
Abstract
The composition of brackish groundwater from Brazilian backlands contains important elements necessary for metabolism in microalgae. This study evaluated the use of 100% brackish groundwater with different amounts of Zarrouk nutrients for Spirulina sp. LEB 18 cultivation. The growth parameters and biomass composition, including the concentrations of proteins, carbohydrates, ash, lipids, and fatty acids, were evaluated. The best growth parameter results were obtained in the assay using 100% brackish groundwater and only 25% of Zarrouk nutrients, which were equal to those obtained for the control culture. The concentrations of carbohydrates and polyunsaturated fatty acids were increased by as much as 4- and 3.3-fold, respectively, when brackish groundwater was used in the cultures. The lipid profile demonstrated that the biomass had the potential for use in biodiesel production. The use of brackish groundwater is a sustainable, economical way to obtain high-quality biomass for different applications during Spirulina sp. LEB 18 cultivation.
Similar content being viewed by others

Explore related subjects
Discover the latest articles, news and stories from top researchers in related subjects.Avoid common mistakes on your manuscript.
Introduction
Population growth and climate change threaten natural resources such as energy, food, and water [1]. Scarcity of water resources is a recurrent problem within the world population. Certain regions, such as the Brazilian backlands, have a semiarid climate and recurrent droughts, which leads to the accumulation of brackish and saline groundwater [2] that is difficult to use for human consumption. The use of most desalination processes is limited by the high cost of project implementation and maintenance. Although it is a serious natural resource problem, it is possible to find more feasible and lower-cost alternatives to mitigate the effects of the use of underground brackish water.
Spirulina is a photosynthetic microalga widely studied and commercially used due to its composition, which is beneficial to the human organism. Certain species can be composed of up to 70% protein and contain essential amino acids, vitamins such as B12, pigments, and essential fatty acids [3, 4]. In addition, carbohydrates and lipids derived from their biomass can be used in biofuel production [5, 6]. Microalgae generally require mineral salts for their growth, which are found in brackish groundwater [7]. Moreover, these microorganisms can be cultivated in adverse regions [8]. For these reasons, Spirulina cultivation in semiarid climates is possible due to the presence of water and nutrients in brackish groundwater that are required for its growth. Thus, cultivation costs would be reduced and biomass and water could be produced for consumption by humans, animals, and vegetation.
There are no reports about the use of brackish groundwater from Brazilian backlands for the growth of Spirulina cultures. Spirulina platensis that was cultivated in water containing 50% residual brackish groundwater desalination concentrate (DC) produced a biomass concentration that was 91.5% greater than that of the control that was cultivated in Paoletti culture medium [9]. Nannochloropsis gaditana cultivated in 75% DC produced a biomass concentration that was approximately 1.4 times greater than that of the control culture, which was cultivated in F/2 medium [10]. Chlorella vulgaris cultures cultivated in 25% DC had similar growth parameters to that of the control culture that was cultivated in BBM culture medium [11]. The present study aimed to evaluate the use of brackish groundwater from Brazilian backlands in cultures of Spirulina sp. LEB 18 in terms of growth parameters and biomass composition.
Materials and Methods
Microorganism and Culture Medium
The cyanobacteria Spirulina sp. LEB 18 used in this study was obtained from the Culture Collection of the Laboratory of Biochemical Engineering at the Federal University of Rio Grande (FURG) and was derived from Mangueira Lagoon in southern Brazil (latitude 32° 32′ 05″ S and longitude 33°31′ 57″ W). Brackish groundwater (composition given in Table 1) was used as the nutritional source for cultivation and was supplemented with different concentrations of Zarrouk [12] nutrients. The brackish groundwater was obtained from a desalination plant in the northeastern Brazilian backlands (Mandassaia II, Riachão do Jacuípe, Bahia, Brazil). The water was collected in plastic bottles, transported to the Federal University of Bahia (UFBA) and frozen at − 18 °C. Part of the brackish groundwater composition was determined following the Standard Methods (SM) for the examination of water and wastewater [13]: conductivity (SM 2510 A/B), salinity (SM 2520C), total dissolved solids (SM 2540C), total alkalinity (SM 2320 A/B), total hardness (SM 2340 A/B/C), sodium (SM 3120B), calcium (SM 3120B), magnesium (SM 3120B), potassium (SM 3120B), zinc (SM 3120B), and copper (SM 3120B). Nitrate, sulfates, bromide, chlorides, and ammonia were determined by ion chromatography.
Culture Conditions
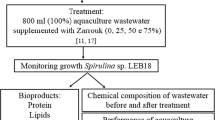
Prior to the experiments, the water was thawed at room temperature and autoclaved. The Spirulina sp. LEB 18 inoculums were centrifuged, and the resulting biomass was washed with distilled water and centrifuged again to remove salts. Erlenmeyer-type photobioreactors with a volume of 1 L were used. Under aseptic conditions, 800 mL of brackish groundwater as well as Spirulina sp. LEB 18 washed biomass (~ 0.3 g L−1) was added to the Erlenmeyer flasks. The experiments were performed in duplicate, during which the cultures were supplemented with different concentrations of Zarrouk nutrients (25, 50, 75, and 100%). Control experiments were performed without addition of brackish groundwater and using 100% Zarrouk culture medium. The duration of the experiments was 7 days, which were conducted in a growth chamber at 30 °C with a 12-h light/dark photoperiod. Fluorescent tubular lamps with a light intensity of 41.6 μmolphotons m−2 s−1 were used as the energy source [14]. Air that was filtered through glass wool in order to remove contaminants was used to agitate the cultures. The cultures were replenished with water daily prior to sample withdrawal.
Determination of Growth Parameters and pH
The biomass concentration of Spirulina sp. LEB 18 was determined daily by measuring the optical density of the cultures in a spectrophotometer at a wavelength of 670 nm. A standard curve was constructed prior to the experiments that correlated the optical density of the inoculum with its dry mass [15]. The growth parameters were determined daily for each of the cultures. Productivity was determined by the equation PX = (Xt − X0)/(t − t0), where Xt is the biomass concentration (g L−1) at time t (days) and X0 is the biomass concentration (g L−1) at time t0 (days). The maximum specific growth rate was calculated based on linear regression of the growth of the cultures during the log phase. The pH was also monitored daily with a digital pH meter.
Biomass Composition
The biomass was recovered from the liquid medium by centrifugation at the end of the experiments, washed with distilled water, and centrifuged again to remove the total salts. Subsequently, the biomasses were frozen in an ultrafreezer, lyophilized, and maintained at − 20 °C until analysis. The protein concentration was measured by Lowry, a colorimetric method using bovine serum albumin as a standard [16] after thermal and alkaline pretreatment of the microalgal biomass. The carbohydrate concentration was determined using the phenol-sulfuric method based on a standard glucose curve [17]. The biomass used for the analysis of proteins and carbohydrates was submitted to pre-treatment for cellular disruption. This procedure consisted of preparation of extract, containing 5 mg of biomass and 10 mL of distilled water, which was sonicated in an ultrasonic probe (COLE PARMER CPX 130, Illinois, USA) with 10 cycles of operation (1 min on and 1 min off ). The ash was measured based on the method described by the Association of Official Analytical Chemists [18]. The lipid concentration was determined by the Folch method [19]. The fatty acids were transmethylated and then separated and identified by gas chromatography equipped with a flame ionization detector (Clarus 680; Perkin Elmer®). The DB-FFAP column (30 m × 0.32 mm × 0.25 mm) and commercial fatty acid methyl esters (FAME) were used for the fatty acid identification. The injector and detector temperatures were 250 °C and 280 °C, respectively. The temperature program used was 150 °C for 16 min, increasing by 2 °C min−1 up to 180 °C (maintained for 25 min); increased by 5 °C min−1 up to 210 °C (maintained for 25 min). Helium (1.0 mL min−1) was used as the carrier gas during the analysis. FAME were identified by retention time comparison with a mixture of known patterns (C4–C24, 18, 919-AMP; Sigma-Aldrich®). The fatty acids were quantified by percentage (%) of total lipids and compared with internal standard methyl tricosanoate (C23:0, Sigma Aldrich®). The Clarus Chromatography Workstation software was used to determine the peak areas to normalize the percentage of total fatty acid [20, 21].
Theoretical Conversion of Biomass Carbohydrates to Ethanol
From the concentrations of carbohydrates in the biomass of Spirulina sp. LEB 18, calculations of the theoretical conversion of this biomolecule into ethanol were performed. Towards that, it was considered a calculation basis for 100 g of biomass, 0.511 of constant theoretical stoichiometric conversion of glucose to ethanol, and 70% efficiency of ethanol formation [22, 23].
Statistical Analysis
The statistical analysis was conducted based on analysis of variance followed by Tukey’s test at a 95.0% confidence level.
Results and Discussion
Growth Parameters
The brackish groundwater was rich in chlorides, sodium, and calcium. In addition, other important elements for the metabolism of microalgae were detected, such as nitrate, phosphate, magnesium, potassium, zinc, and copper (Table 1). Ammonia present in brackish groundwater can be toxic to microalgae cultures, depending on its concentration [11]. However, the growth of Spirulina sp. LEB 18 cultures cultivated in brackish groundwater with a very low ammonia concentration (0.98 mg L−1) was not inhibited. The maximum biomass concentration obtained for Spirulina sp. LEB 18 cultures cultivated in 100% brackish groundwater supplemented with 25% and 75% of Zarrouk nutrients were the same (p < 0.05) as those obtained for the control cultures (Table 2). In addition, cultures cultivated in 100% brackish groundwater supplemented with 25, 50, and 75% of Zarrouk nutrients had the same productivities (p < 0.05) as the control culture. The maximum specific growth rates were the same (p < 0.05) for cultures grown in 100% brackish groundwater containing 25, 50, or 75% Zarrouk nutrients. The cultures cultivated in 100% of brackish groundwater with 100% Zarrouk nutrients supplementation had no increase in biomass during cultivation, most likely due to the high amount of salts that were present. However, the rest of the brackish groundwater cultures showed growth from the first day of culture that ended at 10 days, still during the exponential phase of cell growth (approximately 1 g L−1; Fig. 1).

Spirulina sp. LEB 18 biomass concentrations obtained during cultivation with brackish groundwater and different percentages of Zarrouk culture medium: (ο) (control); (+) 25%; (□) 50%; (Δ) 75% and (∇) 100%
The Zarrouk medium, ideal for the growth of Spirulina sp. LEB 18, has a high salt concentration. Thus, the high salinity of the brackish groundwater used during cultivation may not have had an impact on the adapted microalgal cells, which explains the absence of a physiological adaptation in the cultures supplemented with 25, 50, and 75% Zarrouk nutrients. These results demonstrate the potential of brackish groundwater as a basic water source during Spirulina sp. LEB 18 cultivation. According to Borowitzka and Moheimani [24], the use of this type of water is the most sustainable way to carry out microalgae cultivation on a large scale, as it does not compete with the use of freshwater for agriculture.
Matos et al. [10] cultivated Nannochloropsis gaditana microalga using DC and obtained the greatest biomass concentration in the cultures with 75% of F/2 culture medium and only 25% of DC. Volkmann et al. [9] cultivated Spirulina platensis with 100% of DC and 50% of Paoletti culture medium and obtained biomass concentrations similar to those produced during the present study (~ 1 g L−1). However, in the present study, the best results were obtained when using 100% brackish groundwater and Zarrouk nutrients supplementation of only 25%, which is advantageous in terms of reducing nutrient costs during cultivation.
The initial pH values of the brackish groundwater cultures were similar to those of the control culture. The pH values of the cultures ranged from 9.25–10.30 (control), 9.15–10.05 (25% supplementation), 9.18–10.13 (50% supplementation), 9.27–10.19 (75% supplementation), and 9.3–8.5 (100% supplementation). With the exception of the culture with 100% Zarrouk nutrients supplementation, the rest of the cultures tended to show an increase in pH during cultivation due to the microalgal growth. The pH ranges of the brackish groundwater cultures are within the values considered optimal for the growth of species from the genus Spirulina [25].
Biomass Composition
The final biomass composition of Spirulina sp. LEB 18 cultures containing 100% brackish groundwater was influenced by the concentrations of Zarrouk nutrients that was used (Fig. 2). The main biomolecules that were affected were proteins and carbohydrates. The protein content in the final biomass was inversely proportional to the concentration of Zarrouk nutrients used, while the content of carbohydrates was proportional to the concentration of Zarrouk nutrients. The biomass obtained in the culture containing 100% brackish groundwater and 75% Zarrouk nutrients had approximately 3-fold more carbohydrates and 4-fold less proteins when compared with the control culture. In addition, even cultures supplemented with only 25% Zarrouk nutrients had a carbohydrate production that was increased approximately 2.7-fold.

Biomass composition of Spirulina sp. LEB 18 cultures grown in 100% brackish groundwater and different percentages of Zarrouk culture medium. The presence of the same letters for each macromolecule means no significant difference between the values at a 95% confidence level (p > 0.05)
Leema et al. [26] also observed a reduction in proteins and an increase in carbohydrates in seawater cultures of Spirulina platensis. However, in the present study, cultures containing 100% brackish groundwater had an increase in carbohydrates that was approximately 2-fold greater than that of the seawater cultures cultivated by Leema et al. [26].
The carbohydrates produced by microalgae are intended to serve as a structural component in cell walls and as a storage component within the cell. As storage biomolecules, carbohydrates provide the energy required for metabolism in microalgae subject to stress. Thus, the increase in the saline concentration in the cultures may have caused the elevation of osmotic pressure in the medium and, consequently, led to an increase in carbohydrate production in order to protect the cells [27,28,29]. The biomasses with high amounts of carbohydrates that were obtained from the Spirulina sp. LEB 18 cultures grown in brackish groundwater have the potential to be used for bioethanol production. While in the control cultures the bioethanol production was 6.35 mL, in the cultures with brackish groundwater, this production quadrupled to 25.84 mL (assay with 100% brackish groundwater with supplementation of 75% Zarrouk nutrients). The main advantage in obtaining biofuels from microalgal biomass is that, unlike biofuels from terrestrial lignocellulosic plants, their carbohydrates do not contain lignin, and the cells are not usually recalcitrant, which reduces the processing and costs associated with pretreatment [30]. To obtain reserve biomolecules (carbohydrates and lipids) from microalgal biomass, the cultures are usually subjected to stress, which leads to reduced productivity in many cases [31,32,33]. However, in this study, the biomasses contained high levels of carbohydrates and had similar productivity to that of the control culture, which allows for the production of bioethanol from microalgae cultivated in brackish groundwater to be quite advantageous.
Spirulina sp. LEB 18 cultures cultivated in brackish groundwater had varying protein concentrations within their biomass (20 to 40%) and were smaller (p > 0.05) than the control culture. Matos et al. [11] also observed a decrease in the amount of protein in Chlorella vulgaris biomass when it was cultivated with DC. The reduction in protein may be related to the redirection of energy to processes such as osmoregulation instead of the synthesis of biomolecules [34]. The protein fraction in Spirulina biomass is recognized for its excellent composition in terms of essential amino acids [35]. In addition, when compared with traditional protein sources such as meat or soybeans, microalgae biomass has the advantages of rapid cell growth, reduced culture costs, and a lack of competition with other types of food production. In this way, the use of Spirulina sp. LEB 18 as a protein source for the human diet is a sustainable alternative that can be used to satisfy growing worldwide demand. In addition, microalga proteins can be used for the production of biofilms, emulsifiers, and other products [36,37,38].
The lipid concentration in the biomass obtained from Spirulina sp. LEB 18 cultures cultivated in 100% brackish groundwater and 25 and 50% of Zarrouk nutrients was not affected and was equal to that of the control culture (p < 0.05). However, a reduction (p > 0.05) in lipids was observed in the cultures grown in 100% brackish groundwater and 75% Zarrouk nutrients, which may have occurred due to changes in metabolism that prioritized the production of carbohydrates. Conversely, the concentration of fatty acids in the cultures grown in 100% brackish groundwater and 75% Zarrouk nutrients was 3.4-fold greater than that in the control culture (Table 3). In general, cultures grown in brackish groundwater had higher concentrations of fatty acids than the control culture. According to Fon Sing et al. [39], in order to adjust to changes in salinity associated with membrane permeability in cells, microalgae activate an adaptive osmoregulatory mechanism, which leads to a greater production of saturated fatty acids. In addition, polyunsaturated fatty acid (PUFA) concentrations in the cultures grown in brackish groundwater were higher than those in the control culture and increased in proportion to the supplementation with the Zarrouk medium. Fujii et al. [40] also observed that PUFA concentrations increased proportionally to the salinity of the medium and suggested that the enzymes responsible for the increase in the production of these biomolecules were involved. PUFAs are very important for human and animal health and are widely used to prevent and treat heart and inflammatory diseases and for nutritional supplementation [41,42,43].
For biodiesel production, it is important that the lipid composition consists of saturated and long fatty acids, which provide biofuel with greater calorific value, cetane number, and oxidative stability [44]. In addition, shorter and unsaturated fatty acids are also important for biodiesel production, as they contribute to the characteristics of viscosity and flow at low temperatures [45]. Therefore, considering that all of the cultures grown in brackish groundwater contained more than 50% saturated fatty acids, in addition to short and unsaturated fatty acids, it has been verified that the lipids derived from these cultures are strong candidates for use in biodiesel production.
The amount of ash contents was the same in all of the cultures (p < 0.05), most likely due to the washing performed at the end of the cultivation period, which removed the mineral salts that were present. In the cultures cultivated in 100% brackish groundwater and 100% Zarrouk nutrients, there was no biomass production, so it was not possible to characterize the presence of biomolecules.
Conclusion
The productivities of Spirulina sp. LEB 18 cultures with 100% brackish groundwater were equal to the control assays. The production of carbohydrates and polyunsaturated fatty acids by the microalga was increased by as much as 4- and 3.3-fold, respectively. The lipid composition of the biomass had an ideal combination of saturated and unsaturated fatty acids, which is important for biodiesel production. It was possible to demonstrate the potential benefits of the use of brackish groundwater in the cultivation of Spirulina sp. to obtain high-quality biomass that could be utilized for different biotechnological purposes.
References
McNeill, K., Macdonald, K., Singh, A., & Binns, A. D. (2017). Food and water security: Analysis of integrated modeling platforms. Agricultural Water Management, 194, 100–112.
Sánchez, A. S., Nogueira, I. B. R., & Kalid, R. A. (2015). Uses of the reject brine from inland desalination for fish farming, Spirulina cultivation, and irrigation of forage shrub and crops. Desalination., 364, 96–107.
Belay, A., Kato, T., & Ota, Y. (1996). Spirulina (Arthrospira): potential application as an animal feed supplement. Journal of Applied Phycology, 8(4-5), 303–311.
Vonshak, A. (1997). Spirulina platensis (Arthrospira): Physiology, Cell Biology and Biotechnology. London: Taylor and Francis.
Silva, C. E. F., & Bertucco, A. (2016). Bioethanol from microalgae and cyanobacteria: A review and technological outlook. Process Biochemistry, 51(11), 1833–1842.
Shirazi, H. M., Sabet, J. K., & Ghotbi, C. (2017). Biodiesel production from Spirulina microalgae feedstock using direct transesterification near supercritical methanol condition. Bioresource Technology, 239, 378–386.
Matos, A. P., Moecke, E. H. S., & Sant’Anna, E. S. (2017). The use of desalination concentrate as a potential substrate for microalgae cultivation in Brazil. Algal Research, 24, 505–508.
Singh, A., & Olsen, S. I. (2011). A critical review of biochemical conversion, sustainability and life cycle assessment of algal biofuels. Applied Energy, 88(10), 3548–3555.
Volkmann, H., Imianovsky, U., Oliveira, J. L. B., & Sant’Anna, E. S. (2007). Cultivation of Arthrospira (Spirulina) platensis in desalinator wastewater and salinated synthetic medium: Protein content and amino-acid profile. Brazilian Journal of Microbiology, 39, 98–101.
Matos, A. P., Feller, R., Moecke, E. H. S., & Sant’Anna, E. S. (2015). Biomass, lipid productivities and fatty acids composition of marine Nannochloropsis gaditana cultured in desalination concentrate. Bioresource Technology, 197, 48–55.
Matos, A. P., Ferreira, W. B., Torres, R. O. C., Morioka, L. R. I., Canella, M. H. M., Rotta, J., Silva, J. T., Moecke, E. H. S., & Sant’Anna, E. S. (2014). Optimization of biomass production of Chlorella vulgaris grown in desalination concentrate. Journal of Applied Phycology, 27, 1473–1483.
Costa, J. A. V., Colla, L. M., & Filho, P. D. (2004). Improving Spirulina platensis biomass yield using a fed-batch process. Bioresource Technology, 92(3), 237–241.
APHA. (2005). Standard Methods for the examination of water and wastewater. Washington DC: American Public Health Association/American Water Works Association/Water Environment Federation.
Morais, M. G., & Costa, J. A. V. (2007). Carbon dioxide fixation by Chlorella kessleri, C. vulgaris, Scenedesmus obliquus and Spirulina sp. cultivated in flasks and vertical tubular photobioreactors. Biotechnology Letters, 29(9), 1349–1352.
Costa, J. A. V., Colla, L. M., Duarte Filho, P., Kabke, P. K., & Weber, A. (2002). Modelling of Spirulina platensis growth in fresh water using response surface methodology, World J. Microbial Biotechnology, 18(7), 603–607.
Lowry, O. H., Rosebrough, N. J., Farr, A. L., & Randall, R. J. (1951). Protein measurement with the Folin phenol reagent. The Journal of Biological Chemistry, 193(1), 265–275.
Dubois, M., Gilles, K. A., Hamilton, J. K., Rebers, P. A., & Smith, F. (1956). Colorimetric method for determination of sugars and related substances. Analytical Chemistry, 28(3), 350–356.
AOAC (Ed.). (1995). Official Methods of Analysis of the association of analytical chemists international (16th ed.). Arlington: AOAC International.
Folch, J., Lees, M., & Stanley, G. H. S. (1957). A simple method for the isolation and purification of total lipids from animal tissues. The Journal of Biological Chemistry, 226(1), 497–509.
Andrade, B. B., Cardoso, L. G., Assis, D. J., Costa, J. A. V., Druzian, J. I., & Lima, S. T. C. (2019). Production and characterization of Spirulina sp. LEB 18 cultured in reused Zarrouk’s medium in a raceway-type bioreactor. Bioresource Technology, 284, 340–348.
Nascimento, I. A., Marques, S. S. I., Cabanelas, I. T. D., Carvalho, G. C., Nascimento, M. A., Souza, C. O., Druzian, J. I., Hussain, J., & Liao, W. (2014). Microalgae versus land crops as feedstock for biodiesel: productivity, quality and standard compliance. Bioenergy Research, 7, 1002–1013.
Freitas, B. C. B., Bracher, E. H., Morais, E. G., Atala, D. I. P., Morais, M. G., & Costa, J. A. V. (2017). Cultivation of different microalgae with pentose as carbon source and the effects on the carbohydrate content. Environmental Technology, 40, 1062–1070.
Rosa, G. M., Moraes, L., Cardias, B. B., Souza, M. R., & Costa, J. A. V. (2015). Chemical absorption and CO2 biofixation via the cultivation of Spirulina in semicontinuous mode with nutrient recycle. Bioresource Technology, 192, 321–327.
Borowitzka, M. A., & Moheimani, N. R. (2013). Sustainable biofuels from algae. Mitigation and Adaptation Strategies for Global Change, 18, 13–25.
Tomaselli, L. (1997). Morphology, ultrastructure and taxonomy of Arthrospira (Spirulina). In A. Vonshak (Ed.), Spirulina platensis (Arthrospira) Physiology, cell biology and biotechnology (pp. 01–16). London: Taylor & Francis.
Leema, J. T. M., Kirubagaran, R., Vinithkumar, N. V., Dheenan, P. S., & Karthikayulu, S. (2010). High value pigment production from Arthrospira (Spirulina) platensis cultured in seawater. Bioresource Technology, 101(23), 9221–9227.
Markou, G., Angelidaki, I., & Georgakakis, D. (2012). Microalgal carbohydrates: an overview of the factors influencing carbohydrates production, and of main bioconversion technologies for production of biofuels. Applied Microbiology and Biotechnology, 96(3), 631–645.
Geider, R. J., & La Roche, J. (2002). Redfield revisited: variability of C:N:P in marine microalgae and its biochemical basis. European Journal of Phycology, 37(1), 1–17.
Raven, J. A., & Beardall, J. (2004). Carbohydrate metabolism and respiration in algae. In A. W. D. Larkum, S. E. Douglas, & J. A. Raven (Eds.), Photosynthesis in algae, Advances in photosynthesis and respiration (pp. 205–224). Dordrecht: Springer.
Li, K., Liu, S., & Liu, X. (2014). An overview of algae bioethanol production. International Journal of Energy Research, 38(8), 965–977.
Ma, Y., Gao, Z., Wang, Q., & Liu, Y. (2018). Biodiesels from microbial oils: Opportunity and challenges. Bioresource Technology, 263, 631–641.
Griffiths, M. J., & Harrison, S. T. (2009). Lipid productivity as a key characteristic for choosing algal species for biodiesel production. Journal of Applied Phycology, 21(5), 493–507.
Paliwal, C., Mitra, M., Bhayani, K., Bharadwaj, A. V. V., Ghosh, T., Dubey, S., & Mishra, S. (2017). Abiotic stresses as tools for metabolites in microalgae. Bioresource Technology, 244(Pt 2), 1216–1226.
Kirst, G. O. (1989). Salinity tolerance of eukaryotic marine algae. Annual Review of Plant Physiology and Plant Molecular Biology, 41, 21–53.
Garcia, J. M. R., Fernández, F. G. A., & Sevilla, J. M. F. (2012). Development of a process for the production of L-amino-acids concentrates from microalgae by enzymatic hydrolysis. Bioresource Technology, 112, 164–170.
Zhang, X., Yuan, H., Jiang, Z., Lin, D., & Zhang, X. (2018). Impact of surface tension of wastewater on biofilm formation of microalgae Chlorella sp. Bioresource Technology, 266, 498–506.
Ye, Y., Huang, Y., Xia, A., Fu, Q., Liao, Q., Zeng, W., Zheng, Y., & Zhu, X. (2018). Optimizing culture conditions for heterotrophic-assisted photoautotrophic biofilm growth of Chlorella vulgaris to simultaneously improve microalgae biomass and lipid productivity. Bioresource Technology, 270, 80–87.
Singh, R., Parihar, P., Singh, M., Badguz, A., Kumar, J., Singh, S., Singh, V. P., & Prasad, S. M. (2017). Uncovering potential applications of cyanobacteria and algal metabolites in biology, agriculture and medicine: current status and future prospects. Frontiers in Microbiology, 8, 1–37.
Fon Sing, S. F., Isdepesky, A., Borowitzka, M. A., & Lewis, D. M. (2014). Pilot scale continuous recycling of growth medium for the mass culture of a halotolerant Tetraselmis sp. in raceway ponds under increasing salinity: a novel protocol for commercial microalgal biomass production. Bioresource Technology, 161, 47–54.
Fujii, S., Uenaka, M., Nakayama, S., Yamamoto, R., & Mantani, S. (2001). Effects of sodium chloride on the fatty acids composition in Boekelovia hooglandii (Ochromonadales, Chrysophyceae). Phycological Research, 49(1), 73–77.
Sun, X. M., Geng, L. J., Ren, L. J., Ji, X. J., Hao, N., Chen, K. Q., & Huang, H. (2018). Influence of oxygen on the biosynthesis of polyunsaturated fatty acids in microalgae. Bioresource Technology, 250, 868–876.
Plouguerné, E., Da, G. B., Pereira, R. C., & Barretobergter, E. (2014). Glycolipids from seaweeds and their potential biotechnological applications. Frontiers in Cellular and Infection Microbiology, 4, 174.
Ruxton, C. H., Calder, P. C., Reed, S. C., & Simpson, M. J. (2005). The impact of long chain n-3 polyunsaturated fatty acids on human health. Nutrition Research Reviews, 18, 113–129.
Kondamudi, N., Strull, J., Misra, M., & Mohapatra, S. (2009). A green process for producing biodiesel from feater meal. Journal of Agricultural and Food Chemistry, 57(14), 6163–6166.
Knothe, G. (2005). Dependence of biodiesel fuel properties on the structure of fatty acid alkyl esters. Fuel Processing Technology, 86, 1059–1070.
Acknowledgments
The authors acknowledge CAPES (Coordination for the Improvement of Higher Education Personnel), MCTIC (Ministry of Science Technology, Innovation and Communications), and the Program to Support the Production of Academic Publications/PROPESP/FURG/2018.
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Conflict of Interest
The authors declare that they have no conflict of interest.
Additional information
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
About this article

Cite this article
Duarte, J.H., Cardoso, L.G., de Souza, C.O. et al. Brackish Groundwater from Brazilian Backlands in Spirulina Cultures: Potential of Carbohydrate and Polyunsaturated Fatty Acid Production. Appl Biochem Biotechnol 190, 907–917 (2020). https://doi.org/10.1007/s12010-019-03126-7
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s12010-019-03126-7