
Abstract
To understand the mechanism by which early insulin therapy improves insulin sensitivity in type 2 diabetes, we investigated endoplasmic reticulum (ER) stress in the liver of type 2 diabetic rats. A high fat diet plus a low dose of streptozotocin (STZ) in Sprague–Dawley (SD) rats was implemented to create an animal model mimicking diabetes. After 3 weeks of insulin treatment, the rats were examined for insulin sensitivity and ER stress in the liver. To investigate insulin sensitivity within the liver, serine phosphorylation of IRS-1 (Ser307) and Akt (Ser473) and expression of gluconeogenic genes, PEPCK and G6Pase, were tested. Protein levels of ER stress markers, such as immunoglobulin binding protein (Bip), inositol-requiring protein 1 alpha (IRE1α), and unspliced and spliced x-box binding protein-1 (XBP-1), were determined to assess ER stress. In the diabetic (DM) group, IRS-1 phosphorylation was increased (P < 0.05), Akt phosphorylation was reduced (P < 0.05), expression of PEPCK and G6Pase was elevated (P < 0.05), and ER stress markers were up-regulated (P < 0.05) relative to the non-diabetic rats. In the insulin (INS) therapy group, all of aforementioned changes were attenuated or reversed (P < 0.05). In addition, c-Jun N-terminal kinase (JNK) activity and SREBP-1 expression were decreased (P < 0.05). Adipose tissue mass was increased (P < 0.05). These data suggest that short-term insulin therapy relieved ER stress and enhanced insulin sensitivity in the liver of diabetic rats. The mechanism is likely related to fat redistribution from liver to adipose tissue. These cellular and molecular responses may represent a mechanism for improvement of insulin sensitivity in type 2 diabetic rats by insulin therapy.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
Our recent studies demonstrate that short-term intensive insulin therapy for 2–3 weeks improves systemic and hepatic insulin sensitivity in type 2 diabetes [1–3], which are consistent with those reported by other laboratories that insulin treatment enhances insulin sensitivity [4–7]. However, the mechanism of improved insulin sensitivity is not clear. Several possible mechanisms may be involved, such as reduction of oxidative stress from hyperglycemia, attenuation of lipotoxicity in liver and muscle, and inhibition of inflammatory responses [8–11]. Ectopic lipid deposition induces insulin resistance through endoplasmic reticulum (ER) stress and c-Jun N-terminal kinase (JNK) activation [12]. Our recent study shows that insulin sensitivity is enhanced in the liver with insulin therapy [2].
Hepatic steatosis is associated with obesity and involved in the pathogenesis of insulin resistance [13, 14]. Lipid accumulation in the liver is related to adipose tissue dysfunction in fat storage. Adipocytes have a high level of insulin receptor and glucose transporter 4 (GLUT4). In response to insulin, adipocytes exhibit an increase in glucose uptake through GLUT4 translocation. Insulin also induces triglyceride (TG) synthesis in adipocytes through expression of enzymes for TG biosynthesis. Under insulin therapy, the adipocyte response may induce lipid transfer from the liver to adipose tissue, reducing ectopic fat deposition in the obese condition. Reduction in hepatic steatosis may contribute to insulin sensitivity from insulin therapy.
Lipid may induce insulin resistance in the liver through ER stress. ER stress induces JNK activation and inhibits insulin signal transduction. Serine phosphorylation of insulin receptor substrate 1 (IRS-1) at Ser307 may link JNK activation to the signaling inhibition of insulin [12]. The JNK-IRS-1 pathway is a molecular mechanism of hepatic insulin resistance in ER stress that results from liver lipid deposition. Reduction of lipid accumulation in the liver may attenuate ER stress and restore insulin sensitivity by reducing JNK activation. This possibility is supported by an early study from our lab showing a reduction in hepatic triglyceride content by insulin therapy in diabetic rats [2].
In this study, we examined the ER stress response and JNK activation in the liver of obese diabetic rats after insulin therapy. Our data suggest that insulin therapy reduced ER stress and improved insulin sensitivity in the liver.
Results
Insulin signaling pathway in the liver
In this study, type 2 diabetic rats were generated in SD rats by a high fat diet (HFD) in combination with a single injection of low dose streptozotocin (STZ) (40 mg/kg) as reported in our previous study [2]. Diabetic rats exhibited insulin resistance and hyperglycemia [2, 15], which resemble type 2 diabetes in humans. This animal model is widely used in the study of type 2 diabetes by many groups [16–19]. To investigate the mechanism of insulin therapy, we treated the diabetic rats with insulin for 3 weeks. Insulin sensitivity was improved by the therapy as reported in our previous publication [2]. Here, we report that the visceral adipose tissue mass was significantly increased in the insulin therapy group (INS) in comparison to the diabetic group (DM) (22.40 ± 3.63 g over 11.96 ± 2.63 g, P < 0.05).
To investigate the molecular mechanism of insulin sensitivity, we examined the insulin signaling pathway in liver. Liver plays a critical role in the control of systemic insulin sensitivity. Liver controls plasma insulin levels by removal of at least 50% of insulin at the portal vein. Liver determines fasting blood glucose through gluconeogenesis. When the liver suffers insulin resistance, insulin clearance is reduced leading to hyperinsulinemia; hepatic glucose production will be enhanced contributing to hyperglycemia. In this study, we examined liver insulin sensitivity by measuring the activities of IRS-1 and Akt (Protein kinase B, PKB) in the insulin signaling pathway. IRS-1 Ser307 phosphorylation (a marker of insulin resistance) and the Akt Ser473 phosphorylation (a marker of insulin action) were quantified in the liver by Western blotting after insulin challenge [20]. In the DM group, the IRS-1 phosphorylation was increased (P < 0.05, Fig. 1a), and the Akt phosphorylation was reduced (P < 0.05, Fig. 1b). In the INS group, the alterations were reversed (Fig. 1a, b), suggesting an improvement in the insulin signaling activity. The data provide molecular evidence for improvement of liver insulin sensitivity in the INS group.

Insulin therapy improved insulin sensitivity in the liver of diabetic rats. On day 3 following insulin therapy, liver samples were collected. Serine phosphorylation of IRS-1 and Akt were examined after insulin challenge for 15 min in the rats. Expression of PEPCK and G6Pase were examined in the liver of rats without insulin challenge. a IRS-1 phosphorylation (Ser307). b Akt phosphorylation (Ser473). c Expression of PEPCK. d Expression of G6Pase. IRS-1, β-actin, and GAPDH were used as a control for protein loading. The relative signal levels were determined by densitometry and normalized with the loading control. Each bar represents mean ± SD (n = 4–6 rats). *P < 0.05 vs. NC; # P < 0.05 vs. DM. NC normal control rats; DM diabetic rats with no therapy; INS diabetic rats treated with insulin
Phosphoenolpyruvate carboxykinase (PEPCK) and glucose 6 phosphatase (G6Pase), two rate-limiting enzymes of hepatic gluconeogenesis, are negatively regulated by insulin at the transcriptional level [21]. Under insulin resistance, their expression is enhanced, leading to elevated hepatic gluconeogenesis. We determined expression of the two enzymes in liver lysates by Western blot. In the DM group, expression of both proteins was increased (P < 0.05 for both; Fig. 1c, d). In the INS group, the increases in protein expression were eliminated in both enzymes (P < 0.05 for both; Fig. 1c, d). These data provide another line of molecular evidence for the improved insulin sensitivity in the liver of the INS group.
Reduced JNK activity in the INS group
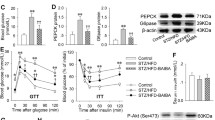
IRS-1 serine 307 is phosphorylated by the serine kinase JNK [22]. The phosphorylation of IRS-1 at serine 307 is a marker of insulin resistance. In the DM group, increased phosphorylation of IRS-1 suggests activation of JNK. To test this possibility, we examined phosphorylation status of the JNK substrate, c-JUN, by Western blot. We found that c-JUN phosphorylation in the liver was increased in the DM group (P < 0.05) but decreased in the INS group (P < 0.05; Fig. 2). The data suggest that JNK activity was increased in the DM group, and the increase was attenuated by the insulin therapy. The changes in JNK activity were not due to an alteration in the protein levels of JNK, as JNK protein was unchanged among the three groups (Fig. 2). Therefore, the reduced JNK activity may contribute to the molecular mechanism of improved insulin sensitivity in the INS group.

Insulin therapy reduced JNK activity in the liver of diabetic rats. JNK activity was determined by phosphorylation status of its substrate c-JUN in a Western blot. To normalize for JNK activity, JNK proteins were examined in cell lysates. Anti-JNK antibody that detects both JNK1 and JNK2 was used. The relative signal levels were determined by densitometry. Each bar represents the mean ± SD (n = 4–6). *P < 0.05 vs. NC; # P < 0.05 vs. DM. NC normal control rats; DM diabetic rats with no therapy; INS diabetic rats treated with insulin
Reduced ER stress in the INS group
Endoplasmic reticulum stress is a risk factor for insulin resistance in obesity. ER stress is a result of lipotoxicity in obesity and impairs insulin signaling by activation of the serine kinase JNK [12]. The reduction in JNK activity suggests a low level of ER stress in the INS group. To test this possibility, we examined expression of immunoglobulin binding protein (Bip), an ER stress marker protein [23]. In the DM group, Bip protein was increased 55% relative to the non-diabetic control group (P < 0.05, Fig. 3a). The increase in Bip was markedly reduced in the INS group (P < 0.05, Fig. 3a). Furthermore, we examined three other ER stress marker proteins, including inositol-requiring protein 1 alpha (IRE1α) and spliced and unspliced x-box binding protein-1 (XBP-1s and XBP-1u). These proteins were all significantly increased in the DM group, and reduced in the INS group (P < 0.05 for all, Fig. 3b–d). The data suggest that ER stress occurred in the liver of diabetic rats, and its level was reduced by the insulin therapy. The reduction in ER stress may be responsible for the decreased JNK activity in the INS group.

Insulin therapy down-regulated the ER stress response in the liver of diabetic rats. Protein concentrations of ER stress markers were examined in the liver by Western blot to determine ER stress. Liver samples were collected on day 3 after insulin therapy. a Expression of Bip. b Expression of IRE1α. c Expression of spliced XBP-1. The signal shown in the XBP-1s immunoblot corresponds to the 54 kDa protein encoded by the sXBP-1 mRNA. d Unspliced XBP-1. The signal shown in the XBP-1s immunoblot corresponds to the 31 kDa protein encoded by the XBP-1u mRNA. β-actin protein is used as the control for protein loading. The relative signal levels were determined by densitometry. Each bar represents mean ± SD (n = 4–6). *P < 0.05 vs. NC; # P < 0.05 vs. DM. NC normal control rats; DM diabetic rats with no therapy; INS diabetic rats treated with insulin
Attenuation of sterol regulatory element binding protein 1 (SREBP-1) expression
SREBP-1 is a transcription factor that plays a crucial role in the regulation of triglyceride synthesis in the liver and adipose tissue [24, 25]. SREBP-1 has two forms, cytoplasmic SREBP-1 and nuclear SREBP-1 (nSREBP-1). In response to sterol depletion or insulin, SREBP-1 is cleaved and translocated into the nucleus to form nSREBP-1 [26–28]. nSREBP-1 protein determines the transcriptional activity of several lipogenic genes. In the DM group, SREBP-1 activity was elevated in both cytoplasmic and nuclear forms in the liver (Fig. 4a, b). The elevation was associated with an increase in liver TG content [24, 29]. In the INS group, the liver TG content was reduced [2]. It is possible that the reduction in TG is due to a decrease in SREBP-1 activity. To test this possibility, we examined SREBP-1 in the liver of INS group. Levels of both SREBP-1 forms were reduced in the INS group (P < 0.05 for both, Fig. 4a, b). The data suggest that the SREBP-1 reduction may contribute to the mechanism by which insulin therapy decreases liver TG in the INS group.

Insulin therapy down-regulated SREBP-1 in the liver of diabetic rats. a Expression of SREBP-1. b Expression of nSREBP-1. β-actin and SP3 protein were used as controls for protein loading. The relative signal levels were determined by densitometry. Each bar represents mean ± SD (n = 4–6). *P < 0.05 vs. NC; # P < 0.05 vs. DM. NC normal control rats; DM diabetic rats with no therapy; INS diabetic rats treated with insulin
Discussion
Our results suggest that ER stress is reduced in the liver of diabetic rats by insulin therapy. Insulin therapy improves glycogen synthesis and reduces inflammation in skeletal muscle [2, 30]. These effects may explain insulin sensitivity due to insulin therapy. However, insulin may also act in the liver, as liver insulin sensitivity is improved by insulin therapy leading to a reduction in hepatic glucose production [31]. The mechanism of improved insulin sensitivity through insulin therapy is not clear. It is unknown if liver lipotoxicity is attenuated by insulin therapy. We address these issues by examining liver ER stress in diabetic rats after insulin therapy. Liver ER stress is an important mechanism of insulin resistance in obese mice [12, 32] and humans [25, 33, 34]. Here, we report that liver ER stress is reduced by insulin therapy. This conclusion is supported by a reduction in ER stress markers, such as BIP, IRE1α, XBP-1s, and XBP-1u. Inhibition of ER stress may contribute to improved insulin sensitivity in the liver. Excessive lipid accumulation may induce ER stress in the liver leading to insulin resistance [9, 29]. We previously reported that the liver fat content was reduced by insulin therapy in diabetic rats [2]. Decreased lipid accumulation may account for the inhibition of ER stress in the liver.
Our data suggest that JNK activity is reduced in the liver of diabetic rats by insulin therapy. JNK may inhibit insulin signaling by serine phosphorylation of IRS-1. Moreover, ER stress is a major risk factor for JNK activation in the liver of obese mice [32]. In this study, we demonstrate that liver JNK activity is reduced in diabetic rats by insulin therapy. Consistently, IRS-1 serine phosphorylation at Ser307 is reduced by insulin therapy. The reduction in JNK activity and IRS-1 serine phosphorylation provides a mechanism for the improved liver insulin sensitivity in the INS group.
Our data suggest that insulin therapy may induce the transfer of fat from the liver to adipose tissue. In our early studies, liver TG content was reduced in diabetic rats by insulin therapy [2]. Insulin also reduced TG in the blood. Here, we report that these reductions are associated with an increase in visceral fat mass. As changes in visceral fat mass are observed in the absence of body weight alterations, the data suggest that liver TG is transferred into visceral adipose tissue. It is unlikely that the reduction in visceral fat mass is from a change in energy balance as the total body weight was not changed by insulin therapy [2]. Thus, the only possibility is fat redistribution from liver to adipose tissue. Adipocytes express a high level of insulin receptor and synthesize TG in response to insulin. Insulin may stimulate adipose tissue expansion through a couple of mechanisms, such as induction of glucose deposition into adipocytes, stimulation of TG biosynthesis, and expression of pro-angiogenic factor (VEGF) in adipocytes [35, 36]. The angiogenic activity of insulin may contribute to improvement of the microenvironment for adipocytes in the adipose tissue. Our previous studies suggest that the microenvironment is critical to maintain adipocyte function [37]. A hypoxia response may occur in the adipose tissue in obesity from an alteration in the microenvironment. Hypoxia suppresses multiple functions of adipocytes, such as TG storage and adiponectin expression [38, 39]. A failure in local angiogenic function may contribute to the adipose tissue hypoxia [40]. Therefore, insulin may promote fat tissue expansion through intracellular and extracellular mechanisms.
Reduction in SREBP-1 expression provides a mechanism for the improvement of hepatic steatosis by insulin therapy. Hepatic lipogenesis depends on the transcription factor SREBP-1c [29]. Overexpression of SREBP-1c induces expression of lipogenic genes and increases lipid deposition [41]. This study suggests that insulin therapy may reduce hepatic lipogenesis by down-regulating SREBP-1 expression as well as its activation. The SREBP-1 analysis was conducted 3 days after the insulin therapy to allow washout of exogenous insulin. In this condition, the acute effect of exogenous insulin is excluded [42].
In conclusion, this study suggests that liver may be a target of insulin for improvement of whole body insulin sensitivity with insulin therapy. We provide molecular evidence for the improvement of insulin sensitivity in the liver of diabetic rats, including inhibition of ER stress, enhanced insulin signaling and suppression of gluconeogenic genes in the liver. These effects are likely due to lipid redistribution from liver to adipose tissue in response to insulin, which was observed in the absence of changes in body weight.
Materials and methods
Type 2 diabetic rat model
All animal experiments were approved by the Animal Care and Use Committee of the Sun Yat-Sen University. The type 2 diabetic rat model was developed as previously described [2, 17]. All procedures were performed in accordance with the principles of laboratory animal care (NIH publication no. 85–23, revised 1985). Forty-two male SD rats (8 weeks old, approximately 200 g in body weight) were purchased from the Central Animal Facility of Southern Medical University (China) and maintained in the animal facility of Sun Yat-Sen University. The animal room was conditioned at 20–25°C with a 12-h light–dark cycle. The rats were allowed free access to water and diet. After acclimatization for 1 week, the rats were randomly divided into the NC group (12 rats) and HFD group (30 rats). The control rats were fed the normal diet (10% calorie in fat, 64% in carbohydrate, and 26% in protein). The HFD had 56% calories from fat, 32% from carbohydrate, and 14% from protein. After 5 weeks on the HFD, the HFD group rats were injected intraperitoneally with a low dose STZ (40 mg/kg, Sigma, USA) after an overnight fast. The NC rats were fasted in an identical manner and injected with the same volume of citrate buffered saline. Blood glucose was measured 3 days after STZ injection. Rats with blood glucose ≥16.7 mmol/l in the fed condition were considered type 2 diabetes. Weight- and glucose-matched diabetic rats were randomly divided into two groups: DM group (untreated diabetic rats) and INS group (treated with NPH insulin for 3 weeks initiated from the 3rd day of STZ injection). Insulin was delivered daily by subcutaneous injection (6–8 U/day) for 3 weeks. Insulin dosage was based on a week long preliminary study using insulin within a range from 4 mmol/l to 8 mmol/l. The NC group and DM group were injected with saline. The diets remained unchanged during the insulin therapy. Blood glucose was measured in the tail vein blood every 3 days at 09:00–10:00 am with a portable glucometer (Roche, USA).
Insulin challenge and tissue sampling
On day 3 after insulin therapy, rats in each group were dived into two subgroups for analysis of insulin sensitivity in the liver. One subgroup (half number of rats in either DM or INS groups) was challenged with insulin (8 U/kg body weight, i.p.) to activate the insulin signaling pathway; the other subgroup was given vehicle. Liver was collected 15 min after the insulin or vehicle injection and frozen in liquid nitrogen immediately after isolation, and then stored at −80°C till experiments for insulin signaling and protein expression assays. Femoral artery blood was collected and used in preparation of serum. Serum samples were stored at −30°C before biochemical assays.
Western blot and Immunoprecipitation (IP)
Whole cell lysates were made from liver (~200 mg) by homogenization in ice-cold lysis buffer (Cell Signaling Technologies, USA). Protein concentration was determined using a bicinchoninic acid (BCA) protein assay (Pierce chemical, USA). The samples were resolved by SDS-PAGE (10%) and then transferred onto polyvinylidene fluoride membranes (0.45 μm; Millipore, USA). After preblotting in blocking buffer for 2 h at room temperature, the membranes were incubated overnight at 4°C with specific antibodies to Bip, XBP-1, PEPCK, G6Pase, IRS-1, SREBP-1 (Santa Cruz, USA), phospho c-Jun (p-c-Jun, Ser63), IRE1α, phospho IRS-1 (Ser307), phospho AKT (Ser473), JNK (Cell Signaling Technologies, USA), COOH terminus XBP-1 (Biolegend, USA), Sp3 and GAPDH (eBioscience, USA), and β-actin (Boster, China). After being washed four times in TBST buffer, the membranes were incubated with an HRP-conjugated secondary antibody for 1 h at room temperature and then treated for chemiluminescence (Santa Cruz, USA). Band intensities were quantified by densitometry. In the immunoprecipitation assay, IRS-1 protein was pulled down with an IRS-1 antibody and Ser307 phosphorylation was determined by Western blot.
Statistical analysis
Data are presented as means ± SEM. Statistical analysis was conducted using one-way ANOVA followed by a Student–Newman–Keuls test. Significance was defined at P < 0.05.
Abbreviations
- ER stress:
-
Endoplasmic reticulum stress
- IRS-1:
-
Insulin receptor substrate 1
- Akt:
-
Protein kinase B
- PEPCK:
-
Phosphoenolpyruvate carboxykinase
- G6Pase:
-
Glucose-6-phosphatase
- JNK:
-
c-Jun N-terminal kinase
- SREBP-1:
-
Sterol regulatory element binding protein 1
- TG:
-
Triglyceride
References
J. Weng, Y. Li, W. Xu et al., Effect of intensive insulin therapy on beta-cell function and glycaemic control in patients with newly diagnosed type 2 diabetes: a multicentre randomised parallel-group trial. Lancet 371, 1753–1760 (2008)
Y. Bi, W.P. Sun, X. Chen et al., Effect of early insulin therapy on nuclear factor kappaB and cytokine gene expressions in the liver and skeletal muscle of high-fat diet, streptozotocin-treated diabetic rats. Acta Diabetol 45, 167–178 (2008)
Y. Li, W. Xu, Z. Liao et al., Induction of long-term glycemic control in newly diagnosed type 2 diabetic patients is associated with improvement of beta-cell function. Diabetes Care 27, 2597–2602 (2004)
M.J. Gormley, D.R. Hadden, R. Woods et al., One month’s insulin treatment of type II diabetes: the early and medium-term effects following insulin withdrawal. Metabolism 35, 1029–1036 (1986)
W.J. Andrews, B. Vasquez, M. Nagulesparan et al., Insulin therapy in obese, non-insulin-dependent diabetes induces improvements in insulin action and secretion that are maintained for two weeks after insulin withdrawal. Diabetes 33, 634–642 (1984)
W.T. Garvey, J.M. Olefsky, J. Griffin et al., The effect of insulin treatment on insulin secretion and insulin action in type II diabetes mellitus. Diabetes 34, 222–234 (1985)
M. Laakso, M. Uusitupa, J. Takala et al., Effects of hypocaloric diet and insulin therapy on metabolic control and mechanisms of hyperglycemia in obese non-insulin-dependent diabetic subjects. Metabolism 37, 1092–1100 (1988)
M.C. Bravi, A. Armiento, O. Laurenti et al., Insulin decreases intracellular oxidative stress in patients with type 2 diabetes mellitus. Metabolism 55, 691–695 (2006)
H. Kaneto, Y. Nakatani, D. Kawamori et al., Role of oxidative stress, endoplasmic reticulum stress, and c-Jun N-terminal kinase in pancreatic beta-cell dysfunction and insulin resistance. Int. J. Biochem. Cell Biol. 37, 1595–1608 (2005)
L. Juurinen, M. Tiikkainen, A.M. Hakkinen et al., Effects of insulin therapy on liver fat content and hepatic insulin sensitivity in patients with type 2 diabetes. Am. J. Physiol. Endocrinol. Metab. 292, E829–E835 (2007)
D. LeRoith, V. Fonseca, Vinik A Metabolic memory in diabetes—focus on insulin. Diabetes Metab. Res. Rev. 21, 85–90 (2005)
U. Ozcan, Q. Cao, E. Yilmaz et al., Endoplasmic reticulum stress links obesity, insulin action, and type 2 diabetes. Science 306, 457–461 (2004)
A. Garg, A. Misra, Hepatic steatosis, insulin resistance, and adipose tissue disorders. J. Clin. Endocrinol. Metab. 87, 3019–3022 (2002)
A. Seppala-Lindroos, S. Vehkavaara, A.M. Hakkinen et al., Fat accumulation in the liver is associated with defects in insulin suppression of glucose production and serum free fatty acids independent of obesity in normal men. J. Clin. Endocrinol. Metab. 87, 3023–3028 (2002)
Y. Bi, M. Cai, H. Liang et al., Increased carnitine palmitoyl transferase 1 expression and decreased sterol regulatory element-binding protein 1c expression are associated with reduced intramuscular triglyceride accumulation after insulin therapy in high-fat-diet and streptozotocin-induced diabetic rats. Metabolism 58, 779–786 (2009)
M.J. Reed, K. Meszaros, L.J. Entes et al., A new rat model of type 2 diabetes: the fat-fed, streptozotocin-treated rat. Metabolism 49, 1390–1394 (2000)
K. Srinivasan, B. Viswanad, L. Asrat et al., Combination of high-fat diet-fed and low-dose streptozotocin-treated rat: a model for type 2 diabetes and pharmacological screening. Pharmacol. Res. 52, 313–320 (2005)
M. Sugano, H. Yamato, T. Hayashi et al., High-fat diet in low-dose-streptozotocin-treated heminephrectomized rats induces all features of human type 2 diabetic nephropathy: a new rat model of diabetic nephropathy. Nutr. Metab. Cardiovasc. Dis. 16, 477–484 (2006)
K. Sahin, M. Onderci, M. Tuzcu et al., Effect of chromium on carbohydrate and lipid metabolism in a rat model of type 2 diabetes mellitus: the fat-fed, streptozotocin-treated rat. Metabolism 56, 1233–1240 (2007)
J.E. Pessin, A.R. Saltiel, Signaling pathways in insulin action: molecular targets of insulin resistance. J. Clin. Invest. 106, 165–169 (2000)
R.W. Hanson, L. Reshef, Regulation of phosphoenolpyruvate carboxykinase (GTP) gene expression. Annu. Rev. Biochem. 66, 581–611 (1997)
J. Hirosumi, G. Tuncman, L. Chang et al., A central role for JNK in obesity and insulin resistance. Nature 420, 333–336 (2002)
A.S. Lee, The ER chaperone and signaling regulator GRP78/BiP as a monitor of endoplasmic reticulum stress. Methods 35, 373–381 (2005)
J.D. Horton, J.L. Goldstein, M.S. Brown, SREBPs: activators of the complete program of cholesterol and fatty acid synthesis in the liver. J. Clin. Invest. 109, 1125–1131 (2002)
G. Boden, X. Duan, C. Homko et al., Increase in endoplasmic reticulum stress-related proteins and genes in adipose tissue of obese, insulin-resistant individuals. Diabetes 57, 2438–2444 (2008)
M.S. Brown, J.L. Goldstein, A proteolytic pathway that controls the cholesterol content of membranes, cells, and blood. Proc. Natl. Acad. Sci. USA 96, 11041–11048 (1999)
M.S. Brown, J.L. Goldstein, The SREBP pathway: regulation of cholesterol metabolism by proteolysis of a membrane-bound transcription factor. Cell 89, 331–340 (1997)
M.S. Brown, J. Ye, R.B. Rawson et al., Regulated intramembrane proteolysis: a control mechanism conserved from bacteria to humans. Cell 100, 391–398 (2000)
H.L. Kammoun, H. Chabanon, I. Hainault et al., GRP78 expression inhibits insulin and ER stress-induced SREBP-1c activation and reduces hepatic steatosis in mice. J. Clin. Invest. 119, 1201–1215 (2009)
T. Pratipanawatr, K. Cusi, P. Ngo et al., Normalization of plasma glucose concentration by insulin therapy improves insulin-stimulated glycogen synthesis in type 2 diabetes. Diabetes 51, 462–468 (2002)
S. Matthaei, M. Stumvoll, M. Kellerer et al., Pathophysiology and pharmacological treatment of insulin resistance. Endocr. Rev. 21, 585–618 (2000)
U. Ozcan, E. Yilmaz, L. Ozcan et al., Chemical chaperones reduce ER stress and restore glucose homeostasis in a mouse model of type 2 diabetes. Science 313, 1137–1140 (2006)
S.K. Das, W.S. Chu, A.K. Mondal et al., Effect of pioglitazone treatment on endoplasmic reticulum stress response in human adipose and in palmitate-induced stress in human liver and adipose cell lines. Am. J. Physiol. Endocrinol. Metab. 295, E393–E400 (2008)
N.K. Sharma, S.K. Das, A.K. Mondal et al., Endoplasmic reticulum stress markers are associated with obesity in nondiabetic subjects. J. Clin. Endocrinol. Metab. 93, 4532–4541 (2008)
G.J. Hausman, Richardson RL Adipose tissue angiogenesis. J. Anim. Sci. 82, 925–934 (2004)
Y. Cao, Angiogenesis modulates adipogenesis and obesity. J. Clin. Invest. 117, 2362–2368 (2007)
J. Ye, Emerging role of adipose tissue hypoxia in obesity and insulin resistance. Int. J. Obes. (Lond) 33, 54–66 (2009)
J. Ye, Z. Gao, J. Yin et al., Hypoxia is a potential risk factor for chronic inflammation and adiponectin reduction in adipose tissue of ob/ob and dietary obese mice. Am. J. Physiol. Endocrinol. Metab. 293, E1118–E1128 (2007)
J. Yin, Z. Gao, Q. He et al., Role of hypoxia in obesity-induced disorders of glucose and lipid metabolism in adipose tissue. Am. J. Physiol. Endocrinol. Metab. 296, E333–E342 (2009)
C. Pang, Z. Gao, J. Yin et al., Macrophage infiltration into adipose tissue may promote angiogenesis for adipose tissue remodeling in obesity. Am. J. Physiol. Endocrinol. Metab. 295, E313–E322 (2008)
H. Shimano, N. Yahagi, M. Amemiya-Kudo et al., Sterol regulatory element-binding protein-1 as a key transcription factor for nutritional induction of lipogenic enzyme genes. J. Biol. Chem. 274, 35832–35839 (1999)
M. Foretz, C. Guichard, P. Ferre et al., Sterol regulatory element binding protein-1c is a major mediator of insulin action on the hepatic expression of glucokinase and lipogenesis-related genes. Proc. Natl. Acad. Sci. USA 96, 12737–12742 (1999)
Acknowledgments
This study was supported by grants from the National Basic Research Program of China (973 program: No. 2006CB503902), the National Natural Science Foundation of China Grant Award (30800539), the Ministry of Health of the People’s Republic of China for Key Projects of Clinical Disciplines of Hospitals Affiliated to Ministry of Health (2007–2009), and the Research Foundation of Science and Technology Plan Project in Guangdong Province of China (Grant No. 2006B60101066).
Author information
Authors and Affiliations
Corresponding author
Additional information
Weiping Sun and Yan Bi contributed equally to the paper.
Rights and permissions
About this article
Cite this article
Sun, W., Bi, Y., Liang, H. et al. Inhibition of obesity-induced hepatic ER stress by early insulin therapy in obese diabetic rats. Endocr 39, 235–241 (2011). https://doi.org/10.1007/s12020-010-9429-y
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s12020-010-9429-y