
Abstract
The myogenic stem cell (satellite cell) is almost solely responsible for the remarkable regeneration of adult skeletal muscle fibers after injury. The availability and the functionality of satellite cells are the determinants of efficient muscle regeneration. During aging, the efficiency of muscle regeneration declines, suggesting that the functionality of satellite cells and their progeny may be altered. Satellite cells do not sit in isolation but rather are surrounded by, and influenced by, many extrinsic factors within the muscle tissue that can alter their functionality. These factors likely change during aging and impart both reversible and irreversible changes to the satellite cells and on their proliferating progeny. In this review, we discuss the possible mechanisms of impaired muscle regeneration with respect to the biology of satellite cells. Future studies that enhance our understanding of the interactions between stem cells and the environment in which they reside will offer promise for therapeutic applications in age-related diseases.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
One of the characteristics of organismal aging is a decline in tissue regenerative capacity leading to reduced physiological function. One reason for this impaired regenerative potential is an age-related decline in the functionality of the resident stem cells that are essential for tissue homeostasis and repair. In skeletal muscle, many aspects of the regenerative process are impaired during aging [1–6]. The role of the muscle stem cell (“satellite cell”) and its contribution to the aging phenotype of muscle tissue has recently been the subject of numerous studies. The focus of this review is the age-related changes in satellite cell function, specifically addressing evidence for cell-intrinsic changes including those that appear to be reversible, and aspects in the local and systemic satellite cell environment that change with age and impact upon satellite cell function.
Muscle Stem Cells
Anatomical Characterization
In response even to extensive injury, adult skeletal muscle can completely regenerate within several weeks in adult mammals [7–9]. This remarkable regeneration is attributed to the function and capabilities of the satellite cells, located closely apposed to the myofiber membrane, beneath its basal lamina [10]. Interstitial cells that express myogenic markers and have myogenic potential have been described [11, 12], and the relationship between these cells those in the satellite cell position remains to be determined. Other cell types, either present in the circulation or otherwise associated with the vasculature, may have some myogenic potential [13–16]. Despite the presence of these other minor populations of cells within the tissue that have myogenic potential under experimental conditions, the satellite cell population appears to be responsible for the regenerative capacity of skeletal muscle [17].
Molecular Markers
Although initially identified by purely anatomic criteria, satellite cells have increasingly been characterized by molecular markers. Among the proteins and immunologically defined markers that have been found to be present on quiescent satellite cells are c-met, M-Cadherin, CD34, Pax7, Syndecan-3, Syndecan-4, and SM-2.6 [18–21]. It has become clear that satellite cells (identified anatomically) represent a heterogeneous population of cells [18, 22, 23] and that not all satellite cells express the same combination of markers. Furthermore, many of the markers are expressed in non-myogenic cells and in myogenic cells outside the satellite cell niche. Therefore, combinations of anatomic and biochemical characterizations provide a more complete and complex description of satellite cells and related populations than either alone. A population of myogenic progenitors has also been identified in skeletal muscle on the basis of flow cytometric properties, the so-called “side population” of cells [14, 24–26]. The relationship between these cells and satellite cells also remains undefined.
Functional Characterization: Quiescence, Activation, Proliferation, Differentiation and Self-Renewal
Although their function is critical to the success of regeneration, satellite cells represent a very small percentage of the nuclear content of the tissue. For example, current estimates suggest there are between 5 and 20 satellite cells per muscle fiber containing approximately 300–500 myonuclei in distal hindlimb muscles of the mouse [18, 27–29]. In uninjured adult muscle, satellite cells are mitotically quiescent [30]. Physiological and pathological stimuli to the muscle drive the normally quiescent satellite cell to activate, enter the cell cycle and begin to express the myogenic regulatory factors (MRFs) MyoD and Myf5. Following a phase of proliferative expansion and myogenic lineage progression, cells exit the cell cycle and terminally differentiate. MyoD and Myf5 levels decline and other MRFs, such as Myogenin are expressed at the onset of differentiation (reviewed in [9]). The cells then fuse and form nascent myotubes that ultimately mature into myofibers. When muscle fibers with their associated satellite cells are placed in culture, the satellite cells activate, proliferate, and give rise to myoblasts and ultimately multinucleated myotubes [31, 32].
In studies of satellite cell activation in vitro a small fraction of satellite cells do not differentiate but rather, exit the cell cycle and return to quiescence, as defined by the loss of MyoD expression but persistence of Pax7 expression [27]. These cells can subsequently reactivate and differentiate. Therefore based on marker expression, a fraction of satellite cells appeared to undergo self-renewal in vitro. Serial transplants of single muscle fibers with their complement of satellite cells demonstrated that the transplanted cells could give rise to both differentiated cells and new satellite cells [33]. Therefore two criteria of a stem cell are fulfilled by the satellite cell: the capacity to give rise to more differentiated tissue and self-renewal.
Aging and Muscle Stem Cells
The declining regenerative potential of skeletal muscle with age raises the following question: To what extent can the impaired regenerative potential of aged muscle be attributed to age-related changes in the satellite cell population? Although there is considerable evidence that satellite cell function declines with age, and that there may be both cell-intrinsic and cell-extrinsic changes that underlie that decline, the basic issue as to whether the number of muscle stem cells changes with age has been studied using different approaches and yielded divergent results (Table 1). Even more importantly, whether any observed change in number could in fact account for age-related changes in regenerative potential has been, in general, poorly addressed. Therefore, before any age-related changes in muscle stem cell functionality are discussed, the issue of age-related changes in stem cell number, and its consequences, will be considered.
Aging and Muscle Stem Cell Number
Determining Progenitor Numbers
Among the many studies that have sought to quantify muscle stem cell number with age and yielded quite different results, the divergent methodologies have precluded direct comparisons (Table 1). From these reports, it has been variably concluded that stem cell number decreases, changes very little if at all, and increases. However, few studies have used the same methods, and the critical differences include species and strain studied, muscle(s) analyzed, ages compared, and methodologies of quantitation. Clearly, it would be unwarranted to extrapolate from one species to another, just as it would be to extrapolate from one strain to another given the well-recognized, profound biological differences between different inbred strains. Likewise, it is impossible to compare one muscle group to another since it is well known that the number of satellite cells associated with slow fibers is, on average, quite different from the number of satellite cells associated with fast fibers [27]. Even if these two variables were controlled, the ages examined and methods of quantitation would still profoundly influence the results and interpretations of estimates of changes in muscle stem cell number.
Ages analyzed
There is no universal agreement about the age at which an individual of any species is considered “old”, but there are several important considerations that are often neglected and that confound studies of aging. First, the comparison age must be a mature adult, not a growing young adult. It is well known that there is a sharp decline in satellite cell number during postnatal growth [34] and it would be a mistake to include this decline in the estimates of the effects of aging. The starting point needs to be at an age when the number of satellite cells in the muscle changes negligibly over the next several months. Second, and equally important, is the mistake of lumping “old” with “very old” animals. In the case of mice, very old animals (e.g. around 30 months of age) virtually all develop a variety of systemic diseases and pathologies that could easily influence any anatomical or biochemical measure. Therefore, it is essential that the age range used to evaluate normal aging begin, at the earliest, in early adulthood (∼3–4 months) and extend only to the point (∼24 months) after which age-related pathologies are likely to confound studies of normal aging.
Methods of quantifying muscle stem cell number
-
i.
Anatomical characterization
Satellite cells were initially identified by electron microscopy by virtue of their sublaminar position [10]. More recently, anatomical methods have been used to quantify the number of mononucleated cells associated with isolated single fibers [28, 29, 35]. From the tibialis anterior and extensor digitorum longus muscles in the mouse, such analyses have yielded estimates of the number of satellite cells per single muscle fiber. Combined with biochemical markers analyzed by immunofluorescence, these estimates are rigorous, but are likely to be underestimates of the number of myogenic progenitors for two reasons. First, there is increasing evidence that fiber-associated satellite cells are likely to be in a dynamic relationship with cells in the interstitium [11, 12], and that, with age, changes in extracellular matrix may result in fewer myogenic progenitors tightly associated with the muscle fiber. Thus, the procedures used to isolate single fibers to study satellite cell function may underestimate the number of myogenic progenitors present, even though the functionality of myogenic progenitors in the interstitium or loosely associated with fibers may differ from the functionality of cells tightly associated with fiber. Second, the identification of progenitors based on this anatomical location but also by biochemical markers has the added caveat of the uncertainty of age-related changes in gene expression (see next section).
-
ii.
Biochemical characterization
Because of the historical precedent of identifying satellite cells based on anatomical location, the use of molecular markers was initially combined with anatomical analysis. However, molecular markers are increasingly being used to identify and quantitate myogenic progenitors. This, of course, comes with inherent uncertainties. No single marker (including c-met, M-cadherin, CD34, Pax7, Pax3, Myf5, Syndecan-4) is 100% sensitive. Thus, there is no gold standard for comparison and the risk of circular reasoning is high. The absence of a marker is not proof that a cell is not a myogenic progenitor. Furthermore, none of the markers are 100% specific for satellite cells as they are expressed in cells of other lineages (including c-met, CD34, Syndecan-4) or are expressed in more differentiated satellite cell progeny (including M-Cadherin, Pax3, Pax7, and Myf5). Finally, and most important with regard to the issue of quantitating changes in myogenic progenitor number with age, there is no reason to assume that there are not changes in expression patterns for any of these markers with age, even within defined myogenic progenitor compartments such as satellite cells [29]. Therefore, reports of reduced numbers of cells expressing any of these “satellite cell markers” in muscles of aged mice do not necessarily prove that there are reduced numbers of satellite cells.
-
iii.
Functional characterization
Estimating myogenic progenitor number based on functional assays is probably the most problematic of all when the methodologies involve growth in vitro, primarily because of the increasingly well-recognized changes in the functionality of satellite cells with age. Most notably, estimating progenitor cell number based on colony size, number following clonal plating of cells from young and aged animals, or the number of myoblasts derived from single myofibers of different ages all assume that all cells will behave equally once placed in culture [36, 37]. Any age-related cell-intrinsic change leading to lower rates of activation or proliferation, even such changes that may be reversed after more prolonged exposure to mitogenic stimuli in vitro, would result in an underestimate of the number of satellite cells plated. Therefore, such functional read-outs cannot be used to estimate satellite cell number with any accuracy.
Summary and Conclusions about Studies of Age-Related Changes of Satellite Cell Number
From the studies listed in Table 1, it is clear that there are no general conclusions about the changes in satellite cell number with age. Even in the same species and muscle (human vastus lateralis) using the same technique (electron microscopic analysis), the results range from no significant change [38] to a 30–40% decline [39]. Among the studies investigating age-related changes in satellite cell number from mouse tibialis anterior and/or extensor digitorum longus muscles, including only those results that compare adult to aged mice (not very young or very old mice), the results range from minimal change [29, 35] to up to a ∼40% decline [28, 40], noting that the studies differ in terms of methods of analysis. Importantly, it is clear that the expression of a widely used marker of satellite cells, Pax7, declines with age in fiber-associated cells [29], highlighting the potential inaccuracy of relying on the assumption that expression patterns of satellite cell markers do not change with age. Interestingly, in all studies, there is no significant decline and maybe even an increase in satellite cell number in the soleus muscle in rodents [35, 40, 41], consistent with the finding that in total hindlimb muscle groups, there is, at most, a minimal decline in satellite cell number with age [5].
Significance of Changes in Satellite Cell Number with Age
It is unlikely that differences in estimates of the changes in satellite cell number with age will be easily reconciled because of these critical methodological differences. However, even if all of these issues could be controlled and systematic analyses, taking into accounts the caveats of each method, could be performed on a range of muscles across strains and species, the fundamental question that remains to be answered is: Does it matter? The important issue is being able to distinguish cause-and-effect from a mere association, a distinction that would seem obvious but is often still overlooked. The underlying assumption in many studies in which changes in satellite cell number with age are measured is that the results of those studies will be causally related to the changes in muscle regenerative potential with age. However, direct experimental evidence suggests any decline in satellite cell number with age is of limited, if any, functional significance. Given the right stimuli and environment, the complement of satellite cells in aged muscle are perfectly capable of engaging in rapid and effective muscle regeneration, yielding results that are essentially indistinguishable or negligibly different from those in young muscle. This was first demonstrated by the studies of Carlson and colleagues using a heterochronic transplantation approach [2]. The conclusion of those studies was that the progenitors present in aged muscle were as efficient in terms of regenerating muscle as were the progenitors present in young muscle as long as they were transplanted into a young host. More recently it was shown, using a heterochronic parabiotic approach, that aged muscle was as efficient in regenerating following injury as was young muscle if that aged muscle had been exposed to the systemic environment which normally surrounds young muscle [42]. Thus, both of these lines of study argue strongly that any normal changes in the number of progenitors present in aged muscle are not significant determinants of age-related changes in regenerative capacity. The experiment that has not been done is one in which young muscle is depleted of its progenitors to varying degrees to test at what point does progenitor number become a limiting factor in terms of promoting normal muscle regeneration. If the largest estimates of age-related declines in progenitor number are accurate, and if the heterochronic experiments described above are correct, then it would seem that one could deplete young muscle of more that half of its resident progenitors and the muscle would still regenerate perfectly well. A decline of progenitor number may be associated with aging, but it appears not to be causally related to age-related impairment of regenerative capacity.
Aging and the Decline in Satellite Cell Functionality
Clearly, the age-related decline in muscle regenerative capacity is due primarily to an age-related decline in satellite cell function, not to a depletion of stem cell numbers. As an example of such a functional decline is the marked reduction in satellite cell activation and proliferation during regeneration of aged muscle compared with young muscle, resulting in the generation of fewer progeny [36, 37, 43]. It is possible that such changes in satellite cell function are cell-intrinsic changes resulting from acquired genetic mutations or to epigenetic changes that occur with age. Any age-related impairment of functionality assessed in vivo could also result from cell-extrinsic influences arising from the local microenvironment of the stem cell niche or from the systemic environment, both of which change with age [2, 6, 42]. As such, many apparently cell-autonomous changes with age may be reversible changes when cells are maintained in the appropriate environment either in vivo or in vitro. In order to differentiate among the various influences of aging on stem cell functionality, it is useful to distinguish cell-intrinsic changes (whether reversible or irreversible) from cell-extrinsic changes in the local and systemic environment.
Cell-Intrinsic Changes with Age
To study intrinsic aspects of cellular aging, differences in the external environment need to be eliminated. Therefore cells need to be transferred to identical environments, either in vitro or in vivo. If cell-intrinsic changes are found to occur with age, it is important to consider whether this measured difference is stable and irreversible (such as cumulative genetic mutations) or potentially reversible (such as epigenetic regulation of gene expression).
Molecular and Biochemical Properties
One irreversible cell-intrinsic change that occurs with age in virtually all dividing somatic cells is telomere shortening [44]. This process has been associated with phenotypic changes in aging tissues because the shortening of telomere length below a critical level can result in cell death or senescence instead of proliferation [45]. Satellite cells isolated from muscles of human patients do not show a decrease in minimum or mean telomere length during aging [46], but a correlation between proliferation potential and telomere length has been observed [47, 48]. Comparable studies have not been done in rodents, but rodents have much longer telomeres than do humans so their satellite cells may not display critical telomere shortening. Furthermore, whether cell senescence occurs in vivo in normal mammalian skeletal muscle aging has yet to be determined. Since age-related impairment of skeletal muscle regeneration extends across species, it is currently not clear to what extent telomere shortening is an important cell-intrinsic change contributing to the aging phenotype.
Gene expression studies from isolated satellite cells or their progeny have demonstrated that the transcriptional profile changes with age [49]. It is not known which of these alterations are due to irreversible genetic change or reversible epigenetic effects. During satellite cell activation and proliferation of myogenic progeny, the expression of muscle specific genes is regulated by epigenetic mechanisms [50]. Epigenetic regulation of gene expression is a possible mechanism to explain stem cell aging as it influences both self renewal and senescence [51].
In Vitro Functional Studies
Beyond the elucidation of any biochemical changes that may differ between muscle stem cells from adult and aged animals, the more complex physiological characterization using in vitro functional assays is likely to reveal changes that reflect changes in cell behavior in vivo. Although overlapping, different sequential phases that satellite cell progeny progress through from the quiescent stem cell to the differentiated, multinucleated myotube can be analyzed for age-related changes, as detailed in the following sections.
-
i.
Activation and Proliferation
Satellite cells isolated from young and aged muscle have been tested for their ability to activate and proliferate in vitro. Myogenic cells from aged muscle show a delayed response to activating stimuli [36, 52], which results in fewer progeny compared to young satellite cells [5, 35, 37]. In contrast, progenitors from old muscle that have been maintained in culture after many passages proliferate almost as well as progenitors from young muscle [35, 52]. Whether the difference observed during initial growth in vitro is a cell-intrinsic change in the aged satellite cells that is reversible, or whether a similar subpopulation of cells are selected for over time from both the young and old satellite cells, is not clear and would require more detailed clonal analysis of the fates of individual cells.
-
ii.
Myogenic Lineage Progression
After initial activation, satellite cell progeny progress in their lineage from early progenitors to fusion-competent myoblasts. Notch signaling regulates the proliferative expansion of transit amplifying cells in the transition from the quiescent satellite cell to fusion-competent myoblasts [53]. In aged muscle, an impairment of Notch signaling, related to a failure of expression of the Notch ligand Delta in response to stimuli that induce Delta expression in young muscle, results in reduced proliferative expansion of progenitors and ineffective regeneration [5]. Forced activation of Notch signaling in injured muscle of aged mice restores regenerative potential to that tissue [5].
We have found that Wnt signaling plays a critical role in the lineage progression of satellite cell progeny after proliferative expansion as Notch signaling declines (Brack et al, unpublished observations). Wnt signaling increases during the proliferative expansion of the transit amplifying cells, decreases their proliferative capacity, and promotes their progression to become fusion-competent myoblasts expressing high levels of desmin. It is not known whether the transition from transit amplifying cell to committed myoblast is affected by aging, but it is clear that effective regeneration depends on a tightly controlled temporal switch between Notch and Wnt signaling to control myogenic lineage progression. In aged muscle, an increase in Wnt signaling in early activated satellite cells shifts this balance and leads to less proliferative expansion and premature differentiation, resulting in less effective regeneration [6]. This argues that altered key signaling pathways may impair lineage progression during aging. However, some satellite cells from old muscle do progress down the myogenic lineage in vitro as there are numerous reports of myoblasts expressing later lineage markers such as desmin and myogenin [35, 37, 54]. Determining whether there is a specific defect in myogenic lineage progression of aged satellite cell progeny will require an analysis of individual cells to determine whether, once they have begun proliferating, they proceed along the myogenic lineage as readily as cells from young tissue.
-
iii.
Alternate Lineage Pathways of the Satellite Cell
Satellite cells have the ability to express markers of alternative mesenchymal lineage pathways, including including osteogenic, chondrogenic, and adipogenic lineages [55–57]. This raises the possibility that the satellite cell has multipotentiality, although that potentially appears to be restricted to the mesenchymal lineage.
Myogenic cells from mice lacking Wnt10b have an enhanced adipogenic profile both in vitro and in vivo and injury to the muscle results in an accumulation of fat deposits [58]. It is interesting that Wnt signaling, while being important in the regulation of myogenic lineage progression, may also be important for lineage determination [58, 59]. We have found that aged myogenic progenitors can diverge from a myogenic lineage and adopt a fibroblast fate [6]. This change of fate, or transdifferentiation, is due to increased canonical Wnt signaling present in the aged progenitors. Furthermore, inhibition of Wnt signaling prevented much of the elevated fibrosis that occurs in the aged muscle following injury [6]. Given the pleiotropic effects of Wnts and the number of different Wnts and Wnt receptors, it is possible that specific combinations have either positive or negative affects on maintaining the myogenic lineage. It may be that the specific timing is critical as to whether a single Wnt may have positive or negative effects on cell fate in aged muscle progenitors and regeneration in vivo.
In addition to aged satellite cell progeny adopting an alternate lineage, an increased tendency of aged satellite cells to undergo senescence or apoptosis may also occur [60]. This would also hinder effective muscle repair and be consistent with a cell intrinsic property of the stem cell being responsible for the impaired regenerative response of aged muscle.
-
iv.
Myogenic Differentiation
Aged muscle does give rise to multinucleated muscle fibers during regeneration, although the regenerating fibers are smaller and therefore differentiation may be impaired. For the limited number of satellite cells that do activate, proliferate, and progress down the myogenic lineage pathway in aged muscle, their ability to differentiate into multinucleated myotubes has been compared to that of young cells. The ability of aged myogenic progenitors to fuse to each other and become multinucleated myotubes does not appear impaired in aged satellite cells [35]. However, from single fiber or myoblast cultures, fewer myogenic cells isolated from old muscle expressed myosin heavy chain after 2 days in differentiating conditions, consistent with impaired differentiation [54]. Even after longer periods of time to allow differentiation to proceed, fewer cells from aged animals expressed the myogenic differentiation transgenic reporter 3F-NLacZ-2E [29]. This argues against a subtle temporal delay in myogenic differentiation of aged myogenic progenitors to explain their phenotype. Conversely, no defects in differentiation were observed if progenitor cells underwent multiple passages prior to assessing myogenic differentiation [5]. These data suggest that there are cell-intrinsic changes with age that render older satellite cells less effective in undergoing myogenic differentiation, but that these changes are in fact reversible when the cells are placed in an environment that promotes the processes of growth and differentiation necessary for effective regeneration.
Cell-Extrinsic Changes with Age
Clearly there are cell-intrinsic changes during aging are indeed reversible, suggesting that satellite cell functionality is influenced by the environment in which they reside, either in their dormant state or their active state in response to injury stimuli. The systemic environment of the aged animal does influence the function of cells in general [61, 62]. Direct evidence for an effect of the systemic environment comes from two different kinds of heterochronic studies. The first is a heterochronic transplantation experiment in rats in which whole muscles were isolated from young or old donors and then transplanted into young or old hosts [2]. These studies demonstrated that engraftment success was improved if old muscle was transplanted into a young host compared to an old host, whereas the regeneration of young muscle transplanted into an old host was impaired relative to that in a young host. These results strongly supported the hypothesis that factors within the host tissue were more important than the source of the cells in determining the effectiveness of the regenerative response. Whether factors in the muscle that influenced regeneration were produced locally or were systemic and localized to the muscle were not addressed. More recently, single muscle fibers from old mice were transplanted into young irradiated mice; the engraftment success of the old muscle fibers was not distinguishable from young fibers injected into irradiated young mice [29]. This provides further evidence that the host environment plays a major role in satellite cell functionality and muscle regeneration.
Parabiotic studies have also demonstrated that the host environment is critical to the success of regeneration by aged satellite cells [6, 42]. In these studies, parabiotic pairings of young and old mice were followed by muscle injury to both mice, and muscle satellite cell activation and regenerative response was analyzed. Consistent with transplant studies, it was found that the efficacy of satellite cell activation and muscle regeneration was determined by the systemic environment. In heterochronic pairs (old mice paired with young mice), the effectiveness of satellite cell activation and muscle regeneration in old parabionts was markedly improved by exposure to the youthful systemic milieu [42]. Conversely, the young parabionts demonstrated a decline in functionality by being paired to an old partner. More recently it was demonstrated that the systemic environment can mediate the lineage of satellite cells. Using the parabiosis technique, satellite cells from a young mouse paired with aged mouse were less able to maintain their myogenic fate; the satellite cells acquired a fibrogenic fate, whereas satellite cells from aged mice that had been paired with young mice could better maintain their myogenic fate [6]. These results suggest that there is some combination of positive influences of the young systemic environment and negative effects of the old systemic environment on myogenic stem cell functionality.
Together, these results demonstrate that during aging the cell-extrinsic environment (local and systemic) mediates stem cell functionality. The environment is complex, ranging from extrinsic factors that are in contact with the stem cell, the stem cell niche, autocrine and paracrine factors both at rest and after injury, and finally circulating factors. Each of these and how they change with age, and ultimately, influence satellite cell functionality will be considered separately.
Satellite Cell Niche
The stem cell niche is the microenvironment that sustains the characteristic of the stem cell, the ability to maintain quiescence until required to activate, differentiate and self-renew. The study of the niche in regards to the muscle stem cell biology is still in its infancy. Therefore it is unknown what are the components that comprise the satellite cell niche. The satellite cell is in close association with the myofiber without any intervening basal lamina, and an overlying basal lamina that separates the satellite cell from other fibers and interstitial cells. Therefore the muscle fiber and the basal lamina are likely components of the niche. Other constituents in the niche could also include fibroblasts, endothelial cells, infiltrating inflammatory cells and local vascular components, particularly adjacent capillaries [63]. There is a reduced capillarization in aged compared to young muscle [64]. This would not only reduce blood flow to the muscle, but it could also reduce the availability of systemic (circulating) factors necessary for satellite cell function (discussed below).
There is an increase in connective tissue between the fibers and surrounding the fibers in aged compared to young muscle [65]. The basal lamina encapsulating the fiber is also thicker [41, 65] resulting in more collagen surrounding each fiber in aged mice [66]. This could influence both the physical connection with the satellite cell as well as paracrine factors released from the fiber and the basal lamina (discussed below). Clearly, age-related alterations in the composition and deposition of extracellular matrix components could influence every aspect of satellite cell function. Recently, it was reported that mesenchymal stem cells can alter their fate depending on stiffness of the matrix the cells reside [67]. Therefore with more collagen surrounding the aged muscle fiber and a stiffer matrix, satellite cells may be less able to maintain their myogenic fate in the aged niche.
Autocrine/Paracrine Factors
There are growth factors that have been isolated from crushed muscle extracts such as hepatocyte growth factor (HGF) and fibroblast growth factor (FGF), which can enhance the activation and proliferation of aged satellite cells in vitro [35, 68–71]. There are many cellular constituents in muscle and it is not completely resolved which factors are secreted from which cell type. It is known that muscle-resident fibroblasts can secrete soluble factors that act on myoblasts [69, 72] and that TGFβ secreted by invading macrophages can affect myogenic cell function [73]. Any changes of these factors with age would be important in terms of understanding declining satellite cell functionality [74]. Indeed, muscle extracts from old mice were less mitogenic to both young and old myogenic progenitors than were extracts from young muscle, consistent with an age-dependent change in paracrine regulation [75]. Whether differences in the aged muscle extract were due to loss of factors that promote satellite cell function or an increase in inhibitory factors was not determined. It is plausible that age-related loss of satellite cell functionality is due to an increase in inhibitory factors [74]. In cell culture, myoblasts from old muscle had elevated TGFβ signaling [76]. TGFβ impairs proliferation of myoblasts [73] and can increase fibrosis [77] making it a candidate for an inhibitory paracrine factor that changes during aging.
Systemic Factors of the Host Environment
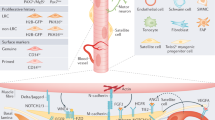
The systemic environment is another potential source of growth-promoting and growth-inhibitory factors that could alter satellite cell function during aging (Fig. 1). Heterochronic parabiosis experiments demonstrated that the systemic environment plays a major role in the functionality of the satellite cell [6, 42]. From in vitro experiments, sera from young or old mice have effects on satellite cell function that correspond to the effects of age and heterochronic parabiotic pairings on Notch activation and satellite cell activation was well as muscle regeneration in vivo [42].

A model of systemic factors influencing the satellite cell environment in uninjured and injured muscle. In uninjured muscle, quiescent satellite cells reside underneath the basal lamina, surrounded by connective tissue. In aged muscle, there is an increase in connective tissue surrounding the myofibers compared to young muscle. The myofibers and their complement of satellite cells are bathed in stimulatory and inhibitory systemic factors present in the circulation. In aged muscle there is an increase in inhibitory components. In injured muscle, myofibers degenerate and the satellite cells are activated. The satellite cells are exposed to stimulatory and inhibitory systemic factors via the circulation. In the young environment, stimulatory factors facilitate proliferation of satellite cells and their progeny and the maintenance of their myogenic lineage, providing a source of myoblasts to fuse to form nascent myofibers. In the aged environment, an increase in inhibitory components inhibits the active of satellite cells and the proliferation of their progeny. In addition, the aged systemic milieu is less effective in maintaining the myogenic lineage of satellite cell progeny, leading to a conversion of some of these cells to a fibrogenic fate. As a result, there are not only fewer myogenic progenitors available to form nascent myofibers, but the increased production of fibroblasts in the regenerating tissue leads to an increase in connective tissue deposition and an increase in fibrosis
Serum has a vast array of constituents and the effects of either young or old serum on satellite cell function is likely to be multifactorial. Most protein growth factors that are known to regulate myogenic progenitor proliferation in vitro are present in the circulation such as IGF-1 and TGF-β. IGF-1 can stimulate myoblast cell proliferation in cell culture [78]. In aged humans, reduced serum levels of IGF-1 have been observed [79], a change that could account for differences in satellite cell activation and proliferation with age.
Following activation, satellite cells from aged muscle have increased Wnt signaling compared to satellite cells from young muscle [6]. We have observed that this is mediated by the systemic environment using both in vivo and in vitro assays [6]. Intriguingly, aged serum has increased levels of proteins that bind to the canonical Wnt receptor, Frizzled [6], raising the possibility that some form of Wnt or Wnt-like molecule is present in the circulation and may modulate satellite cell function. Recent evidence also suggests that Klotho may act as a circulating Wnt-inhibitory protein and that the premature aging phenotypes seen in Klotho-deficient mice may be mediated by heightened Wnt signaling in the stem cell compartments of tissues throughout the body [80].
Steroid hormones are another category of circulating factors that need to be considered as systemic modulators of stem cell function. Androgens have been found to play a role in proliferation and differentiation of satellite cells [81]. Decreased levels of androgens have been reported in aged men [82], raising the possibility that this could account in part for declining satellite cell functionality.
Conclusions
Clearly, muscle stem cell function is profoundly altered during the aging process. Understanding the underlying mechanisms will reflect the basic biology of aging that is likely to affect all stems cells analogously. There may also be unique aspects of satellite cell aging related to the specific combination of signal transduction pathways and transcriptional networks that are not shared with stem cells in other tissues. However, understanding the aging of stem cell functionality by distinguishing irreversible from reversible cell-intrinsic changes is essential for every cell type. Furthermore, elucidating the relative contributions of cell-intrinsic changes from environmental influences at the time of stem cell activation, proliferation, and differentiation is necessary to understand the key factors that render stem cells in aged tissues less effective in tissue maintenance and repair.
References
Sadeh, M., Czyewski, K., & Stern, L. Z. (1985). Chronic myopathy induced by repeated bupivacaine injections. Journal of the Neurological Sciences, 67, 229–238.
Carlson, B. M., & Faulkner, J. A. (1989). Muscle transplantation between young and old rats: age of host determines recovery. American Journal Physiology, 256, C1262–C1266.
McGeachie, J. K., & Grounds, M. D. (1995). Retarded myogenic cell replication in regenerating skeletal muscles of old mice: An autoradiographic study in young and old BALBc and SJL/J mice. Cell & Tissue Research, 280, 277–282.
Carlson, B. M., Dedkov, E. I., Borisov, A. B., & Faulkner, J. A. (2001). Skeletal muscle regeneration in very old rats. J Gerontol Ser A Biol Sci Med Sci, 56, B224–B233.
Conboy, I. M., Conboy, M. J., Smythe, G. M., & Rando, T. A. (2003). Notch-mediated restoration of regenerative potential to aged muscle. Science, 302, 1575–1577.
Brack, A. S., Conboy, M. J., Roy, S., Lee, M., Kuo, C. J., Keller, C., et al. (2007). Increased Wnt signaling during aging alters muscle stem cell fate and increases fibrosis. Science, 317, 807–810.
Hawke, T. J., & Garry, D. J. (2001). Myogenic satellite cells: Physiology to molecular biology. Journal Applied Physiology, 91, 534–551.
Chargé, S. B., & Rudnicki, M. A. (2004). Cellular and molecular regulation of muscle regeneration. Physiological Reviews, 84, 209–238.
Zammit, P. S., Partridge, T. A., & Yablonka-Reuveni, Z. (2006). The skeletal muscle satellite cell: The stem cell that came in from the cold. Journal of Histochemistry and Cytochemistry, 54, 1177–1191.
Mauro, A. (1961). Satellite cells of skeletal muscle fibers. Journal of Biophysical and Biochemical Cytology, 9, 493–495.
Tamaki, T., Akatsuka, A., Ando, K., Nakamura, Y., Matsuzawa, H., Hotta, T., et al. (2002a). Identification of myogenic-endothelial progenitor cells in the interstitial spaces of skeletal muscle. Journal Cell Biology, 157, 571–577.
Tamaki, T., Akatsuka, A., Yoshimura, S., Roy, R. R., & Edgerton, V. R. (2002b). New fiber formation in the interstitial spaces of rat skeletal muscle during postnatal growth. Journal of Histochemistry and Cytochemistry, 50, 1097–1111.
De Angelis, L., Berghella, L., Coletta, M., Lattanzi, L., Zanchi, M., Cusella-De Angelis, M. G., et al. (1999). Skeletal myogenic progenitors originating from embryonic dorsal aorta coexpress endothelial and myogenic markers and contribute to postnatal muscle growth and regeneration. Journal of Cell Biology, 147, 869–878.
McKinney-Freeman, S. L., Jackson, K. A., Camargo, F. D., Ferrari, G., Mavilio, F., & Goodell, M. A. (2002). Muscle-derived hematopoietic stem cells are hematopoietic in origin. Proceedings of the National Academy of Sciences of the United States of America, 99, 1341–1346.
Polesskaya, A., Seale, P., & Rudnicki, M. A. (2003). Wnt signaling induces the myogenic specification of resident CD45+ adult stem cells during muscle regeneration. Cell, 113, 841–852.
Galvez, B. G., Sampaolesi, M., Brunelli, S., Covarello, D., Gavina, M., Rossi, B., et al. (2006). Complete repair of dystrophic skeletal muscle by mesoangioblasts with enhanced migration ability. Journal of Cell Biology, 174, 231–243.
Zammit, P., & Beauchamp, J. (2001). The skeletal muscle satellite cell: Stem cell or son of stem cell? Differentiation, 68, 193–204.
Beauchamp, J. R., Heslop, L., Yu, D. S., Tajbakhsh, S., Kelly, R. G., Wernig, A., et al. (2000). Expression of CD34 and Myf5 defines the majority of quiescent adult skeletal muscle satellite cells. Journal Cell Biology, 151, 1221–1234.
Cornelison, D. D., Filla, M. S., Stanley, H. M., Rapraeger, A. C., & Olwin, B. B. (2001). Syndecan-3 and syndecan-4 specifically mark skeletal muscle satellite cells and are implicated in satellite cell maintenance and muscle regeneration. Developments in Biologicals, 239, 79–94.
Seale, P., Ishibashi, J., Scime, A., & Rudnicki, M. A. (2004). Pax7 Is necessary and sufficient for the myogenic specification of CD45(+):Sca1(+) stem cells from injured muscle. PLoS Biol, 2, E130.
Fukada, S. I., Uezumi, A., Ikemoto, M., Masuda, S., Segawa, M., Tanimura, N., et al. (2007). Molecular signature of quiescent satellite cells in adult skeletal muscle. Stem Cells (in press).
Kuang, S., Charge, S. B., Seale, P., Huh, M., & Rudnicki, M. A. (2006). Distinct roles for Pax7 and Pax3 in adult regenerative myogenesis. Journal of Cell Biology, 172, 103–113.
Kuang, S., Kuroda, K., Le, G. F., & Rudnicki, M. A. (2007). Asymmetric self-renewal and commitment of satellite stem cells in muscle. Cell, 129, 999–1010.
Jackson, K. A., Mi, T., & Goodell, M. A. (1999). Hematopoietic potential of stem cells isolated from murine skeletal muscle. Proceedings of the National Academy of Sciences of the United States of America, 96, 14482–14486.
Asakura, A., Seale, P., Girgis-Gabardo, A., & Rudnicki, M. A. (2002). Myogenic specification of side population cells in skeletal muscle. Journal of Cell Biology, 159, 123–134.
Uezumi, A., Ojima, K., Fukada, S., Ikemoto, M., Masuda, S., Miyagoe-Suzuki, Y., et al. (2006). Functional heterogeneity of side population cells in skeletal muscle. Biochemical and biophysical research communications, 341, 864–873.
Zammit, P. S., Heslop, L., Hudon, V., Rosenblatt, J. D., Tajbakhsh, S., Buckingham, M. E., et al. (2002). Kinetics of myoblast proliferation show that resident satellite cells are competent to fully regenerate skeletal muscle fibers. Experimental Cell Research, 281, 39–49.
Brack, A. S., Bildsoe, H., & Hughes, S. M. (2005). Evidence that satellite cell decrement contributes to preferential decline in nuclear number from large fibres during murine age-related muscle atrophy. Journal Cell Science, 118, 4813–4821.
Collins, C. A., Zammit, P. S., Ruiz, A. P., Morgan, J. E., & Partridge, T. A. (2007). A population of myogenic stem cells that survives skeletal muscle aging. Stem Cells, 25, 885–894.
Schultz, E., Gibson, M. C., & Champion, T. (1978). Satellite cells are mitotically quiescent in mature mouse muscle: An EM and radioautographic study. Journal Experimental Zoology, 206, 451–456.
Bischoff, R. (1975). Regeneration of single skeletal muscle fibers in vitro. Anatomical Record, 182, 215–235.
Rosenblatt, J. D., Lunt, A. I., Parry, D. J., & Partridge, T. A. (1995). Culturing satellite cells from living single muscle fiber explants. In Vitro Cell Dev Biol, 31, 773–779.
Collins, C. A., Olsen, I., Zammit, P. S., Heslop, L., Petrie, A., Partridge, T. A., et al. (2005). Stem cell function, self-renewal, and behavioral heterogeneity of cells from the adult muscle satellite cell niche. Cell, 122, 289–301.
Cardasis, C. A., & Cooper, G. W. (1975). An analysis of nuclear numbers in individual muscle fibers during differentiation and growth: A satellite cell-muscle fiber growth unit. Journal Experimental Zoology, 191, 347–358.
Shefer, G., Van de Mark, D. P., Richardson, J. B., & Yablonka-Reuveni, Z. (2006). Satellite-cell pool size does matter: Defining the myogenic potency of aging skeletal muscle. Developments in Biologicals, 294, 50–66.
Schultz, E., & Lipton, B. H. (1982). Skeletal muscle satellite cells: Changes in proliferation potential as a function of age. Mechanism of Ageing and Development, 20, 377–383.
Bockhold, K. J., Rosenblatt, J. D., & Partridge, T. A. (1998). Aging normal and dystrophic mouse muscle: Analysis of myogenicity in cultures of living single fibers. Muscle Nerve, 21, 173–183.
Roth, S. M., Martel, G. F., Ivey, F. M., Lemmer, J. T., Metter, E. J., Hurley, B. F., et al. (2000). Skeletal muscle satellite cell populations in healthy young and older men and women. Anatomical Record, 260, 351–358.
Sajko, S., Kubinova, L., Cvetko, E., Kreft, M., Wernig, A., & Erzen, I. (2004). Frequency of M-cadherin-stained satellite cells declines in human muscles during aging. Journal of Histochemistry and Cytochemistry, 52, 179–185.
Gibson, M. C., & Schultz, E. (1983). Age-related differences in absolute numbers of skeletal muscle satellite cells. Muscle Nerve, 6, 574–580.
Snow, M. H. (1977). The effects of aging on satellite cells in skeletal muscles of mice and rats. Cell & Tissue Research, 185, 399–408.
Conboy, I. M., Conboy, M. J., Wagers, A. J., Girma, E. R., Weissman, I. L., & Rando, T. A. (2005). Rejuvenation of aged progenitor cells by exposure to a young systemic environment. Nature, 433, 760–764.
Conboy, I. M., & Rando, T. A. (2005). Aging, stem cells and tissue regeneration: Lessons from muscle. Cell Cycle, 4, 407–410.
Bekaert, S., Derradji, H., & Baatout, S. (2004). Telomere biology in mammalian germ cells and during development. Developments in Biologicals, 274, 15–30.
Campisi, J. (2001). From cells to organisms: Can we learn about aging from cells in culture? Experimental Gerontology, 36, 607–618.
Decary, S., Mouly, V., Hamida, C. B., Sautet, A., Barbet, J. P., & Butler-Browne, G. S. (1997). Replicative potential and telomere length in human skeletal muscle: Implications for satellite cell-mediated gene therapy. Human Gene Therapy, 8, 1429–1438.
Decary, S., Hamida, C. B., Mouly, V., Barbet, J. P., Hentati, F., & Butler-Browne, G. S. (2000). Shorter telomeres in dystrophic muscle consistent with extensive regeneration in young children. Neuromuscular Disorders, 10, 113–120.
Mouly, V., Aamiri, A., Bigot, A., Cooper, R. N., Di, D. S., Furling, D., et al. (2005). The mitotic clock in skeletal muscle regeneration, disease and cell mediated gene therapy. Acta Physiologica Scandinavica, 184, 3–15.
Bortoli, S., Renault, V., Eveno, E., Auffray, C., Butler-Browne, G., & Pietu, G. (2003). Gene expression profiling of human satellite cells during muscular aging using cDNA arrays. Gene, 321, 145–154.
Palacios, D., & Puri, P. L. (2006). The epigenetic network regulating muscle development and regeneration. Journal of Cellular Physiology, 207, 1–11.
Chambers, S. M., Shaw, C. A., Gatza, C., Fisk, C. J., Donehower, L. A., & Goodell, M. A. (2007). Aging hematopoietic stem cells decline in function and exhibit epigenetic dysregulation. PLoS Biol, 5, e201.
Barani, A. E., Durieux, A. C., Sabido, O., & Freyssenet, D. (2003). Age-related changes in the mitotic and metabolic characteristics of muscle-derived cells. Journal of Applied Physiology, 95, 2089–2098.
Conboy, I. M., & Rando, T. A. (2002). The regulation of Notch signaling controls satellite cell activation and cell fate determination in postnatal myogenesis. Developments in Cell Biology, 3, 397–409.
Chargé, S. B., Brack, A. S., & Hughes, S. M. (2002). Aging-related satellite cell differentiation defect occurs prematurely after Ski-induced muscle hypertrophy. American Journal of Physiology Cell Physiol, 283, C1228–C1241.
Asakura, A., Komaki, M., & Rudnicki, M. (2001). Muscle satellite cells are multipotential stem cells that exhibit myogenic, osteogenic, and adipogenic differentiation. Differentiation, 68, 245–253.
Wada, M. R., Inagawa-Ogashiwa, M., Shimizu, S., Yasumoto, S., & Hashimoto, N. (2002). Generation of different fates from multipotent muscle stem cells. Development, 129, 2987–2995.
Shefer, G., Wleklinski-Lee, M., & Yablonka-Reuveni, Z. (2004). Skeletal muscle satellite cells can spontaneously enter an alternative mesenchymal pathway. Journal of Cell Science, 117, 5393–5404.
Vertino, A. M., Taylor-Jones, J. M., Longo, K. A., Bearden, E. D., Lane, T. F., McGehee, R. E., Jr., et al. (2005). Wnt10b deficiency promotes coexpression of myogenic and adipogenic programs in myoblasts. Molecular Biology of the Cell, 16, 2039–2048.
Taylor-Jones, J. M., McGehee, R. E., Rando, T. A., Lecka-Czernik, B., Lipschitz, D. A., & Peterson, C. A. (2002). Activation of an adipogenic program in adult myoblasts with age. Mechanism of Ageing and Development, 123, 649–661.
Jejurikar, S. S., Henkelman, E. A., Cederna, P. S., Marcelo, C. L., Urbanchek, M. G., & Kuzon, W. M., Jr. (2006). Aging increases the susceptibility of skeletal muscle derived satellite cells to apoptosis. Experimental Gerontology, 41, 828–836.
Sidorenko, A. V., Gubrii, I. B., Andrianova, L. F., Macsijuk, T. V., & Butenko, G. M. (1986). Functional rearrangement of lymphohemopoietic system in heterochronically parabiosed mice. Mechanism of Ageing and Development, 36, 41–56.
Hirayama, R., Takemura, K., Nihei, Z., Ichikawa, W., Takagi, Y., Mishima, Y., et al. (1993). Differential effect of host microenvironment and systemic humoral factors on the implantation and the growth rate of metastatic tumor in parabiotic mice constructed between young and old mice. Mechanism of Ageing and Development, 71, 213–221.
Fujino, H., Kohzuki, H., Takeda, I., Kiyooka, T., Miyasaka, T., Mohri, S., et al. (2005). Regression of capillary network in atrophied soleus muscle induced by hindlimb unweighting. Journal Applied Physiology, 98, 1407–1413.
Ryan, N. A., Zwetsloot, K. A., Westerkamp, L. M., Hickner, R. C., Pofahl, W. E., & Gavin, T. P. (2006). Lower skeletal muscle capillarization and VEGF expression in aged vs. young men. Journal Applied Physiology, 100, 178–185.
Goldspink, G., Fernandes, K., Williams, P. E., & Wells, D. J. (1994). Age-related changes in collagen gene expression in the muscles of mdx dystrophic and normal mice. Neuromuscular Disorders, 4, 183–191.
Alexakis, C., Partridge, T., & Bou-Gharios, G. (2007). Implication of the satellite cell in dystrophic muscle fibrosis: A self perpetuating mechanism of collagen over-production. American Journal of Physiology Cell Physiology.
Engler, A. J., Sen, S., Sweeney, H. L., & Discher, D. E. (2006). Matrix elasticity directs stem cell lineage specification. Cell, 126, 677–689.
Bischoff, R. (1986). A satellite cell mitogen from crushed adult muscle. Developments in Biologicals, 115, 140–147.
Chen, G., & Quinn, L. S. (1992). Partial characterization of skeletal myoblast mitogens in mouse crushed muscle extract. Journal of Cellular Physiology, 153, 563–574.
Yablonka-Reuveni, Z., Seger, R., & Rivera, A. J. (1999). Fibroblast growth factor promotes recruitment of skeletal muscle satellite cells in young and old rats. Journal of Histochemistry and Cytochemistry, 47, 23–42.
Sheehan, S. M., Tatsumi, R., Temm-Grove, C. J., & Allen, R. E. (2000). HGF is an autocrine growth factor for skeletal muscle satellite cells in vitro. Muscle Nerve, 23, 239–245.
Quinn, L. S., Ong, L. D., & Roeder, R. A. (1990). Paracrine control of myoblast proliferation and differentiation by fibroblasts. Developments in Biologicals, 140, 8–19.
Pampusch, M. S., Hembree, J. R., Hathaway, M. R., & Dayton, W. R. (1990). Effect of transforming growth factor beta on proliferation of L6 and embryonic porcine myogenic cells. Journal of Cellular Physiology, 143, 524–528.
Vandenburgh, H. H., Sheff, M. F., & Zacks, S. I. (1984). Soluble age-related factors from skeletal muscle which influence muscle development. Experimental Cell Research, 153, 389–401.
Mezzogiorno, A., Coletta, M., Zani, B. M., Cossu, G., & Molinaro, M. (1993). Paracrine stimulation of senescent satellite cell proliferation by factors released by muscle or myotubes from young mice. Mechanism of Ageing and Development, 70, 35–44.
Beggs, M. L., Nagarajan, R., Taylor-Jones, J. M., Nolen, G., Macnicol, M., & Peterson, C. A. (2004). Alterations in the TGFbeta signaling pathway in myogenic progenitors with age. Aging Cell, 3, 353–361.
Li, Y., Foster, W., Deasy, B. M., Chan, Y., Prisk, V., Tang, Y., et al. (2004). Transforming growth factor-beta1 induces the differentiation of myogenic cells into fibrotic cells in injured skeletal muscle: A key event in muscle fibrogenesis. American Journal of Pathology, 164, 1007–1019.
Florini, J. R., & Magri, K. A. (1989). Effects of growth factors on myogenic differentiation. American Journal Physiology, 256, C701–C711.
Reeves, I., Abribat, T., Laramee, P., Jasmin, G., & Brazeau, P. (2000). Age-related serum levels of insulin-like growth factor-I, -II and IGF-binding protein-3 following myocardial infarction. Growth Hormone & IGF Research, 10, 78–84.
Liu, H., Fergusson, M. M., Castilho, R. M., Liu, J., Cao, L., Chen, J., et al. (2007). Augmented Wnt signaling in a mammalian model of accelerated aging. Science, 317, 803–806.
Chen, Y., Zajac, J. D., & MacLean, H. E. (2005). Androgen regulation of satellite cell function. Journal Endocrinology, 186, 21–31.
Snyder, P. J. (2001). Effects of age on testicular function and consequences of testosterone treatment. Journal of clinical endocrinology and metabolism, 86, 2369–2372.
Renault, V., Rolland, E., Thornell, L. E., Mouly, V., & Butler-Browne, G. (2002). Distribution of satellite cells in the human vastus lateralis muscle during aging. Experimental Gerontology, 37, 1513–1514.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article
Cite this article
Brack, A.S., Rando, T.A. Intrinsic Changes and Extrinsic Influences of Myogenic Stem Cell Function During Aging. Stem Cell Rev 3, 226–237 (2007). https://doi.org/10.1007/s12015-007-9000-2
Published:
Issue Date:
DOI: https://doi.org/10.1007/s12015-007-9000-2