USA), fluorescein isothiocyanate (FITC) anti-CD8 mAb (clone 53-6.7), fixed and permeabilized using an intracellular staining kit (Invitrogen, Carlsbad, CA, USA), followed by staining with phycoerythrin (PE)-conjugated anti-IFN-γ mAb (clone XMG 1.2) and anti-IL-4 mAb (clone BVD-24G2). Immunostained cells were analyzed using a FACS Calibur (Becton Dickinson, San Jose, CA), and data were analyzed with WinMDI 2.9 software. Analysis of the Th1 and Th2 cell population was performed by gating on CD3+CD8– T cells.
Statistical analysis
All data are expressed as the mean ± SD. One-way ANOVA analysis was performed to determine whether there was an overall statistically significant change among the groups, and post-test comparison was carried out using the Bonferroni’s test. Data were analyzed with GrapPad Prism 5 Software. P values <0.05 were considered significant.
Results
Construction of recombinants pIRES/VP1, pIRES/mGITRL and pIRES/VP1/mGITRL vector
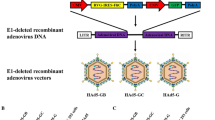
To construct the recombinant plasmid pIRES/VP1, the VP1 gene (891 bp) was amplified from plasmid pET-32a-VP1 with specific primers [28] (Fig. 1a). The PCR product was digested by restriction enzyme Xho I and Mlu I and inserted into pIRES plasmid. The full length mGITRL gene (522 bp) was cloned (Fig. 1b) and subcloned into plasmid pIRES or pIRES/VP1 by restriction enzyme Sal I and Xba I, designated as pIRES/mGITRL and pIRES/VP1/mGITRL. The recombinant plasmids were analyzed by double restriction enzyme digestion (data not shown) and sequenced by Life Technology Corporation. The results showed that recombinants were successfully constructed (Fig. 2).

Amplification of VP1 gene of EV71 and mGITRL gene by PCR. a Lane 1 which shows no band of PCR product is PCR negative control that uses water as template. Lane 2 shows a band of size 891 bp, which represents the PCR product of VP1 gene of EV71. Lane 3 shows DL 2 000 DNA marker (Takara, Dalian, China). b Lane 1 which shows no band of PCR product is PCR negative control that uses water as template. Lane 2 shows a band of size 522 bp, which represents the PCR product of the mGITRL gene. Lane 3 shows DL 2000 DNA marker

Construction of DNA vaccine pIRES/mGITRL, pIRES/VP1 and pIRES/VP1/mGITRL vector. The VP1 and mGITRL DNA fragments are inserted into eukaryotic expression vector pIRES, respectively. CMV human cytomegalovirus immediate-early promoter, IRES internal ribosome entry site, SV40 simian virus 40
mGITRL as a molecular adjuvant enhanced humoral response against EV71 VP1
To examine the effect of mGITRL as molecular adjuvant on humoral response, female Balb/c mice were immunized with pIRES, pIRES/VP1, pIRES/mGITRL, pIRES/VP1/mGITRL or PBS on days 0, 14, 28 intramuscularly (Fig. 3a). On day 7 after the third immunization, mice were sacrificed and the serum samples were collected for detect VP1-specific antibodies by ELISA (Fig. 3b). Compared with the group immunized with pIRES/VP1, mice received pIRES/VP1/mGITRL produced higher titers of VP1-specific IgG antibody. Furthermore, VP1-specific neutralizing antibodies from the immunized mice were identified by neutralization antibody assay. VP1-specific neutralizing antibodies were found in mice immunized with pIRES/VP1 and pIRES/VP1/mGITRL. The group administered with plasmid pIRES/VP1/mGITRL induced high levels of VP1-specific neutralizing antibodies than the pIRES/VP1 group (Fig. 3c).

Effect of mGITRL as adjuvant on the level of antibody production. a In vivo DNA immunization scheme. Animals are immunized intramuscularly on days 0, 14, 28. Serum samples are collected on day 7 after the final boost. Titers of total IgG (b) and neutralizing antibody (c) against EV71 are determined by ELISA and neutralization antibody assay. Results are expressed as mean ± SD (***P < 0.001; *P < 0.05)
To confirm the possible reasons of high titers VP1-specific antibody production in mice immunized with pIRES/VP1/mGITRL, the percentages of total B cells in spleens from immunized mice were analyzed using flow cytometry. Compared with the pIRES/VP1 group, no increased number of total B cells was observed in the pIRES/VP1/mGITRL immunized mice (Fig. 4a–c). We next examined antigen-specific antibody-forming cells in spleens with ELISPOT. The high level of VP1-specific antibody-forming cells was induced in the mice received recombinant vaccine containing mGITRL gene (Fig. 4d, e).

Expanded VP1-specific antibody-forming cells in mice administered with pIRES/VP1/mGITRL. a The proportions of total B cells in spleens from immunized mice with different treatments are analyzed by flow cytometry. Percentages of total B cells are indicated in representative flow-cytometric images. b Percentages of total B cells from different treatment groups are shown. c The number of total B cells in spleens from PBS, pIRES, pIRES/mGITRL, pIRES/VP1 and pIRES/VP1/mGITRL groups are shown. d Representative images of VP1-specific antibody-forming cells of spleens in PBS, pIRES, pIRES/mGITRL, pIRES/VP1 and pIRES/VP1/mGITRL groups. e The number of VP1-specific antibody-forming cells are compared between pIRES/VP1 group and pIRES/VP1/mGITRL group
mGITRL as molecular adjuvant expanded the percentages of Th1, Th2 and CD8+IFN-γ+ T cells in spleen of mice
Since GITR/GITRL act as a co-stimulating signal could enhance T cell activation and function, we sought to investigate whether the Th1, Th2 and CD8+IFN-γ+ T cells were affected by mGITRL. The expression of IFN-γ+ in CD4+ and CD8+ T cells was higher in the group intramusculary immunized with pIRES/VP1/mGITRL than mice immunized with pIRES/VP1. At the same time, the percentage of CD4+IL-4+ T cell was up-regulated by the plasmid containing mGITRL (Fig. 5b). Taken together, these data indicated that the mice immunized with pIRES/VP1/mGITRL could enhance Th1, Th2 and CD8+IFN-γ+ T cell responses.

Analysis of CD4+IFN-γ+, CD8+IFN-γ+ and CD4+IL-4+ T cells by FCM. All mice are sacrificed on day 7 after the final boost. T cells are isolated and stimulated with PMA, ionomycin and monensin in vitro. Intracellular staining for IL-4 and IFN-γ in CD4+ and CD8+ T cells is performed. a Representative flow-cytometric images of Th1, Th2 and CD8+IFN-γ+ T cells in spleens from PBS, pIRES, pIRES/mGITRL, pIRES/VP1 and pIRES/VP1/mGITRL. b Percentages of Th1, Th2 and CD8+IFN-γ+ T cells in spleens from different treatments groups are shown. Results are expressed as mean ± SD (**P < 0.01; *P < 0.05)
Discussion
Enterovirus 71 (EV71) is a 7.4-kb-long, single-stranded RNA virus in the enterovirus genus of the picornavirus family. EV71 is made up of four structural viral proteins: VP1, VP2, VP3 and VP4 [14]. VP1, VP2 and VP3 each contain 270 amino acid residues and are located in the exterior of the viral particle. VP1 plays critical roles in adsorption and the uncoating process of the virus [29]. Previous studies showed that the VP1 capsid protein can bind three human proteins including ornithine decarboxylase (ODC1), gene trap ankyrin repeat (GTAR) and KIAA0697. These interactions may contribute to the pathogenesis of EV71 disease [30]. As a major capsid protein, the VP1 protein contains a number of important neutralization sites. Thus, VP1 protein is a prime target for vaccine development [31, 32].
EV71 DNA vaccine encoding the VP1 gene has been constructed and could stimulate humoral immunity [33]. To increase immune stimulation ability of EV71 DNA vaccine, various strategies have been developed including co-expression of stimulatory molecules. In this study, mGITRL, a co-stimulatory molecule which could supply a co-stimulatory signal to enhance CD4+ and CD8+ T cell activation and cytokines production, was inserted into a eukaryotic expression vector pIRES for constructing pIRES/VP1/mGITRL which was used to enhance immune response in mice.
In this study, after administration of pIRES/VP1 and pIRES/VP1/mGITRL in mice, anti-VP1 specific antibody responses were generated. Moreover, the titers of VP1-specific antibody and the amount of VP1-specific antibody-forming cells in the animals received pIRES/VP1/mGITRL were higher than the group received with pIRES/VP1. Apart from this, the proliferation of Th2 cells was significantly enhanced in the mice immunized with pIRES/VP1/mGITRL. The differentiation of B cells into plasma cells is essential during the process of humoral immune response and need the assistant of Th2 cells. The interaction of GITR/GITRL can promote the activation and proliferation of CD4+ effector T cells. The data indicated that mGITRL played a key role in the formation of plasma cell and the production of antigen-specific antibody.
Besides of humoral immunity, the clearance of virus is associated with cellular immunity. CD8+ cytotoxic T lymphocyte (CTL) is essential for adaptive cellular immunity against viruses or other intracellular pathogens. mGITRL can promote the activation and proliferation of cytotoxic T lymphocyte (CTL) and the expansion of Th1 cells. Our data shown that Th1 and CD8+IFN-γ+ T cells were enhanced in the group immunized with pIRES/VP1/mGITR. It may suggest that mGITRL as the molecular adjuvant for EV71 DNA vaccine can promote adaptive cellular immunity.
Conclusions
In summary, our results suggested that GITRL could enhance the expansion of Th1, Th2 and CD8+IFN-γ+ T cells in the mice administrated with pIRES/VP1/mGITRL and it may promote the production of antigen-specific antibody and cell-mediated immune response further. Thus, the GITRL may be served as a promising adjuvant for EV71 DNA vaccine.
References
Solomon T, Lewthwaite P, Perera D, et al. Virology, epidemiology, pathogenesis, and control of enterovirus 71. Lancet Infect Dis. 2010;10:778–90.
Huang PN, Shih SR. Update on enterovirus 71 infection. Curr Opin Virol. 2014;5:98–104.
Schmidt NJ, Lennette EH, Ho HH. An apparently new enterovirus isolated from patients with disease of the central nervous system. J Infect Dis. 1974;129:304–9.
Blomberg J, Lycke E, Ahlfors K, et al. Letter: new enterovirus type associated with epidemic of aseptic meningitis and-or hand, foot, and mouth disease. Lancet. 1974;2:112.
Tagaya I, Takayama R, Hagiwara A. A large-scale epidemic of hand, foot and mouth disease associated with enterovirus 71 infection in Japan in 1978. Jpn J Med Sci Biol. 1981;34:191–6.
Ho M, Chen ER, Hsu KH, et al. An epidemic of enterovirus 71 infection in Taiwan. N Engl J Med. 1999;341:929–35.
Tseng FC, Huang HC, Chi CY, et al. Epidemiological survey of enterovirus infections occurring in Taiwan between 2000 and 2005: analysis of sentinel physician surveillance data. J Med Virol. 2007;79:1850–60.
Chan LG, Parashar UD, Lye MS, et al. Deaths of children during an outbreak of hand, foot, and mouth disease in Sarawak, Malaysia: clinical and pathological characteristics of the disease. Clin Infect Dis. 2000;31:678–83.
Ooi MH, Wong SC, Podin Y, et al. Human enterovirus 71 disease in Sarawak, Malaysia: a prospective clinical, virological, and molecular epidemiological study. Clin Infect Dis. 2007;44:646–56.
Ooi MH, Wong SC, Mohan A, et al. Identification and validation of clinical predictors for the risk of neurological involvement in children with hand, foot, and mouth disease in Sarawak. BMC Infect Dis. 2009;9:3.
Yang F, Ren L, Xiong Z, et al. Enterovirus 71 outbreak in the People’s Republic of China in 2008. J Clin Microbiol. 2009;47:2351–2.
Zhang Y, Tan XJ, Wang HY, et al. An outbreak of hand, foot, and mouth disease associated with subgenotype C4 of human enterovirus 71 in Shandong, China. J Clin Virol. 2009;44:262–7.
Bible JM, Pantelidis P, Chan PK, Tong CY. Genetic evolution of enterovirus 71: epidemiological and pathological implications. Rev Med Virol. 2007;17:371–9.
Zhou SL, Ying XL, Han X, et al. Characterization of the enterovirus 71 VP1 protein as a vaccine candidate. J Med Virol. 2015;87:256–62.
Oberste MS, Maher K, Kilpatrick DR, Pallansch MA. Molecular evolution of the human enteroviruses: correlation of serotype with VP1 sequence and application to picornavirus classification. J Virol. 1999;73:1941–8.
Foo DG, Alonso S, Chow VT, Poh CL. Passive protection against lethal enterovirus 71 infection in newborn mice by neutralizing antibodies elicited by a synthetic peptide. Microbes Infect. 2007;9:1299–306.
Wu CN, Lin YC, Fann C, et al. Protection against lethal enterovirus 71 infection in newborn mice by passive immunization with subunit VP1 vaccines and inactivated virus. Vaccine. 2001;20:895–904.
Svanholm C, Lowenadler B, Wigzell H. Amplification of T-cell and antibody responses in DNA-based immunization with HIV-1 Nef by co-injection with a GM-CSF expression vector. Scand J Immunol. 1997;46(3):298–303.
Placke T, Kopp HG, Salih HR. Glucocorticoid-induced TNFR-related (GITR) protein and its ligand in antitumor immunity: functional role and therapeutic modulation. Clin Dev Immunol. 2010;2010:239083.
Nocentini G, Riccardi C. GITR: a modulator of immune response and inflammation. Adv Exp Med Biol. 2009;647:156–73.
Kohm AP, Williams JS, Miller SD. Cutting edge: ligation of the glucocorticoid-induced TNF receptor enhances autoreactive CD4+ T cell activation and experimental autoimmune encephalomyelitis. J Immunol. 2004;172:4686–90.
Ronchetti S, Zollo O, Bruscoli S, et al. GITR, a member of the TNF receptor superfamily, is costimulatory to mouse T lymphocyte subpopulations. Eur J Immunol. 2004;34:613–22.
Stephens GL, McHugh RS, Whitters MJ, et al. Engagement of glucocorticoid-induced TNFR family-related receptor on effector T cells by its ligand mediates resistance to suppression by CD4+CD25+ T cells. J Immunol. 2004;173:5008–20.
Shimizu J, Yamazaki S, Takahashi T, et al. Stimulation of CD25+CD4+ regulatory T cells through GITR breaks immunological self-tolerance. Nat Immunol. 2002;3:135–42.
McHugh RS, Whitters MJ, Piccirillo CA, et al. CD4+CD25+ immunoregulatory T cells: gene expression analysis reveals a functional role for the glucocorticoid-induced TNF receptor. Immunity. 2002;16:311–23.
Muriglan SJ, Ramirez-Montagut T, Alpdogan O, et al. GITR activation induces an opposite effect on alloreactive CD4+ and CD8+ T cells in graft-versus-host disease. J Exp Med. 2004;200:149–57.
Wang S, Shi Y, Yang M, et al. Glucocorticoid-induced tumor necrosis factor receptor family-related protein exacerbates collagen-induced arthritis by enhancing the expansion of Th17 cells. Am J Pathol. 2012;180:1059–67.
Mao ZW, Liu HB, Zheng D, et al. Construction, expression and immunogenicity of EV71 multiepitope-mGITRL eukaryotic plasmid. Xi Bao Yu Fen Zi Mian Yi Xue Za Zhi. 2012;28:797–800.
Li C, Wang H, Shih SR, et al. The efficacy of viral capsid inhibitors in human enterovirus infection and associated diseases. Curr Med Chem. 2007;14:847–56.
Yeo WM, Chow VT. The VP1 structural protein of enterovirus 71 interacts with human ornithine decarboxylase and gene trap ankyrin repeat. Microb Pathog. 2007;42:129–37.
Lyu K, Wang GC, He YL, et al. Crystal structures of enterovirus 71 (EV71) recombinant virus particles provide insights into vaccine design. J Biol Chem. 2015;290:3198–208.
Kung YA, Hung CT, Liu YC, et al. Update on the development of enterovirus 71 vaccines. Expert Opin Biol Ther. 2014;14:1455–64.
Tung WS, Bakar SA, Sekawi Z, Rosli R. DNA vaccine constructs against enterovirus 71 elicit immune response in mice. Genet Vaccines Ther. 2007;5:6.
Acknowledgments
This work was supported by the National Natural Science Foundation of China (Grant No. 31100648, 81072453), Specialized Project for Clinical Medicine of Jiangsu Province (Grant No. BL2014065), Specialized Research Fund for the Doctoral Program of Higher Education (Grant No. 20133227110008), Health Department Foundation of Jiangsu Province (Grant No. Z201312), Science and technology support program (Social Development) of Zhenjiang (Grant No. SH2014039), Graduate Student Research and Innovation Program of Jiangsu Province (Grant No. CXZZ13_0700, KYLX_1074), Jiangsu Province “333” Project and Priority Academic Program Development of Jiangsu Higher Education Institutions.
Conflict of interest
The authors declare that they have no competing interests.
Author information
Authors and Affiliations
Corresponding authors
Additional information
Jing Yuan, Xinyi Tang and Kai Yin have contributed equally to this work.
Rights and permissions
About this article

Cite this article
Yuan, J., Tang, X., Yin, K. et al. GITRL as a genetic adjuvant enhances enterovirus 71 VP1 DNA vaccine immunogenicity. Immunol Res 62, 81–88 (2015). https://doi.org/10.1007/s12026-015-8637-1
Published:
Issue Date:
DOI: https://doi.org/10.1007/s12026-015-8637-1