
Abstract
Background
In vivo studies have suggested Caucasians achieve lower average knee flexion than non-Western populations. Some previous studies have also suggested gender may influence condylar AP translation and axial rotation, while others report an absence of such an influence.
Questions/purposes
We determined whether different ethnic and gender groups residing in the United States had different knee translations and rotations.
Methods
Three-dimensional knee rotations and translations were determined for 72 healthy subjects (24 Caucasian men, 24 Caucasian women, 13 Japanese men, 11 Japanese women) from full extension to maximum flexion using a fluoroscopic technique, under in vivo, weightbearing conditions.
Results
Although we observed substantial variability in all groups, small differences between groups were found, especially in deep flexion. Japanese women and men and Caucasian women achieved higher maximum flexion (153°, 151°, and 152°, respectively) than Caucasian men (146°). External rotation was higher for these three groups than for Caucasian men. The medial condyle remained more anterior for Caucasian women and all Japanese subjects than for Caucasian men, possibly leading to greater axial rotation and flexion, observed for these three groups.
Conclusion
We identified small differences in maximum flexion between genders and ethnic groups. While no differences were identified in the lateral condyle translation, the medial condyle remained more stationary and more anterior for the groups that achieved highest (and similar) maximum flexion. Therefore, it may be important for future implant designs to incorporate these characteristics, such that only the lateral condyle experiences greater posterior femoral rollback, while the medial condyle remains more stationary throughout flexion.
Similar content being viewed by others

Explore related subjects
Discover the latest articles, news and stories from top researchers in related subjects.Avoid common mistakes on your manuscript.
Introduction
Current TKA reduces pain, restores mobility, and provides satisfactory longevity [44, 45]. However, even patients having high Knee Society Scores often experience limitations in their daily activities, especially those requiring deep knee flexion [38]. Surgery is sometimes refused in non-Western cultures due to the anticipated limited postoperative ROM [47]. Therefore, improvements to facilitate deep flexion activities are required in the present TKA designs. To make design changes, it is important to understand which factors influence deep flexion capabilities. Since women seem to be more flexible than men [2, 5] and Asian populations can achieve more flexion than Caucasians (156.9° [24], 157.3° [22], 159.6° [1], 160° [16], 162°[37] or even 165° [17] for Asian populations and 143.8° [40], 145° [16] for Caucasian), these two factors, gender and ethnicity, might reveal certain factors leading to higher knee flexion.
Physiologic and anatomic characteristics can vary considerably between individuals. However, several common features can be classified with respect to gender. One study suggests women have different motion patterns from men while performing the same athletic tasks. During running or cutting maneuvers, women reveal 8° less flexion, but 11° more valgus position then men [35]. Women reportedly have higher ACL laxity than men (6.4 mm and 4.9 mm, respectively [27]), possibly related to anatomic variations or hormonal differences [23, 39, 43]. During closed kinetic chain knee extension, women experience 23.6% less knee joint surface rolling than men [26]. During single-legged squatting exercise, women demonstrate 6.7° more ankle dorsiflexion and 2.7° more pronation, 9.1° more hip flexion, 3.2° adduction, and 6.6° of external rotation than their male counterparts [51]. Furthermore, women cannot develop as much torsional or shear stiffness as men [48, 49], despite experiencing greater quadriceps activation than men [9, 10, 26, 35, 51]. However, a radiographic study focusing on more detailed analysis of the tibiofemoral joint suggested (but could not prove due to limited cohorts) that tibiofemoral kinematics is independent of gender [30].
Research data originating from different regions of the world suggest ROM may also be a function of ethnicity or a lifestyle. Caucasians can flex to about 140°, while in societies where daily activities require full kneeling or squatting as in Japan, India, or the Middle East, subjects achieve up to 165°, providing an energy-saving flexed posture [16, 17]. Saudi Arabian men flex their knees, on average, to 159.6° [1], 15° more than Scandinavians [40]. Arabs maintain the flexion of 157.3° in their prayer position [22]. However, the differences in the methods employed in these studies make it difficult to compare and explain the higher ROM observed for Asian populations. Previous three dimensional, in vivo kinematic studies have routinely reported findings from population sizes limited to 10 to 20 subjects [4, 11, 14, 22, 30, 34, 37], but none of these studies involved large sample sizes of non-implanted knees, especially taking into account gender and ethnicity. Studies that did involve larger sample sizes most often utilized non weight-bearing conditions, two-dimensional or in vitro assessments.
Therefore, we asked whether gender and ethnicity influenced (1) maximum flexion; (2) femoral axial rotation and abduction/adduction (valgus/varus rotations); (3) AP translations of the tibiofemoral joint; and (4) type of motion experienced by each femoral condyle.
Patients and Methods
In vivo knee kinematics was determined for 72 subjects (24 Caucasian men, 24 Caucasian women, 13 Japanese men, and 11 Japanese women) (Table 1). All Caucasian subjects were of European descent and reported they had grown up in the United States, and Japanese subjects were included only if they reported they grew up in Japan and were of Japanese descent. All Japanese subjects in this study grew up in their natural environment and moved to the United States as adults. Therefore, their bone morphology was established before they moved and participated in this study, ensuring their outcomes will be representative of their ethnicity. All subjects had healthy, well-functioning knees with no ligament instability, laxity, or past knee injury in the examined knee. The study protocol was approved by the Internal Review Board at the University of Tennessee and by the Research Subjects Review Board at the University of Rochester. Informed consent was obtained from each subject before participating in the study.
While under fluoroscopic surveillance, each subject was asked to perform a deep knee bend followed by a forward lean of the examined knee so that maximum flexion could be achieved (Fig. 1). Participants did not use any supporting devices. The activity was analyzed from full extension to maximum flexion in 30° increments. The computer-aided design (CAD) models of the tibia, fibula, and femur bones were recreated in a segmentation process, based on MR images obtained for each participant. The MR images were acquired with a 3D gradient echo sequence in the sagittal plane (TE: 17 ms; TR: 45 ms; NEX: 1; flip angle: 30; imaging time: 7 minutes). The field of view was 140 mm, with a 512 × 512 matrix resulting in an in-plane image resolution of 0.27 mm and 1.2-mm-thick slices. Segmentation was performed by manually digitizing the bone surfaces using ScanIP software (Simpleware, Exeter, Devon, UK). The resulting data were used to create full 3D polygonal surface models for the distal part of the femur and the proximal part of the tibia and fibula.

To achieve maximum flexion, each participant performed deep knee bend activity followed by a forward lean.
The 3D kinematics was recovered by overlaying the 3D CAD bone models onto their respective silhouettes in fluoroscopic images (Fig. 2) using a previously described 3D-to-2D registration technique [12, 13, 25, 33]. An error analysis for this process was previously performed for TKA components, documenting a translational error of less than 0.5 mm and a rotational error of less than 0.5° [33].

During the 3D-to-2D registration process, the bone CAD models were fitted into their respective silhouettes visible in the fluoroscopic images. Original fluoroscopic video (top row) overlaid CAD models onto their respective silhouettes (middle row) and 3D view of the recreated relative kinematics (bottom row) for full extension and 30°, 60°, 90°, 120°, 150°, and maximum flexion (from left to right).
Unlike for metal objects, the fluoroscopic images of bones contain visible internal details, rather than just a black silhouette. This allows prediction of the intensities inside the object silhouette, which can lead to even more accurate prediction and registration of 3D bone models to the fluoroscopic images [33].
Once the 3D kinematics was recreated, the flexion, axial rotation, and abduction/adduction (valgus/varus rotation) were determined using a joint coordinate system [19]. Next, the medial and lateral tibiofemoral contact locations were identified based on the distance between the surface of the tibial plateau and the respective femoral condyles (Fig. 3) (Video 1 [supplemental materials are available with the online version of CORR]). The AP positions of the contact points on the tibial plateau were measured in the tibial (local) coordinate system.

Tibiofemoral contact point location was determined based on the relative orientation and position of the femur and tibia bones (see also Video 1 [supplemental materials are available with the online version of CORR]).
Although the shape of the femoral condyles is complex and nonuniform, its sagittal curvature can be divided into four main facets [36] out of which the extension and flexion facets cover almost the entire functional arc of the distal femoral condyles (Fig. 4). The centers of these facets were determined for medial and lateral condyles. By tracking these centers, the AP movement of femoral condyles could be adequately studied (Video 2 [supplemental materials are available with the online version of CORR]) [29].

(A) Although the curvatures of the distal femoral condyles are complex, they can be approximated by (B) fitting circles to the extension (R) and flexion (r) facets [28]. The centers of these two circles were tracked to help to better understand the motion of each condyle (see also Video 2 [supplemental materials are available with the online version of CORR]).
Based on the femoral rotations, translations, and curvature of the condyles, the amount of arc length rotation was calculated for each condyle and compared with the amount of pure condylar translations. The proportions of these two quantities indicated the type of motion present during the analyzed activity. Knee motion can be described using five unambiguous terms; roll, spin, combined roll-spin, concordant translation, and countertranslation [7] (Fig. 5). The rolling motion occurs when the body rolls on the surface without slipping (amount of arc length is equal to the distance traveled), spin occurs when the body rotates without translating, combined roll-spin is present when the body translates less than the arc length rotation, while the opposite case is called concordant translation. The countertranslation occurs whenever the body turns in the opposite direction to its translation. Theoretically, there might be another type of motion: sliding, when a body translates without rotating (Fig. 5) (Video 3 [supplemental materials are available with the online version of CORR]).

The motion of the knee condyles may be compared to a translating and rotating disk. Depending on the amount of translation and rotation, there are six types of motion: (A) sliding, (B) spinning, (C) rolling, (D) rolling and spinning, (E) concordant translation, and (F) countertranslation (see also Video 3 [supplemental materials are available with the online version of CORR]).
Since there were differences in height and weight (Table 1) among the individuals, it was necessary to normalize the results so that proper intersubject comparisons could be made. Therefore, the AP translations of the tibiofemoral contact point and of the femoral condyles were normalized with respect to the tibial plateau length (measured from the segmented CAD models of the tibia bone). The tibial plateau length and width were smaller for women than for men in both Caucasian and Japanese groups (Table 1). However, after normalizing with respect to subjects height, the length was larger (p = 0.003) only for Caucasian men compared to Caucasian women. The normalized width was smaller (p = 0.002, p = 0.013) for women compared to men in each ethnic group. The medial and lateral extension and flexion facet radii were also smaller for women compared to men in each ethnic group. However, after normalizing with respect to height, there were no differences, except for the femoral flexion facet radii for Caucasian men compared to Caucasian women (Table 1).
Differences in the group means of numerous demographic (Table 1) and kinematic (Table 2) variables were tested by one-way ANOVA. However, due to limited and unequal sample sizes the homoscedasticity assumption underlying the ANOVA test may not always have been satisfied. Moreover, one-way ANOVA is optimal for equal sample sizes, but it is conservative for unequal samples [21]. This could potentially lead to failing to reject the null hypothesis, while in fact the means are not equal (Type II error). Therefore, to determine differences between specific groups, the ANOVA was followed by a nonparametric post hoc Wilcoxon rank-sum (also known as Wilcoxon–Mann-Whitney) test, which does not assume equal sample sizes and does not require the data to be normally distributed. All statistical calculations were completed using Matlab® software (The MathWorks, Inc, Natick, MA).
Results
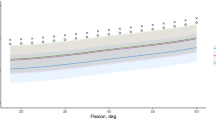
Caucasian women achieved similar (p = 0.403, p = 0.354) maximum flexion as the Japanese women and Japanese men (Table 2). These three groups also outnumbered the Caucasian males capable of achieving knee flexion greater than 150° (Fig. 6). Consequently, the average maximum flexion was lower for Caucasian men than for Caucasian women (p = 0.025) or Japanese men (p = 0.063) (Fig. 7A). These differences were also apparent when comparing the flexion ROM; the highest was attained by Japanese women, followed by Japanese men, Caucasian women, and Caucasian men.

The majority of the participants capable of achieving higher flexion were women and Japanese.

(A) Caucasian women and both Japanese groups achieved considerably higher (p = 0.028) maximum knee flexion than Caucasian men. (B) These three groups also revealed higher (p = 0.148) external femoral rotation than Caucasian men.
The maximum axial rotation was lower (p = 0.034) for Caucasian men compared to Caucasian women (Table 2). Caucasian women and both Japanese groups achieved similar average maximum axial rotation (Fig. 7B). The absolute range of axial rotation revealed a similar trend; it was lower for Caucasian men than for the three other groups, and there was no difference among the three other groups. When compared at each flexion increment, the femur rotated externally with increasing flexion, in a similar pattern for all four groups (Fig. 8A). However, in deep flexion, Caucasian men had the least amount of external rotation. For all groups, the femur was slightly abducted at the beginning of the activity but adducted with increasing knee flexion (Fig. 8B). Above 120° of flexion, the femur remained about 4° adducted for Caucasian women and both Japanese groups, while for Caucasian men, the adduction reduced and the femur became slightly abducted at 150° of flexion.

(A) The femur rotated externally with increasing flexion in a similar pattern for all four groups. However, Caucasian men revealed the lowest amount of external rotation in deep flexion. (B) For Caucasian women and both Japanese groups, the femur remained adducted above 120° of flexion, while for Caucasian men the femur actually abducted.
The lateral tibiofemoral contact point translated posteriorly in virtually the same pattern for all four groups and there was no difference at any flexion increment or in the absolute range of translation between any of the groups (Fig. 9). The AP translation of the medial tibiofemoral contact point was also similar for all groups, except at maximum flexion where it was more posterior (p = 0.0215) for Caucasian men than for Caucasian women. Similarly, the AP translation of the lateral femoral flexion facet center was similar between the groups at any flexion as was the medial femoral flexion facet center translation at each flexion increment. However, the posterior translation of the femoral flexion facet center from full extension to maximum flexion was higher (p = 0.089) for Caucasian men than for any other group.

No differences between any of the groups were found in the AP translation of (A) the lateral tibiofemoral contact point and (B) the lateral femoral flexion facet center. However, on the medial side in deep flexion, both the tibiofemoral contact point and the femoral flexion facet center translated more posteriorly for Caucasian men than for any other group.
The medial and lateral condyles moved in different modes (Fig. 10). The lateral condyle was spinning and rolling posteriorly throughout the ROM. However, the medial condyle countertranslated in the early flexion and then was spinning until 120°. Above 120°, the motion changed to more of a rolling type for both condyles, which may justify the use of the term “posterior femoral rollback.” There were no differences in the type of motion of the lateral condyle between the analyzed groups at any flexion increment. However, above 120° of flexion, the medial condyle revealed more rolling for Caucasian men compared to Caucasian women and both Japanese groups.

The lateral condyle was spinning and rolling posteriorly throughout the ROM and revealed no differences between any groups. However, (A) the medial condyle countertranslated in the early flexion, (B) then was spinning until 120°, and (C) finally rolled back to close to its initial position. This posterior rollback at the maximum flexion was highest for Caucasian men.
Discussion
Studies have suggested Caucasians achieve lower average knee flexion than non−Western populations and gender may influence condylar AP translation and axial rotation. We determined whether subjects of different gender or ethnicity reveal differences in their knee kinematics that may influence their ability to achieve high knee flexion. Identification of the motion patterns leading to higher flexion may help to improve existing TKA designs, which could restore higher postoperative ROM.
We acknowledge limitations associated with our study. Despite our best efforts, we were unable to recruit equally large sample sizes within each group. Therefore, we deemed it necessary to also use the post hoc test that does not require equal and normally distributed samples. Second, the Japanese population was recruited from the people currently living in the United States. Adjusting to the American lifestyle might have changed their native habits. However, we recruited only those subjects who grew up in Japan and moved to the United States as adults. Therefore, their bone morphology, if not the adaptation of the soft tissues, was established prior to their move and we believe that their kinematics outcomes remain representative of their ethnicity. Third, we estimated the tibiofemoral contact point location based on the distance between the tibial and femoral bone CAD models, which did not include the deformable cartilage. More accurate determination of the contact location could be made based on MRI scans. However, the long time required for image acquisition would make it impossible to study the activities under dynamic conditions. Moreover, based on the cartilage model compared to bone models, one study reported no differences in the location of the tibiofemoral contact points except for the lateral condyle at full extension [11].
Previous studies reported Caucasian populations achieve lower flexion (143.8° [40]) than non-Western populations (159.6° [1] and 157.3° [22] for Arabs, 156.9° for Indians [24], 162° for Japanese [37]) (Table 3). However, almost all subjects participating in these studies were men. We found lower maximum flexion for Caucasian men compared to Japanese men but no difference in flexion between Caucasian women and Japanese women. Similarly, we confirmed women in general achieve more flexion then men, but the difference was significant only for Caucasian population, but not for Japanese.
At full extension, we found the femur was internally rotated with respect to the tibia, which can be attributed to the screw-home mechanism being engaged at the arc of terminal extension [6, 16, 18, 20]. With increasing flexion, the femur rotated externally, similar to previously reported data [4, 14–16, 30, 31, 34]. The largest amounts of external rotation were observed between full extension and 30° of flexion, similar to previous studies of Caucasian [14] and Japanese populations [4]. Our data suggest the knee continues to rotate an additional 10° externally when flexed above 120°. Other studies reported an additional 13° [37] or even 20° [3] of external rotation above 120° of flexion for Japanese populations but suggested there is little longitudinal rotation observed for Caucasians during passive flexion [16]. Although in our study each group experienced this secondary increase of external rotation, it was indeed the lowest for Caucasian men. A previous study also reported lower range of tibial axial rotation for Caucasian men compared to women and suggested gender, in general, may be an affecting factor [46]. We found a similar difference for the Caucasian population, but there was no difference between Japanese genders; therefore, gender seemed to be an influencing factor only for Caucasians.
For Caucasian women and both Japanese groups the knee was continuously adducted above 30° of flexion. From the mechanical point of view, this suggests more of the tibiofemoral contact force was applied to the medial condyle, conceivably allowing the lateral condyle to more freely pivot about the medial tibiofemoral contact point. For Caucasian men, the knee abducted above 120° of flexion possibly shifting the contact load distribution more on the lateral condyle, which could hamper the pivoting and external femoral rotation.
The tibiofemoral contact points and the femoral flexion facet centers translated posteriorly with increasing flexion, with more translation observed on the lateral side, as widely reported in the literature [4, 11, 14–16, 29–31, 34, 41]. One study comparing a Caucasian population of 10 men and two women found no differences in the tibiofemoral movement [30], suggesting tibiofemoral kinematics was independent of gender, but the small sample sizes could not offer substantial support. We found the medial tibiofemoral contact point and femoral flexion facet center translated more posterior from full extension to maximum flexion for Caucasian men compared to Caucasian women, but there was no difference between the Japanese groups.
Our data confirm Caucasian women and both Japanese groups achieved similarly high knee flexion and suggest they achieve slightly higher axial rotation. For these three groups the femur remained adducted in deeper flexion. The Caucasian men who achieved the lowest axial rotation and whose knees were abducted in deeper flexion achieved the lowest femoral flexion and ROM. Our findings suggest higher axial rotation is not related to the lateral condyle, as its motion was almost identical for all three groups (the lateral femoral condyle was spinning and rolling in the same manner for all four groups [Fig. 10]). The differences were observed on the medial condyle. From full extension to maximum flexion, for Caucasian men, the medial tibiofemoral contact point and flexion facet center translated more posteriorly than for any other group. The medial condyle was also rolling more for the Caucasian men than for any other group above 120°. In fact, for the three other groups, the medial condyle remained more stationary, spinning close to the center of the tibial plateau, possibly allowing the lateral condyle to pivot about it and to rotate more externally. The ability to achieve more external rotation may also be related to ACL laxity, as it was higher for women than for men and for Japanese than for Caucasians [27, 42]. Also, the rotatory joint laxity is higher for women [28], which may contribute to higher ROM.
These findings may be important for future high flexion implant designs. We found subjects having a nonimplanted knee experienced more posterior femoral rollback of the lateral condyle, versus the medial condyle, leading to a normal axial rotation pattern where the tibia internally rotated with increasing flexion. In previous TKA studies we determined that subjects having a posterior stabilized TKA experienced similar rollback of their medial and lateral condyles, leading to minimal axial rotation and subjects having a posterior cruciate retaining TKA experienced a paradoxical motion where one or both condyles slid in the anterior direction with increasing knee flexion [8, 32, 50]. The future goal for newly designed TKA should be to induce more posterior femoral rollback of the lateral condyle, leading to the lateral condyle rotating around the medial condyle, but allowing the medial condyle to achieve a desired amount of translation in the anteroposterior direction.
References
Ahlberg A, Moussa M, Al-Nahdi M. On geographical variations in the normal range of joint motion. Clin Orthop Relat Res. 1988;234:229–231.
Anderson B, Burke E. Scientific, medical, and practical aspects of stretching. Clin Sports Med. 1991;10:63–86.
Andriacchi TP, Dyrby CO, Johnson TS. The use of functional analysis in evaluating knee kinematics. Clin Orthop Relat Res. 2003;410:44–53.
Asano T, Akagi M, Tanaka K, Tamura J, Nakamura T. In vivo three-dimensional knee kinematics using a biplanar image-matching technique. Clin Orthop Relat Res. 2001;388:157–166.
Austin B. Physical activity/exercise. In: Olshansky E, ed. Integrated Women’s Health: Holistic Approaches for Comprehensive Care. Gaithersburg, MD: Aspen Publishers, Inc; 2000:103.
Barnett C. Locking at the knee joint. J Anat. 1953;87:91–95.
Blaha J, Wojtys E. Motion and stability of the knee. In: Scott WN, ed. Surgery of the Knee. 4th ed. New York, NY: Churchill Livingstone; 2005:227–239.
Cates HE, Komistek RD, Mahfouz MR, Schmidt MA, Anderle M. In vivo comparison of knee kinematics for subjects having either a posterior stabilized or cruciate retaining high-flexion total knee arthroplasty. J Arthroplasty. 2008;23:1057–1067.
Chappell JD, Yu B, Kirkendall DT, Garrett WE. A comparison of knee kinetics between male and female recreational athletes in stop-jump tasks. Am J Sports Med. 2002;30:261–267.
Decker MJ, Torry MR, Wyland DJ, Sterett WI, Richard Steadman J. Gender differences in lower extremity kinematics, kinetics and energy absorption during landing. Clin Biomech (Bristol, Avon). 2003;18:662–669.
DeFrate LE, Sun H, Gill TJ, Rubash HE, Li G. In vivo tibiofemoral contact analysis using 3D MRI-based knee models. J Biomech. 2004;37:1499–1504.
Dennis DA, Komistek RD, Colwell CE Jr, Ranawat CS, Scott RD, Thornhill TS, Lapp MA. In vivo anteroposterior femorotibial translation of total knee arthroplasty: a multicenter analysis. Clin Orthop Relat Res. 1998;356:47–57.
Dennis DA, Komistek RD, Hoff WA, Gabriel SM. In vivo knee kinematics derived using an inverse perspective technique. Clin Orthop Relat Res. 1996;331:107–117.
Dennis DA, Mahfouz MR, Komistek RD, Hoff W. In vivo determination of normal and anterior cruciate ligament-deficient knee kinematics. J Biomech. 2005;38:241–253.
Freeman MA. How the knee moves. Curr Orthop. 2001;15:444–450.
Freeman MA, Pinskerova V. The movement of the knee studied by magnetic resonance imaging. Clin Orthop Relat Res. 2003;410:35–43.
Freeman MA, Pinskerova V. The movement of the normal tibio-femoral joint. J Biomech. 2005;38:197–208.
Fuss F. Principles and mechanisms of automatic rotation during terminal extension in the human knee joint. J Anat. 1992;180(Pt 2):297–304.
Grood ES, Suntay WJ. A joint coordinate system for the clinical description of three-dimensional motions: application to the knee. J Biomech Eng. 1983;105:136–144.
Hallen L, Lindahl O. The “screw-home” movement in the knee joint. Acta Orthop Scand, 1966;37:97–106.
Hayter A. A proof of the conjecture that the Tukey-Kramer multiple comparisons procedure is conservative. Ann Stat. 1984;12:61–75.
Hefzy MS, Kelly BP, Cooke TD. Kinematics of the knee joint in deep flexion: a radiographic assessment. Med Eng Phys. 1998;20:302–307.
Heitz NA, Eisenman PA, Beck CL, Walker JA. Hormonal changes throughout the menstrual cycle and increased anterior cruciate ligament laxity in females. J Athl Train. 1999;34:144–149.
Hemmerich A, Brown H, Smith S, Marthandam SS, Wyss UP. Hip, knee, and ankle kinematics of high range of motion activities of daily living, J Orthop Res. 2006;24:770–781.
Hoff WA, Komistek RD, Dennis DA, Gabrietl SM, Walker SA. Three-dimensional determination of femoral-tibial contact positions under in vivo conditions using fluoroscopy. Clin Biomech (Bristol, Avon). 1998;13:455–472.
Hollman JH, Deusinger RH, Van Dillen LR, Matava MJ. Gender differences in surface rolling and gliding kinematics of the knee. Clin Orthop Relat Res. 2003;413:208–221.
Hovinga KR, Lerner AL. Anatomic variations between Japanese and Caucasian populations in the healthy young adult knee joint. J Orthop Res. 2009;27:1191–1196.
Hsu WH, Fisk JA, Yamamoto Y, Debski RE, Woo SL. Differences in torsional joint stiffness of the knee between genders. Am J Sports Med. 2006;34:765–770.
Iwaki H, Pinskerova V, Freeman MA. Tibiofemoral movement 1: the shapes and relative movements of the femur and tibia in the unloaded cadaver knee. J Bone Joint Surg Br. 2000;82:1189–1195.
Johal P, Williams A, Wragg P, Hunt D, Gedroyc W. Tibio-femoral movement in the living knee. a study of weight bearing and non-weight bearing knee kinematics using “interventional” MRI. J Biomech. 2005;38:269–276.
Komistek RD, Dennis DA, Mahfouz MR. In vivo fluoroscopic analysis of the normal human knee. Clin Orthop Relat Res. 2003;410:69–81.
Komistek RD, Scott RD, Dennis DA, Yasgur D, Anderson DT, Hajner ME. In vivo comparison of femorotibial contact positions for Press-Fit posterior stabilized and posterior cruciate-retaining total knee arthroplasties. J Arthroplasty. 2002;17:209–216.
Mahfouz MR, Hoff WA, Komistek RD, Dennis DA. A robust method for registration of three-dimensional knee implant models to two-dimensional fluoroscopy images. IEEE Trans Med Imaging. 2003;22:1561–1574.
Mahfouz MR, Komistek RD, Dennis DA, Hoff WA. In vivo assessment of the kinematics in normal and anterior cruciate ligament-deficient knees. J Bone Joint Surg Am. 2004;86:56–61.
Malinzak RA, Colby SM, Kirkendall DT, Yu B, Garrett WE. A comparison of knee joint motion patterns between men and women in selected athletic tasks. Clin Biomech (Bristol, Avon) 2001;16:438–445.
Martelli S, Pinskerova V. The shapes of the tibial and femoral articular surfaces in relation to tibiofemoral movement. J Bone Joint Surg Br. 2002;84:607–613.
Nakagawa S, Kadoya Y, Todo S, Kobayashi A, Sakamoto H, Freeman MA, Yamano Y. Tibiofemoral movement 3: full flexion in the living knee studied by MRI. J Bone Joint Surg Br. 2000;82:1199–1200.
Noble PC, Gordon MJ, Weiss JM, Reddix RN, Conditt MA, Mathis KB. Does total knee replacement restore normal knee function? Clin Orthop Relat Res. 2005;431:157–165.
Pollard CD, Braun B, Hamill J. Influence of gender, estrogen and exercise on anterior knee laxity. Clin Biomech (Bristol, Avon). 2006;21:1060–1066.
Roaas A, Andersson GB. Normal range of motion of the hip, knee and ankle joints in male subjects, 30–40 years of age. Acta Orthop Scand. 1982;53:205–208.
Scarvell JM, Smith PN, Refshauge KM, Galloway H, Woods K. Comparison of kinematics in the healthy and ACL injured knee using MRI. J Biomech. 2005;38:255–262.
Sernert N, Kartus JT Jr, Ejerhed L, Karlsson J. Right and left knee laxity measurements: a prospective study of patients with anterior cruciate ligament injuries and normal control subjects. Arthroscopy. 2004;20:564–571.
Sharma L, Lou C, Felson DT, Dunlop DD, Kirwan-Mellis G, Hayes KW, Weinrach D, Buchanan TS. Laxity in healthy and osteoarthritic knees. Arthritis Rheum. 1999;42:861–870.
Sorrells RB, Stiehl JB, Voorhorst PE. Midterm results of mobile-bearing total knee arthroplasty in patients younger than 65 years. Clin Orthop Relat Res. 2001;390:182–189.
Stern SH, Insall JN. Posterior stabilized prosthesis: results after follow-up of nine to twelve years. J Bone Joint Surg Am. 1992;74:980–986.
Varadarajan KM, Gill TJ, Freiberg AA, Rubash HE, Li G. Gender differences in trochlear groove orientation and rotational kinematics of human knees. J Orthop Res. 2009;27:871–878.
Villar RN, Solomon VK, Rangam J. Knee surgery and the Indian knee. The importance of the preservation of flexion. Trop Doct. 1989;19:21–24.
Wojtys EM, Ashton-Miller JA, Huston LJ. A gender-related difference in the contribution of the knee musculature to sagittal-plane shear stiffness in subjects with similar knee laxity. J Bone Joint Surg Am. 2002;84:10–16.
Wojtys EM, Huston LJ, Schock HJ, Boylan JP, Ashton-Miller JA. Gender differences in muscular protection of the knee in torsion in size-matched athletes. J Bone Joint Surg Am. 2003;85:782–789.
Yoshiya S, Matsui N, Komistek RD, Dennis DA, Mahfouz M, Kurosaka M. In vivo kinematic comparison of posterior cruciate-retaining and posterior stabilized total knee arthroplasties under passive and weight-bearing conditions. J Arthroplasty. 2005;20:777–783.
Zeller BL, McCrory JL, Kibler WB, Uhl TL. Differences in kinematics and electromyographic activity between men and women during the single-legged squat. Am J Sports Med. 2003;31:449–456.
Acknowledgments
We thank William Badger, Jason Horan, and Matthew Anderle for assistance with MR and fluoroscopic imaging.
Author information
Authors and Affiliations
Corresponding author
Additional information
One or more of the authors (RDK, MRM, ALL) have received research grant from DePuy, Inc (Warsaw, IN). Each author certifies that he or she has no commercial associations (eg, consultancies, stock ownership, equity interest, patent/licensing arrangements, etc) that might pose a conflict of interest in connection with the submitted article.
Each author certifies that his or her institution approved the human protocol for this investigation, that all investigations were conducted in conformity with ethical principles of research, and that informed consent for participation in the study was obtained.
This work was performed at both the University of Rochester and the University of Tennessee.
Electronic supplementary material
Below is the link to the electronic supplementary material.
(MPG 852 kb)
(MPG 963 kb)
(MPG 11,130 kb)
About this article
Cite this article
Leszko, F., Hovinga, K.R., Lerner, A.L. et al. In Vivo Normal Knee Kinematics: Is Ethnicity or Gender an Influencing Factor?. Clin Orthop Relat Res 469, 95–106 (2011). https://doi.org/10.1007/s11999-010-1517-z
Published:
Issue Date:
DOI: https://doi.org/10.1007/s11999-010-1517-z