
Abstract
A novel alkaline protease was purified from Pseudomonas aeruginosa HY1215 using ammonium sulfate, DEAE-Sepharose and Sephacryl S-200 gel filtration chromatographic techniques. The protease had a relative molecular weight of 32.8 KDa by SDS-PAGE, and the optimal temperature and pH for excellent stability and activity were determined as 25 °C and 10.0, respectively. Within the pH range of 7.0–11.0, the protease had a good stability, which could retain more than 80 % of its original activity; in the temperature range of 15–35 °C, the protease had a higher activity, and its activity at 20 °C could amount to 85 % of the maximum activity at 25 °C. Besides, the enzyme activity showed a valuable stability towards several commercially available surfactants (Tween-80, Tween-40, and Triton X-100) and bleaches (H2O2) even when their concentrations ranged up to 2.0 and 1.6 %.
Similar content being viewed by others


Explore related subjects
Discover the latest articles, news and stories from top researchers in related subjects.Avoid common mistakes on your manuscript.
Introduction
Microbial proteases constitute one of the most important groups of industrial enzymes, which account for more than 60 % of the total industrial enzyme market [1]. Among the protease group, alkaline protease is of particular interest due to the primary applications as a cleaning additive in the detergent industry since its introduction in 1914 [2–4]. According to a released Business Communications Company, Inc. Study Global Markets for Enzymes in Industrial Applications, the global market for industrial enzymes was valued at $3.1 billion in 2009 and reached about $3.6 billion in 2010. The estimated market for 2016 is about $6 billion.
Alkaline protease in the laundry detergents plays a specific catalytic role in the hydrolysis of protein strains such as blood, milk, human sweat, etc. Due to its cleaning capability, this protease has been widely used as a detergent additive in environmentally acceptable nonphosphate detergents. The performance of alkaline protease is influenced by several factors such as pH of washing solution, wash temperature, and detergent composition. An ideal detergent enzyme should be stable over a broad pH range and a wide temperature zone and can withstand oxidizing and chelating agents. Meanwhile, it should be effective on cleaning clothing even at low enzyme level in detergent solutions. Moreover, considering mass production, it should also have broad substrate specificity [2].
In recent years, alkaline proteases from high yield strains have been studied extensively. However, very few published reports are available on the compatibility of alkaline proteases with detergent ingredients and cold activity [5–7]. In this article, a novel antisurfactant and antioxidant alkaline protease with cold adaptability was purified from Pseudomonas aeruginosa HY1215, and this protease could be evaluated as a cold washing detergent enzyme.
Materials and Methods
Strains and Culture Conditions
The psychrophilic strain HY1215 was isolated from Yellow Sea in China and was identified as P. aeruginosa. The psychrophilic bacterium were cultivated in PM liquid medium (1 % glucose, 3.5 % bean cake meal, 0.4 % Na2HPO4, 0.03 % KH2P04, 0.1 % Na2CO3, 0.02 % MgSO4, 0.2 % CaCl2, initial pH of 7) at 20 °C with stirring of 200 rpm for 36 h.
Purification Procedures
All the fractionations were done at 4 °C unless otherwise specified, and the chromatography purification for protein was monitored by measuring absorbance at the wavelength of 280 nm.
Step 1. The culture solution was centrifuged at 4000 rpm for 40 min at 4 °C to remove cells and insoluble residues, and the supernatant was transferred to ammonium sulfate of 80 % saturation. The resulting precipitate was dissolved in 25-mM borate buffer (pH 10.0) and then dialyzed against 2 L of the same buffer for 48 h, with four intervals at every 12 h.
Step 2. The dialyzed enzyme was loaded onto a cation exchanger (DEAE-Sepharose, Pharmacia, Sweden, 1.5 cm × 20 cm) equilibrated with 25-mM borate buffer (pH 10.0), and the flow rate was adjusted to 1 ml min−1. The enzyme was eluted from the column using a linear NaCl gradient (0–1.0 M), and the eluted fractions were collected. These fractions were precipitated by addition of 0.5 volume of chilled ethanol, and the precipitated enzyme was removed by centrifugation at 4000 rpm for 40 min. The partially purified enzyme was used for further purification.
Step 3. The protease solution prepared in Step 2 was loaded onto a Sephacryl S-200 gel filtration column (Pharmacia, Sweden, 1 cm × 80 cm) equilibrated with 25-mM borate buffer (pH 10.0). Fractions of the elution were collected at a flow rate of 0.2 ml min−1, and the active protease fractions were pooled and stored at−20 °C.
Enzyme Assay
Protease activity was assayed by measuring the absorbance at 660 nm of the substance liberated from the reaction of a casein solution with Folin-Ciocalteu’s reagent. 0.1-ml enzyme (pH = 10.0) was added to 5.0 ml 0.6 % casein solution containing 25-mM borate buffer, followed by incubation at 25 °C for 10 min. The enzyme reaction was terminated by addition of 5.0-ml reagent containing 0.33 M CH3COOH, 0.11 M CCl3COOH, and 0.22 M CH3COONa. Then, the mixture was filtered through filter paper, and 2.0-ml filtrate was mixed with 5.0-ml Na2CO3 of 0.55 M and 1.0-ml Folin-Ciocalteu’s reagent of 0.6 N. Afterwards, the mixture was placed at 25 °C for 30 min, and the absorbance at 660 nm was measured. Here, one unit enzyme activity was defined as the amount of enzyme that liberated 1-μg tyrosine per minute under these conditions.
Protein Estimation
Protein concentration was measured by the method of Bradford using bovine serum albumin (fraction V; Sigma) as a standard [8].
Characterization
The protease activity at various pH values was measured using casein as the substrate. The reaction pH values were adjusted to 6.0–12.0 with the buffer systems of Tris-maleate (pH 6–7), Tris-HCl (pH 7.0–8.0), and glycine-NaOH (pH 9.0–12.0). The stability of alkaline protease on pH variation was determined in different buffers after incubating the solutions at 25 °C for 1 h, and the residual protease activities were subsequently determined as described above at the pH of 10.0. The optimum temperature was determined with casein as substrate at different temperatures (from 0–40 °C) in 25-mM borate buffer (pH = 10.0). The effect of pH on the protease stability was evaluated by incubating the enzyme at different pH values for 24 h. To test stability at different temperatures, the enzyme was incubated at the desired temperature for 30 min at the pH of 10.0. Afterwards, residual activity was determined for each sample. The effects of various chemical reagents and metal ions on enzyme activity were examined by incubating the protease solution (in 25-mM borate buffer, pH = 10.0) with different compounds at 25 °C for 1 h, and the relative activities were determined under standard assay conditions.
Results and Discussion
Purification of Protease from P. aeruginosa HY1215
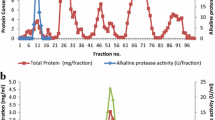
The purification result of protease was summarized in Table 1. A 10-fold purification was achieved with an overall yield of 60 % from the culture supernatant. SDS-PAGE of the purified enzyme presented a single band corresponding to the position of MW 32,800 (Fig. 1). Generally, the molecular mass of alkaline proteases from microorganisms ranges from 15 to 30 kDa [9] other than a few reports of higher molecular mass, such as 31.2 kDa [10], 33 kDa [11], 36 kDa [12], and 45 kDa [13]. However, an enzyme from Kurthia spiroforme had an extremely low molecular weight of 8 kDa [14].

SDS-PAGE of protease during purification steps
Lanes 1, molecular mass markers; Lanes 2, precipitate with 60 % saturated ammonium sulfate; Lanes 3, precipitate with 80 % saturated ammonium sulfate; Lanes 4, fraction from the DEAE-Sepharose; Lanes 5, fraction from the Sephacryl S-200.
Effects of pH and Temperature on the Activity and Stability
Since the pH of laundry detergents is commonly in the range of 9.0–11.0 [15], protease and other enzymes in current detergent formulations should be alkaline with a high pH in nature. The protease from P. aeruginosa HY1215 showed an optimum pH of 10.0 and a relatively high activity in a broad pH range of 7.0–11.0 (Fig. 2). One of the most important features of the protease from P. aeruginosa HY1215 was its maximum activity (pH = 10) and the excellent stability in the alkaline pH range up to 11.0. Generally, the optimum pH range of alkaline proteases is from 8.0 to 12.0 [16–18, 12], with a few exceptions of higher pH optima. For example, two alkaline proteases from alkalophilic Bacillus sp. NCDC, Ap-1 and Ap-2 were optimally active at the pH of 11.0 and 12.0, respectively [13]. Besides, an alkaline protease from Bacillus sp. AH-101 was reported to retain 80 % enzyme activity in the pH range of 5.0–9.0 while 90 % in the pH range of 10.0–11.5 [19].

Effect of pH variation on enzyme activity at 25 °C
The enzyme exhibited an optimum performance at 25 °C, with relatively high activity over a broad temperature range of 10–35 °C (Fig. 3). The enzyme activity at 10 and 20 °C amounted to 70 and 85 % of that at 25 °C, respectively, which meant that lower wash temperatures were allowed when the enzyme was added to detergents. Compared with the mesophilic subtilisin Carlsberg (60 °C), the Antarctic subtilisin S41 (40 °C), the commercial Alcalase®, and the commercial Savinase®, the present protease had a low optimum temperature (25 °C), and a high relative activity at 0 °C could be retained as 29 % of the maximum activity at 25 °C (Table 2). Therefore, it was a true psychrophilic enzyme, and both kinetically and thermodynamically adapted to the cold natural habitat of the bacterium.

Effect of temperature on enzyme activity at the pH of 10
In the pH range from 7 to 11 (Fig. 4a), the enzyme showed good stability, and more than 80 % of the activity could be retained, with the maximum activity at the pH of 10. Thermal stability of the protease was monitored by measuring the residual enzymatic activity after incubation for 30 min in 25-mM borate buffer (pH 10.0), and the results showed that protease was sensitive to temperature inactivation above 35 °C, which lost about 40 % of its activity at 45 °C (Fig. 4b).

Effects of pH (a) and temperature (b) on stability of purified protease. 4 °C (squares); 25 °C (triangles)
Effects of Various Chemicals on the Protease Activity
The effect of EDTA on the activity of protease was examined, and at least 15 mM EDTA was required to inhibit more than 30 % of the protease activity, indicating there was no requirement for metal cofactors. The detergents contained high amounts of sequestering agents, and the agents could chelate the metal ions and make them unavailable in the washing solution.
The effects of metal ions on the protease activity were investigated, and the results were shown in Table 3. The protease could be activated by Mg2+, K+, Ca2+, Ba2+, and Zn2+, of which the Ca2+ and Zn2+ were very strong activators. In the presence of Ca2+ (1 mM) and Zn2+ (2 mM), the activities of the protease were respectively 21 and 15 % higher than that observed in the control solution. In contrast, the protease activity was completely inhibited in the presence of Ag+ (10 mM).
The enzyme activity was potently inhibited by phenylmethanesulfonyl fluoride (PMSF), suggesting the involvement of serine residues in the enzyme reaction.
The alkaline protease from P. aeruginosa HY1215 was very stable towards both common surfactants and bleaches. As Fig. 5a shows, in the presence of Tween-80, Tween-40, and Triton-X100 at the concentration of 2 % after incubation for 1 h at 25 °C, the enzyme activity was highly stable, together with appreciable oxidation stability. At an oxidizing agent concentration of 0.4 % H2O2, the protease activity could be increased by up to 45 %, and the enzyme still showed a striking stability towards the bleaches with increasing concentrations of oxidizing agents (Fig. 5b). For most proteases, the oxidizing agent H2O2 would seriously inhibit the enzyme activity. According to earlier reports on the stability of alkaline proteases toward oxidants, an alkaline protease from Bacillus sp. RGR-14 showed 40 % loss of enzyme activity with 1 % H2O2 [24], while a subtilisin-like protease from Bacillus sp. KSMKP43 lost little or no enzyme activity with 10 % H2O2 treatment for 30 min [25]. Combining these results, the Pseudomonas sp. HY1215 protease exhibited a significant compatibility and stability toward both surfactants and oxidizing agents. In general, the oxygen could oxidize the Cys residue and make the enzyme lost their activity, but the present protease had a distinct antioxygenation. Maybe there was no Cys residue in this protease at all, so that this protease had an antioxygenation. Nevertheless, the exact reason for this phenomenon was not clear, and the specific mechanism still needed more evidence. Ideally, proteases used as detergent additives should tolerate oxidizing chemicals and surfactants. However, not all of currently used detergent enzymes can remain active in the presence of bleaching chemicals. Hence, the latest trend in enzyme-based detergents is to introduce site-directed mutagenesis and protein engineering techniques to produce enzymes with better antioxidization and heat stability [26]. For instance, Kulakova reported that they improved the stability of cold active serine alkaline protease from the psychrotrophic bacterium Shewanella sp. strain Ac 10 by rational mutagenesis [27]. Notably, the present P. aeruginosa alkaline protease has already possessed good stability without any subsequent treatment.

Effects of surfactants (a) and bleach (b) on activity of purified protease. a Tween-40 (squares), Tween-80 (diamonds), Triton-X100 (triangles); b H2O2 (diamonds)
In summary, the current study showed that the alkaline protease secreted by HY1215 was a useful enzyme for detergent formulations, especially in the cold washing industry. The protease had high proteolytic activity at the pH of 10.0 at the cold temperature of 25 °C, and its stability towards various surfactants and bleaches was also desirable. Our results suggested that this alkaline protease was suitable for inclusion in detergent compositions and was consistent with the trend of lowering washing temperature and reducing energy consumption. However, before its industrial application, further studies such as its gene isolation and the detailed characterization of enzymes including substrate specificity are still needed.
References
Ponnambalam, S., Neelamegam, A., Ayyappan, S., & Thangavel, B. (2012). Biologia, 67(4), 629–635.
Anwar, A., & Mohammed, S. (1998). Bioresource Technology, 64, 139–144.
Erikson, N. (1996). Industrial enzymology. 2nd. The Macmillan Press Ltd. London.
Gupta, R., Beg, Q. K., & Lorenz, P. (2002). Applied Microbiology and Biotechnology, 59, 15–32.
Zhu, H. Y., Tian, Y., Hou, Y. H., & Wang, T. H. (2009). Molecular Biology Reports, 36(8), 2169–2174.
Wang, F., Hao, J. H., Yang, C. Y., & Sun, M. (2010). Applied Biochemistry and Biotechnology, 162(5), 1497–1505.
Iram, S., Parvaiz, H. Q., & Shabir, A. R. (2012). India. World Journal of Microbiology and Biotechnology, 28(3), 1071–1079.
Lowry, O. H., Rosebrough, N. J., Farr, A. L., & Randall, R. J. (1951). Journal of Biological Chemistry, 193, 265–275.
Fogarty, W. M., Griffin, P. J., & Joyce, A. M. (1974). Process Biochemistry, 9, 27–29.
Stephane, D., Georges, F., Emmanuel, N., & Charles, G. (1994). Journal of Biological Chemistry, 26, 17448–17453.
Samal, B. B., Karan, B., Parker, C., & Stabinsky, Y. (1991). Enzyme and Microbial Technology, 13, 66–70.
Kasana, R. C., & Yadav, S. K. (2007). Current Microbiology, 54, 224–229.
Kumar, C. G., Tiwari, M. P., & Jany, K. D. (1999). Process Biochemistry, 34, 441–449.
Steel, D. B., Fiske, M. J., Steele, B. P., & Kelley, V. C. (1992). Enzyme and Microbial Technology, 14, 358–360.
Banerje, U. C., Sani, R. K., & Azmi, W. (1999). Process Biochemistry, 35, 213–219.
Beg, K. B., Sahai, V., & Gupta, R. (2003). Process Biochemistry, 39, 2003–2209.
Kumar, D., & Bhalla, T. C. (2004). Indian Journal of Experimental Biology, 42, 515–521.
Miyaji, T., Otta, Y., & Shibata, T. (2005). Letters in Applied Microbiology, 41, 253–257.
Takami, H., Akiba, T., & Horikoshi, K. (1989). Applied Microbiology and Biotechnology, 30, 120–124.
Papagianni, M., & Sergelidis, D. (2014). Applied Biochemistry and Biotechnology, 172, 3926–3938.
Niyonzima, F. N., & More, S. (2014). Preparative Biochemistry and Biotechnology, 44(7), 738–759.
Ghareib, M., Fawzi, E. M., & Aldossary, N. A. (2014). Annals of Microbiology, 64(2), 859–867.
Raval, V. H., Pillai, S., Rawal, C. M., & Singh, S. P. (2014). Process Biochemistry, 49(6), 955–962.
Oberoi, R., Beg, Q. K., & Puri, S. (2001). World Journal of Microbiology and Biotechnology, 17, 493–497.
Saeki, K., Hitomi, J., Okuda, M., & Hatada, Y. (2002). Extremophiles, 6, 65–72.
Rao, M. B., Tanksale, A. M., Ghatge, M. S., & Deshpande, V. V. (1998). Microbiology and Molecular Biology Reviews, 62, 597–635.
Kulakova, L., Galkin, A., Nakayama, T., Nishino, T., & Esaki, N. (2003). Journal of Molecular Catalysis B-Enzyme, 22, 113–117.
Acknowledgments
This research work has been supported by National Natural Science Foundation of China (No. 41376175) and Qingdao Municipal Science and Technology Plan Project (14-2-4-11-jch).
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article

Cite this article
Hao, JH., Sun, M. Purification and Characterization of a Cold Alkaline Protease from a Psychrophilic Pseudomonas aeruginosa HY1215. Appl Biochem Biotechnol 175, 715–722 (2015). https://doi.org/10.1007/s12010-014-1315-2
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s12010-014-1315-2