
Abstract
Antioxidant properties and angiotensin-converting enzyme (ACE) inhibitory activities of protein hydrolysates from goby (Zosterisessor ophiocephalus) muscle, with different degrees of hydrolysis (DH) from 5 to 25 %, prepared by treatment with crude proteases extract from smooth hound intestines, were investigated. Goby protein hydrolysates (GPHs) are rich in Gly and Thr, which accounted for 14.1–15 % and 11.6–13.2 % of the total amino acids, respectively. The antioxidant activities of GPHs were investigated by using several in vitro assay systems. All GPHs exhibited significant metal chelating activity and DPPH free radical-scavenging activity, and inhibited linoleic acid peroxidation. For the ACE-inhibitory activity, as the DH increased, the activity of GPHs increased. The obtained results revealed that antioxidant and ACE-inhibitory activities of GPHs were influenced by the degree of hydrolysis. A medium degree of enzymatic hydrolysis was appropriate to obtain GPHs with good antioxidant activity, while small peptides were essential to obtain high ACE inhibitory activity.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
Several studies in the past few decades have reported that protein hydrolysates, in addition to their nutritional properties, exhibited biological activities, such as anticoagulant [1], antioxidant [2], antihypertensive [3, 4]), and cholesterol-lowering activity [5]. These functions are associated with bioactive peptides present in hydrolysates. These peptides, which are inactive within the sequence of the original protein, can be liberated by enzymatic hydrolysis, either during gastrointestinal digestion in the body, or during food processing, or by proteolysis processes using appropriate exogenous proteasess [6]. These peptides usually contain 3–20 amino acids, and their activity depends on their amino acid composition and sequence [7, 8]. Bioactive peptides can be used as ingredients in functional foods, nutraceuticals, or pharmaceuticals due to their therapeutic potential in the treatment or prevention of various diseases.
The enzymatic hydrolysis of whole protein molecules is the most commonly used method for producing bioactive peptides. Proteolytic enzymes from bacteria, fungus, plants, and digestive enzymes can be used for the proteolysis of protein substrates [9]. In some cases, multiple specific or nonspecific proteases can be used in order to enhance protein hydrolysis and obtain hydrolysate enriched with low molecular weight peptides of interest. In addition to the appropriate enzyme and the adequate protein substrate, proteolysis reaction conditions for the generation of protein hydrolysates from raw materials should be controlled to obtain potent and desired biopeptides. Degree of hydrolysis is an important factor affecting the bioactivities of protein hydrolysates. Hydrolysates produced, which had different hydrolysis degree, exhibited different range of bioactivities. Thus, hydrolysis degree should be controlled to obtain reproducible functional and biological properties and high nutritional value.
Lipid oxidation is of great concern to the food industry and consumers because it leads to the development of undesirable off-flavors, odors, dark colors, and potentially toxic reaction products [10]. Furthermore, cancer, coronary heart disease, and Alzheimer’s diseases are also reported to be caused in part by oxidation or free radical reactions in the body [11]. To prevent oxidative deterioration of foods and to provide protection against serious diseases, it is important to inhibit the oxidation of lipids and the formation of free radicals occurring in the foodstuff and living body. Antioxidants are used to preserve food products by retarding discoloration and deterioration caused by oxidation [12]. Synthetic antioxidants, such as butylated hydroxyanisole (BHA) and butylated hydroxytoluene (BHT), have been widely used in stabilization of foods [13]. However, use of these chemical compounds has begun to be restricted because of their induction of DNA damage and their toxicity [14]. Moreover, both BHT and BHA appear to be involved in tumor promotion [15]. Therefore, there is great interest in finding new and safe antioxidants from natural sources, especially peptides derived from hydrolyzed food proteins. Recently, a great deal of interest has been expressed regarding marine-derived bioactive peptides because of their numerous health benefits. Protein hydrolysates, from different muscle of species such as zebra blenny (Salaria basilisca) [16], and silver carp [17], have been found to possess antioxidant activity. Further, several antioxidant peptides have been isolated from various food proteins such as salmon byproduct [18] and collagen from scales of the croceine croacker (Pseodosciana crocea) [19].
On the other hand, elevated blood pressure or hypertension has been considered as one of the major risk factors for the development of cardiovascular diseases (CVD) that affects 30 % of the adult population in the world [20]. Angiotensin I-converting enzyme, a dipeptidyl carboxypeptidase, plays an important physiological role in the renin-angiotensin system (RAS), which regulates human blood pressure and fluid homeostasis. The enzyme catalyzes the conversion of angiotensin I [Asp-Arg-Val-Tyr-Ile-His-Pro-Phe-His-Leu] to a potent vasoconstrictor angiotensin II, an octapeptide [Asp-Arg-Val-Tyr-Ile-His-Pro-Phe] by removing the C-terminal dipeptide His-Leu [21]. In addition, ACE catalyzes the degradation of bradykinin, which is a known vasodilator [22]. Hence, since ACE plays an important role in the regulation of blood pressure, the inhibition of ACE is considered to be a useful approach in the treatment of hypertension. Currently, commonly used synthetic ACE inhibitors, such as captopril and lisinopril, have strong side effects in humans, such as cough, loss of taste sensation, and renal impairment [23]. In recent years, an increasing number of studies have been performed to investigate the hypotensive effects of food-derived protein hydrolysates [24, 25].
The goby (Z. ophiocephalus), is common in the Mediterranean Sea, Black Sea, and Sea of Azovthe. It reaches a maximum length of 25 cm and it is carnivorous; juveniles feed on small crustaceans, polychaets, and molluscs. It is relatively important in the fish catches of Tunisia, and is utilized for human consumption. In Tunisia, goby (Z. ophiocephalus) catches were about 130 tonnes in 2004 [26]. So far, no information regarding bioactive peptides from the goby has been documented.
The objective of this study was to evaluate the influence of the extent of enzymatic hydrolysis on antioxidative and ACE-inhibitory activities of goby protein hydrolysates (GPHs) obtained by treatment with smooth hound intestine crude proteases extract.
Materials and Methods
Reagents
Angiotensin I-converting enzyme (ACE) from rabbit lung, ACE synthetic substrate hippuril-L-histidyl-L-leucine (HHL), butylated hydroxyanisole (BHA), β-carotene, linoleic acid, ethylenediaminetetraacetic acid (EDTA), 1,1-diphenyl-2-picrylhydrazyl (DPPH), and L-ascorbic acid were purchased from Sigma Chemical Co. (St. Louis, MO, USA).
All other chemicals, namely potassium ferricyanide, trichloroacetic acid (TCA), ferric chloride, sodium hydroxide, ammonium thiocyanate, ferrous chloride, ferrozine, Tween 40, and other solvents, were of analytical grade. All solutions were freshly prepared in distilled water.
Materials
Goby (Z. ophiocephalus) was purchased from the fish market of Sfax city, Tunisia. The samples were packed in polyethylene bags, placed in ice with a sample/ice ratio of approximately 1:3 (w/w), and then transported to the research laboratory within 30 min. Muscle was separated, rinsed with cold distilled water to remove salts and other contaminants, and then stored in sealed plastic bags at −20 °C until it was used for protein hydrolysates production, less than 1 week later.
Smooth hound (Mustelus mustelus) was purchased from the local market at Sfax city, Tunisia. After the fish was washed with tap water, internal organs were separated into individual organs and only intestines were collected.
Preparation of Smooth Hound Crude Proteases Extract
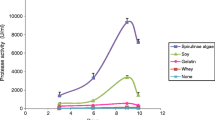
Intestines from the smooth hound (150 g) were thoroughly rinsed with cold distilled water, cut into pieces with a thickness of 1 to 1.5 cm, and then homogenized for 1 min with 300 ml of extraction buffer (10 mM Tris–HCl buffer, pH 8.0). The homogenate was centrifuged at 8,500×g for 15 min at 4 °C using refrigerated centrifuge. The pellet was discarded and the supernatant was collected. The supernatant was then fractionated with ammonium sulfate (80 % saturation). The pellet was recovered by centrifuging at 10,000×g for 15 min at 4 °C. The precipitate was suspended in 100 mM Tris–HCl buffer (pH8.0) and used as crude alkaline proteases extract. Alkaline proteases activity was determined according to the method of Kembhavi et al. [27] using casein as a substrate at pH 8.0 and 60 °C for 15 min. One unit of proteases activity was defined as the amount of enzyme required to liberate 1 μg of tyrosine per minute under the experimental conditions used. Values are the means of three independent experiments.
Production of Goby Muscle Protein Hydrolysates with Different DH
Goby muscle (500 g), in 1,000 ml distilled water, were first minced using a grinder (Moulinex Charlotte HV3, France), and then cooked at 90 °C for 20 min to inactivate endogenous enzymes. The cooked muscle sample was then homogenized in a Moulinex® blender for about 5 min. The pH of the mixture was adjusted to the optimum activity value (pH = 8.0). The protein solution was allowed to equilibrate for 30 min before hydrolysis was initiated. Control experiments were also performed without enzyme addition. After the equilibrium was reached, the hydrolysis reaction was started by the addition of the enzyme at a 3:1 (U/mg) enzyme/protein ratio. The reaction was conducted at pH 8.0, 50 °C for 250 min. During the reaction, the pH of the mixture was maintained at the desired value by continuous addition of 4 mol/L NaOH solution. The hydrolysates with varying degrees of hydrolysis, namely 5, 10, 15, 20, and 25 %, were obtained after incubation at 60 °C. After the required digestion time, the reaction was stopped by heating the solution for 20 min at 80 °C to inactivate the enzymes and then cooled down to room temperature. Protein hydrolysates were then centrifuged at 5,000×g for 20 min to separate insoluble and soluble fractions. Finally, the supernatant was freeze-dried using freeze-dryer (Bioblock Scientific Christ ALPHA 1–2, IllKrich-Cedex, France) and stored at −20 °C until required for characterization, and functional and biological properties analyses.
The degree of hydrolysis, defined as the percent ratio of the number of peptide bonds cleaved (h) to the total number of peptide bonds in the protein substrate (h tot ), was determined by the pH-stat method as described by Adler-Nissen [28]:
Chemical Analysis
Moisture and ash contents of GPHs were determined according to the AOAC methods 930.15 and 942.05, respectively [29]. The protein content was determined by estimating its total nitrogen content by Kjeldahl method according to the AOAC method number 984.13 [29]. A factor of 6.25 was used to convert the nitrogen value to protein. Lipids were determined gravimetrically after Soxhlet extraction of dried samples with hexane. All measurements were performed in triplicate. The protein, ash, and fat contents were expressed on a dry weight basis.
Amino Acid Analysis
Goby protein hydrolysate (GPH) sample was hydrolyzed with 0.5 mL of 6 mol/L HCl at 110 °C for 24 h on a heating block, and then filtered through a 0.45-μm membrane filter prior to analysis. About 10 μl of the treated sample was derivatized using 6-aminoquinolyl-N-hydroxysuccinimidyl carbamate Waters AccQ · Fluor Reagent Kit (according to Waters AccQ · Tag Chemistry Package Instruction Manual).
The HPLC analyses were performed with a Waters 2996 Separation Module equipped with a Waters 2,475 multi-wavelength fluorescence detector, and amino acids were separated on a Waters AccQ · Tag amino acid analyzing Column (Nova-Pak C18, 150 × 3.9 mm). The amount of amino acids was calculated, based on the peak area in comparison with that of standard. The amino acid content was expressed as a percentage of total amino acids in the sample. All analyses were performed in duplicate.
Determination of Antioxidant Activities
DPPH Free Radical-scavenging Activity
The DPPH free radical-scavenging activity of GPHs was determined as described by Bersuder et al. [30]. A volume of 500 μl of each sample at a concentration of 5 mg/ml was added to 375 μl of 99 % ethanol and 125 μl of DPPH solution (0.02 % in ethanol) as free radical source. After vortexing, the reaction mixture was incubated for 60 min in the dark at room temperature. The absorbance of the resulting solution was measured at 517 nm. In its radical form, DPPH has an absorption band at 517 nm which disappears upon reduction by an antiradical compound. Lower absorbance of the reaction mixture indicated higher DPPH free radical-scavenging activity. BHA was used as positive control. DPPH radical-scavenging activity was calculated as follows:
where A blank is the absorbance of the reaction containing all reagents except the sample and A sample is the absorbance of GPHs (with the DPPH solution). The experiment was carried out in triplicate and the results are mean values.
Ferric-reducing Activity
The ability of GPHs to reduce iron III was determined by the method of Yildirim et al. [31]. A sample solution (0.5 ml; 0.5 mg/ml) of each hydrolysate were mixed with 1.25 ml of 0.2 M phosphate buffer (pH 6.6) and 1.25 ml of 1 % potassium ferricyanide solution. The mixtures were incubated for 30 min at 50 °C. After incubation, 1.25 ml of 10 % TCA was added and the reaction mixtures were centrifuged for 10 min at 3,000×g. An aliquot of the supernatant (1.25 ml) from each sample mixture was mixed with 1.25 ml of distilled water and 0.25 ml of 0.1 % ferric chloride solution. After a 10-min reaction time, the absorbance of the resulting solutions was measured at 700 nm. Higher absorbance of the reaction mixture indicated higher reducing power. The control was conducted in the same manner, except that distilled water was used instead of sample. Values presented are the mean of triplicate analyses.
Antioxidant Assay Using the ß-Carotene Bleaching Method
The ability of GPHs to prevent bleaching of β-carotene was assessed as described by Koleva et al. [32]. A stock solution of β-carotene/linoleic acid mixture was prepared by dissolving 0.5 mg of β-carotene, 25 μl of linoleic acid, and 200 μl of Tween 40 in 1 ml of chloroform. The chloroform was completely evaporated under vacuum in a rotatory evaporator at 40 °C, then 100 ml of bi-distilled water were added, and the resulting mixture was vigorously stirred. The emulsion obtained was freshly prepared before each experiment. Aliquots (2.5 ml) of the β-carotene/linoleic acid emulsion were transferred to test tubes containing 0.5 ml of each protein hydrolysate (5 mg/ml). Following incubation for 2 h at 50 °C, the absorbance of each sample was measured at 470 nm. BHA was used as positive standard. A blank consisted of 0.5 ml of distilled water instead of sample.
The antioxidant activity of the hydrolysates was evaluated in terms of bleaching of β-carotene using the following formula:
where A0 and A’0 are the absorbances of the sample and the blank, respectively, measured at time zero, and At and A’t are the absorbances of the sample and the control, respectively, measured after incubation for 2 h. The same procedure was repeated with BHA as positive control. Values presented are the mean of triplicate analyses.
Metal Chelating Activity
The chelating activity of samples towards ferrous ion (Fe2+) was determined according to the method of Decker and Welch [33]. One milliliter of each sample solution at different concentrations (1, 2, 5 mg/ml) was mixed with 3.7 ml of distilled water. The mixture was then reacted with 0.1 ml of 2 mM FeCl2 4H20 and 0.2 ml of 5 mM 3-(2-pyridyl)-5,6-bis(4-phenyl-sulfonic acid)-1,2,4-triazine (ferrozine) for 20 min at room temperature. The absorbance was then read at 562 nm. The blank was prepared in the same manner except that distilled water was used instead of the sample. EDTA was used as reference. The chelating activity (%) was calculated as follows:
Determination of ACE Inhibition Activity
ACE-inhibitory activity was assayed as reported by Nakamura et al. [34]. A volume of 80 μl of GPH (2 mg/ml) sample was added to 200 μl of 5 mmol/l HHL, and then preincubated for 3 min at 37 °C. GPHs and HHL were prepared in 100 mM borate buffer (pH 8.3) containing 300 mM NaCl. The reactions were then initiated by adding 20 μl of 0.1 U/ml ACE from rabbit lung, prepared in the same buffer. After incubation for 30 min at 37 °C, the enzymatic reactions were stopped by adding 250 μl of 0.05 M HCl. The hippuric acid (HA) released was extracted with ethyl acetate (1.7 ml), and after removal of ethyl acetate by heat evaporation (10 min at 95 °C), HA was redissolved in 1 ml of distilled water and the absorbance of the extract at 228 nm was determined using a UV–visible spectrophotometer (Cecil CE 2021, Lab Equip Instruments Ltd). ACE-inhibitory activity was calculated using the equation
where A is the absorbance of HA generated in the presence of ACE inhibitor component, B the absorbance of HA generated without ACE inhibitors, and C is the absorbance of HA generated without ACE (corresponding to HHL autolysis in the course of enzymatic assay). The IC50 value was defined as the concentration of hydrolysate (mg/ml) required to reduce 50 % of ACE activity under the above condition.
Statistical Analyses
Statistical analyses were performed with Stratgraphics ver. 5.1, professional edition (Manugistics Corp., USA) using ANOVA analysis. Differences were considered significant at p < 0.05. All tests were carried out in triplicate.
Results and Discussion
Production and Characterization of GPHs with Different DH
Enzymatic proteolysis is a valuable bioprocess to enhance biological activities and functional properties of the original proteins, and some peptides generated from the hydrolysis possess stronger activity than others [35]. The biological activities of protein hydrolysates are closely related to different extrinsic parameters which are usually the DH, the specificity of the enzyme, the physicochemical conditions, etc. [36] Hence, the DH should be controlled to obtain reproducible and optimum peptide size distribution when the enzymatic hydrolysis is applied to produce potent bioactive peptides.
Hydrolysates with increasing DH (from 5 to 25 %) were prepared in order to study the effect of the extent of enzymatic hydrolysis on the evolution of some biological activities. The hydrolysis curve of goby proteins after 250 min of incubation is shown in Fig. 1. The curve showed a high rate of hydrolysis during the initial 30 min, and thereafter the rate subsequently decreased. The shape of the hydrolysis curve is similar to those previously published for hydrolysates from the yellow stripe trevally (Selaroides leptolepis) [2], goby (Z. ophiocephalus) [1], zebra blenny (S. basilisca) [16], and sardinelle (Sardinella aurita) [5].

Hydrolysis curve of goby proteins treated with crude proteases extract from smooth hound viscera
The proximate composition of undigested goby proteins (UGP) and their hydrolysates are shown in Table 1. The raw material (UGP) contained high amount of proteins and lipids up to 72 and 7.4 %, respectively. The protein contents of GPHs with different DH were determined to be 69.53–71.9 %. The GPHs had lower levels of lipid which decrease with the increase of DH, which might significantly contribute to stability of GPHs. Further, protein and lipids contents decreased with the increase of DH, while the ash content increased with the increase of degree of hydrolysis from 13.3 to 17.6 %. The ash content of all hydrolysates was higher than that of UGP (9.45 %). These results are similar to those of other published studies on fish protein hydrolysates [37, 38]. The high ash content of protein hydrolysates may be attributed to the addition of alkali required to control the pH during the hydrolytic process [39, 40].
The amino acids compositions of the freeze-dried GPHs, expressed as residues per 100 residues, are presented in Table 2. The results revealed the presence of essential and non essential amino acids. The percentages of amino acids between GPHs are slightly different, which may be attributed to the differences in the incubation time of protein digestion. Glycine and threonine were the most abundant amino acids, which accounted for 14.1–15 %, 11.6–13.2 % of the total amino acids, respectively.
Under the conditions of the acid hydrolysis, tryptophane was destroyed. On the other hand, GPHs have a relatively high content of Val, Met, Lys, Ile, Leu, Phe, and Thr. However, cysteine is in low amount. Several amino acids, such as Met, His, Tyr, Lys, and Trp contributed significantly to the antioxidant activity of the hydrolysates [41]. Antioxidant activity of histidine or histidine-containing peptides may be attributed to the chelating and lipid radical-trapping ability of the imidazole ring, whereas the tyrosine residue in the peptide may act as a potent hydrogen donor [42]). In addition, GPHs have also relatively high content of Pro, Phe, and Tyr. Indeed, it was reported that these amino acids as well as hydrophobic amino acids at the C-terminal of peptides are very important to the ACE-inhibitory activity [43]).
From the results, GPHs had a high nutritional value based on their amino acids compositions, and therefore could possibly be a dietary protein supplement to poorly balanced dietary proteins.
Effect of DH on Antioxidant Activity
The overall antioxidant action of protein hydrolysates is most likely attributed to the cooperative effects of several mechanisms, including, metal ion chelating, free radical scavenging, and singlet electron transferring (i.e., reducing capacity) [44].
DPPH Radical-scavenging Activity
Figure 2 shows the results of DPPH radical-scavenging activity of goby protein hydrolysates, with different DH values at final assay concentration of 5 mg/ml, compared to the undigested goby proteins (UGP). UGP and All GPHs were able to scavenge DPPH radicals. Further, the DPPH radical-scavenging activity was improved by the providing of shorter chain peptides with higher bioactivity by the proteolysis of goby muscle proteins. Moreover, the antioxidant activity of GPHs increased with increasing DH and reached its maximum with hydrolysate having a DH of 15 % (64 ± 5 %). However, further increase in the DH from 15 to 25 % reduced the activity. Similar results have been reported for loach protein hydrolysate [45] and cuttlefish muscle proteins hydrolyzed by Bacillus mojavensis A21 proteases [38]. In the case of cuttlefish protein hydrolysates, the activity increased with the increase of DH and was maximal with hydrolysate of 16 % and further digestion resulted in a slight decrease in the activity. Thus, the higher DH can have negative effect on the bioactivity by the formation of very shorter peptides possessing less hydrophobicity. Those peptides had lesser efficacy in scavenging DPPH activity [41]. However, in another study, Klompong et al. [2] have shown a decrease in the DPPH radical-scavenging activity of the yellow stripe trevally protein hydrolysate, prepared with Flavourzyme and Alcalase, with the increase in DH.

DPPH radical-scavenging activities of GPHs. BHA was used as positive control
DPPH is a stable free radical and can be scavenged with a proton-donating substrate, such as an antioxidant [46]. Therefore, the results show that protein hydrolysates from goby more likely contained some bioactive peptides, which are proton donor, and could react with free radicals to convert them to more stable products, thereby terminating the radical chain reaction [44]. The differences in the radical-scavenging ability between hydrolysates may be attributed to the differences in chain length and amino acid sequence of peptides which contributed to varying degree of scavenging DPPH radicals.
Reducing Power
The reducing power assay is often used to evaluate the ability of an antioxidant to donate an electron to the free radical [47]. Different studies have reported that there is a direct correlation between antioxidant activities and reducing powers of some bioactive compounds [48]. Samples with higher reducing power have better abilities to donate electron. In this study, the ability of GPHs to reduce Fe3+ to Fe2+ was determined. As reported in Fig. 3, all GPHs and UGP showed reducing power activities. Further, the reducing power activity increased with the increase of DH and was maximal with hydrolysate of 10 %. Nevertheless, further increase in the DH from 10 to 25 % resulted in a decrease in the activity. However, all protein hydrolysates showed lower reducing power activities than BHA at the same concentrations.

Reducing power of GPHs. BHA was used as positive control
The above results also indicated that DH greatly influenced the antioxidant activity.
Antioxidant Activity Measured by the β-carotene Bleaching Method
The antioxidant assay using the discoloration of β-carotene is widely used to measure the antioxidant activity of bioactive compounds because β-carotene is extremely susceptible to free radical mediated oxidation of linoleic acid [49]. In this test, β-carotene undergoes rapid discoloration in the absence of antioxidant, which results in a reduction in absorbance of the test solution with reaction time. The presence of antioxidant hinders the extent of bleaching by neutralizing the linoleic free radical formed. In Fig. 4, the antioxidant activities of GPHs with the different DH are compared with that of UGP. The β-carotene bleaching decreased with addition of GPHs and UGP. The GPH with a DH of 15 % exhibited a significantly (p < 0.05) higher ability to prevent the oxidation of linoleic acid. However, further increases in the DH from 20 to 25 % decreased the antioxidant activity of GPH.

Determination of antioxidant activity using the β-carotene bleaching method of GPHs
These results demonstrated that GPHs have strong effects against the discoloration of β–carotene.
Ferrous Ion-chelating Activity
The chelation of Fe2+ was also used to determine the ability of protein hydrolysates in metal chelating activity. Ferrozine can quantitatively form complexes with Fe2+ ion. In the presence of chelating agents, the complex formation is disrupted, resulting in a decrease in the red color of the complex [50]. Indeed, some proteins and peptides can chelate metal ions like Fe2+ due to the presence of carboxyl and amino groups in the side chains of acidic and basic amino acids [51].
The ferrous chelating activities of hydrolysates, from the muscle of goby with different DH, at different concentrations are reported in Fig. 5. Metal chelating activity of all hydrolysates and of the UGP increased with increasing concentrations of hydrolysates from 1 to 5 mg/ml. The chelating effect of the UGP is lower than the GPHs at different concentrations. Further, chelating activity against Fe2+ of all GPHs increased with increasing DH and reached a maximum chelating effect of 84.77 ± 1.67 % with a DH of 20 % at a concentration of 5 mg/ml and further increase in DH resulted in a decrease in metal chelating activity. The results indicate that the metal ion-binding capacity of protein hydrolysates was enhanced by enzymatic hydrolysis. The result was in accordance with Nalinanon et al. [52] who reported that chelating activity of Fe2+ of protein hydrolysates from the muscle of ornate threadfin bream treated with pepsin from skipjack tuna increased with increasing DH, and hydrolysate with 30 % DH exhibited the highest Fe2+ chelating activity (58.1 %).

Metal chelating activity of GPHs. EDTA was used as positive control
The results so obtained indicate that hydrolysates from the muscle of goby can exhibit, to a various extent, antioxidant ability by capturing ferrous ion or other ions. Therefore, chelation of transition metal ions by antioxidants peptides in hydrolysates could retard the oxidation reaction [53].
The results so obtained showed that UGP and GPHs with different DH exhibited to a variable extent antioxidant activities against various antioxidant systems in vitro. Further, the results revealed that hydrolysates with medium molecular weight peptides are appropriate to obtain hydrolysates with excellent antioxidant activities, since higher antioxidant activities were observed with hydrolysates having a DH of 10 or 15 %, and further digestion resulted in decrease in activities.
ACE-Inhibitory Activities of GPHs
The ACE-inhibitory activity of GPHs with different DH was investigated and reported in Fig. 6. Inhibition activity was correlated with the degree of hydrolysis (p < 0.05), since the activity increased with the increase of DH, reached a value of 82.9 % with a DH of 25 %, while no significant activity was detected with undigested proteins (UGP).

ACE-inhibitory activities of GPHs with different DH
The results so obtained demonstrate that ACE-inhibitory peptides are encrypted within goby proteins and could be released by proteolysis. The differences in ACE-inhibitory activities of the hydrolysates might be due to the different size of ACE-inhibitory peptides obtained in protein hydrolysates. Depending on the DH, protein hydrolysates contained different sizes of peptides and free amino acids. Indeed, hydrolysates with low DH should contain high and medium molecular weight peptides, while hydrolysates with high DH contained medium and low molecular weight peptides. The obtained results revealed that the activity was associated with very low molecular weight peptides. Therefore, a high degree of hydrolysis was essential to obtain highly potent ACE-inhibitory peptides from the muscle of goby. Furthermore, GPHs with a high DH showed high solubility (data not shown), which is useful in its application in the food industry as a functional ingredient.
These findings further support the reports of Cai et al. [54] and Barbana and Boye, [55], who showed that hydrolysates with high DH had higher ACE-inhibitory activity.
According to the above results, the degree of hydrolysis greatly influenced the molecular weight and amino acid residues composition of peptides in protein hydrolysates and further influenced the ACE-inhibitory activity.
Conclusion
The results of this study indicate that proteolysis is a good alternative to obtain bioactive protein hydrolysates. Indeed, GPHs with different DH exhibited varying degrees of antioxidant activities evaluated by using various antioxidant assays, and medium DH significantly improved the antioxidant activity. These differences may be due to the differences in the amino acid compositions and sequences as well as the length of peptides in hydrolysates. Additionally, goby protein hydrolysates exhibited notable ACE-inhibitory activity, and the activity was associated with very short-chain peptides. These results have demonstrated that DH should be controlled in the production of protein hydrolysates or peptides with high and reproducible desired biological activities.
The obtained results demonstrate that hydrolysates from the muscle of goby can be used as a promising source of bioactive peptides.
References
Nasri, R., Ben Amor, I., Bougatef, A., Nedjar-Arroume, N., Dhulster, P., Gargouri, J., Karra-Châabouni, M., & Nasri, M. (2012). Food Chemistry, 133, 835–841.
Klompong, V., Benjakul, S., Kantachote, D., & Shahidi, F. (2007). Food Chemistry, 102, 1317–1327.
Nasri, R., Chataigne, G., Bougatef, A., Karra-Châabouni, M., Dhulster, P., Nasri, M., & Nedjar-Arroume, N. (2013). Journal of Proteomics, 91, 444–452.
He, H. L., Liu, D., & Ma, C. B. (2013). Applied Biochemistry and Biotechnology, 169, 738–749.
Ben Khaled, H., Ghlissi, Z., Chtourou, Y., Hakim, A., Ktari, N., Makni-Ayedi, F., Barkia, A., Sahnoun, Z., & Nasri, M. (2012). Food Research International, 45, 60–68.
Clare, D. A., & Swaisgood, H. E. (2000). Journal of Dairy Science, 83, 1187–1195.
Korhonen, H., & Pihlanto, A. (2006). International Dairy Journal, 16, 945–960.
Agyei, D., & Danquah, M. K. (2011). Biotechnology Advance, 29, 272–277.
Simpson, B. K., Nayeri, G., Yaylayan, V., & Ashie, I. N. A. (1998). Food Chemistry, 61, 131–138.
Lin, C. C., & Liang, J. H. Journal of Food Science, 67, 530–533.
Diaz, M. N., Frei, B., Vita, J. A., & Keaney, J. F. (1997). England Journal of Medicine, 337, 408–416.
Halliwell, B., Murcia, M. A., Chirico, S., & Aruoma, O. I. (1995). Critical Reviews in Food Science and Nutrition, 35, 7–20.
Löliger, J. (1991). (Eds.), by Aruoma OI and Halliwell B, London, pp. 121–150.
Ito, N., Hirose, M., Fukushima, S., Tsuda, H., Shirai, T., & Tatematsu, M. (1986). Food Chemistry and Toxicology, 24, 1071–1082.
Botterweck, A. A. M., Verhagen, H., Goldbohm, R. A., Kleinjans, J., & Van den Brandt, P. A. (2000). Food Chemistry and Toxicology, 38, 599–605.
Ktari, N., Jridi, M., Bkhairia, I., Sayari, N., Ben Salah, R., Ben Salah, R., & Nasri, M. (2012). Food Research International, 49, 747–756.
Dong, S. Y., Zhao, Y. H., Xu, D. X., Liu, Z. Y., & Zeng, M. Y. (2013). Journal of Aquatic Food Product Technology, 22, 573–583.
Ahna, C. B., Kim, J. G., & Je, J. Y. (2014). Food Chemistry, 147, 78–83.
Wang, B., Wang, Y. M., Chi, C. F., Luo, H. Y., Deng, S. G., & Ma, J. Y. (2013). Marine Drugs, 11, 4641–4661.
Harris, T., Cook, E. F., Kannel, W., Schatzkin, A., & Goldman, L. (1985). Hypertension, 7, 118–124.
Lavoie, J. L., & Sigmund, C. D. (2003). Endocrinology, 144, 2179–2183.
Ondetti, M. A., & Cusham, D. W. (1977). Science, 96, 441–444.
Acharya, K. R., Sturrock, E. D., Riordan, J. F., & Ehlers, M. R. W. (2003). Nature Reviews Drug Discovery, 2, 891–902.
Megias, C., Pedroche, J., Yust, M. M., Alaiz, M., Giron-Calle, J., Millan, F., & Vioque, J. (2009). LWT-Food Science and Technology, 42, 228–232.
Escudero, E., Mora, L., Fraser, P. D., Aristory, M. C., Arihara, K., & Toldra, F. (2013). Journal of Proteomics, 78, 499–507.
F.A.O. Food and Agriculture Organisation. Fishery Statistics, Rome, Italy, 2004.
Kembhavi, A. A., Kulkarni, A., & Pant, A. (1993). Applied Biochemistry and Biotechnology, 38, 83–92.
Adler-Nissen, J. (1986). Elsevier, Copenhagen
AOAC. Official methods of analysis. Arlington, VA Secs. 930.15–942.05, 2000.
Bersuder, P., Hole, M., & Smith, G. (1998). Journal of the American Oil Chemists’ Socity, 75, 181–187.
Yildirim, A., Mavi, A., & Kara, A. A. (2001). Journal of Agriculture and Food Chemistry, 49, 4083–4089.
Koleva, I. I., van Beek, T. A., Linssen, J. P. H., de Groot, A., & Evstatieva, L. N. (2002). Phytochemistry Analysis, 13, 8–17.
Decker, E. A., Welch, B. Journal of Agriculture and Food Chemistry, 38, 674–677
Nakamura, Y., Yamamoto, N., Sakai, K., Okubo, A., Yamazaki, S., & Takano, T. (1995). Journal of Dairy Science, 8, 777–783.
Chen, H. N., Muramoto, K., & Yamauchi, F. (1995). Journal of Agriculture and Food Chemistry, 1995(43), 574–578.
Van Der Ven, C., Gruppen, H., de Bont, D. B. A., & Voragen, A. G. (2002). International Dairy Journal, 12, 813–820.
Gbogouri, G. A., Linder, M., Fanni, J., & Parmentier, M. (2004). Journal of Food Science, 69, 615–622.
Hmidet, N., Balti, R., Nasri, R., Sila, A., Bougatef, A., & Nasri, M. (2011). Food Research International, 44, 2703–2711.
Liceaga-Gesualdo, A. M., & Li-Chan, E. C. Y. (1999). Journal of Food Science, 64, 1000–1004.
Kristinsson, H. G., & Rasco, B. A. (2000). Journal of Food Biochemistry, 24, 177–87.
Chen, H. M., Muramoto, K., Yamauchi, F., & Nokihara, K. (1996). Journal of Agriculture and Food Chemistry, 44, 2619–2623.
Je, Y. J., Park, P. J., & Kim, S. K. (2005). Food Research International, 38, 45–50.
Gobbetti, M., Ferranti, P., Smacchi, E., Goffredi, F., & Addeo, F. (2000). Applied Environmental Microbiology, 9, 3898–3904.
Chen, H. M., Muramoto, K., Yamauchi, F., Fujimoto, K., & Nokihara, K. (1998). Journal of Agriculture and Food Chemistry, 46, 49–53.
You, L., Zhao, M., Cui, C., Zhao, H., & Yang, B. (2009). Innovative Food Science and Emerging Technology, 10, 235–240.
Blois, M. S. (1958). Antioxidant determinations by the use of a stable free radical. Nature, 181(4617), 1199–1200. doi:10.1038/1811199a0#blank.
Khantaphant, S., & Benjakul, S. (2008). Comparative Biochemistry and Physiology, 151, 410–419.
Duh, P. D., Du, P. C., & Yen, G. C. (1999). Food Chemistry and Toxichology, 37, 1055–1061.
Kumazawa, S., Taniguchi, M., Suzuki, Y., Shimura, M., Kwon, M. S., & Nakayama, T. (2002). Journal of Agriculture and Food Chemistry, 50, 373–377.
Thiansilakul, Y., Benjakul, S., & Shahidi, F. (2007). Food Chemistry, 10, 1385–1394.
Saiga, A., Tanabe, S., & Nishimura, T. (2003). Journal of Agriculture and Food Chemistry, 51, 3661–3667.
Nalinanon, S., Benjakul, S., Kishimura, H., & Shahidi, F. (2011). Food Chemistry, 124, 1354–1362.
Sherwin, E. R. (1990). Food additives (pp. 139–193). New York: Marcel Dekker.
Cai, L., Xiao, L., Liu, C., & Ying, T. (2013). Food and Bioprocess Technology, 6, 2109–2117.
Barbana, C., & Boye, J. I. (2010). Food Research International, 43, 1642–1649.
Acknowledgments
This work was funded by the Ministry of Higher Education and Scientific Research, Tunisia.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article

Cite this article
Nasri, R., Jridi, M., Lassoued, I. et al. The Influence of the Extent of Enzymatic Hydrolysis on Antioxidative Properties and ACE-Inhibitory Activities of Protein Hydrolysates from Goby (Zosterisessor ophiocephalus) Muscle. Appl Biochem Biotechnol 173, 1121–1134 (2014). https://doi.org/10.1007/s12010-014-0905-3
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s12010-014-0905-3