
Abstract
Biogenic amines are compounds, produced primarily by lactic acid bacteria (LAB) that negatively affect the wholesomeness of wine. Standard winemaking practices can greatly influence the levels of biogenic amines in wine. The aim of this study was to determine the relative contribution of different malolactic fermentation (MLF) practices and ageing of wines on fermentation lees to the final levels of biogenic amines. Wines were made on small scale over two harvest seasons with two red grape cultivars. Treatments included spontaneous MLF, co-inoculated MLF, MLF inoculated after alcoholic fermentation (conventional inoculation) and 4 months of ageing in the presence and absence of fermentation lees of all MLF treatments. Biogenic amine concentrations were measured by high-performance liquid chromatography at key winemaking stages and statistically analysed for the effects of MLF treatment and winemaking stage. Results indicate that the presence of indigenous LAB increased the risk of biogenic amine formation. Inoculation proved to reduce biogenic amine production over time compared to spontaneous MLF and co-inoculation even more than conventional inoculation. The presence of yeast lees during ageing generally led to higher final concentrations of biogenic amines in wines than the absence of lees. This study confirms other works that conclude that spontaneous MLF and uncontrolled ageing on yeast lees are generally unpredictable and pose a risk of biogenic amine contamination in finished wines.
Similar content being viewed by others


Explore related subjects
Discover the latest articles, news and stories from top researchers in related subjects.Avoid common mistakes on your manuscript.
Introduction
Biogenic amines are a group of nitrogenous compounds likely to occur in fermented foods and beverages that contain their precursors (free amino acids), can sustain the growth and metabolism of the causative microorganisms and favour the activity of the relevant decarboxylase enzymes. Wine is a medium in which these conditions are potentially satisfied. The main biogenic amines associated with wine are putrescine, histamine, tyramine and cadaverine. These are mainly the products of microbial decarboxylation of ornithine, histidine, tyrosine and lysine, respectively (Smit et al. 2008), although putrescine can also be formed via the arginine deiminase pathway from arginine (Arena and Manca de Nadra 2001; Mangani et al. 2005). Some biogenic amines, including putrescine, spermine and spermidine, are also formed by the metabolisms of plants and may be transferred from the grape berry to the must and finally to the wine (Halász et al. 1994). Many other biogenic amines such as phenylethylamine, agmatine, tryptamine, isoamylamine, methylamine and ethylamine have also been associated with wine (Glória et al. 1998; Marcobal et al. 2006; Soufleros et al. 2007; Smit et al. 2008).
Some biogenic amines are present at low levels in the human body and are involved in normal physiological functions. However, if an excessive amount of biogenic amines is ingested, or if the normal detoxification routes via amine oxidases are deficient or inhibited by substances such as anti-depressant drugs or ethanol, several physiological disorders can occur in sensitive humans (Ten Brink et al. 1990; Maynard and Schenker 1996; Shalaby 1996). This is of particular importance in wine where high levels of ethanol can increase biogenic amine toxicity due to amine oxidase inhibition. Thus, the presence of biogenic amines in wine should be avoided. Considering an increasing global awareness of “total food quality” (Giusti et al. 2008), wine producers have to carefully consider the impact of wine production and storage practices on the food safety and wholesomeness of their product.
Biogenic amines can be produced by various microorganisms associated with the different stages of wine production and storage. In the literature, it is generally agreed that the contribution to biogenic amine spoilage by yeast during alcoholic fermentation is much less significant than the contribution by lactic acid bacteria (LAB) during malolactic fermentation (MLF) (Granchi et al. 2005; Herbert et al. 2005; Marcobal et al. 2006). Biogenic amine formation in wine has been associated with species from all four genera of wine LAB. LAB are present on healthy grapes in low numbers and can be transferred to winery equipment, where they can remain present in significant numbers (Wibowo et al. 1985). These indigenous LAB are usually held responsible for spontaneous malolactic fermentations. However, the metabolic characteristics of the indigenous flora are usually unknown and may contain decarboxylase activities, which may lead to biogenic amine production. Oenococcus oeni is predominantly responsible for MLF, but species of Pediococcus and Lactobacillus, generally associated with spoilage, may survive and grow during MLF, particularly if the pH of the wine is above pH 3.5 (Wibowo et al. 1985; Lonvaud-Funel 1999; Du Toit et al. 2011). It was proposed by Aredes Fernandez et al. (2010) that biogenic amine content could also be increased under poor nutritional conditions when spoilage LAB with amino acid decarboxylase activity (such as Lactobacillus hilgardii strain 5w in their study, a histamine producer) are present during MLF and compete with commercial O. oeni strains. Commercial preparations of O. oeni are said to be selected for the absence of amino acid decarboxylases and are therefore unable to produce biogenic amines. This has been confirmed by a number of studies performed on existing and potential commercial malolactic starter cultures, in particular for histamine, tyramine and putrescine (Moreno-Arribas et al. 2003; Martín-Álvarez et al. 2006; Ruiz et al. 2010). Constantini et al. (2009) scrutinised the production of commercial starter cultures of yeast and bacteria used in winemaking and found that contamination by decarboxylase-positive, amine-producing bacteria is indeed possible in commercial preparations. In particular, they found the contaminants to be present in Saccharomyces cerevisiae starter cultures, while all examined O. oeni starters were found uncontaminated. Du Toit et al. (2011) recently reviewed the suitability of using Lactobacillus species as starter cultures for MLF, including the distribution and detection of biogenic amine producers within the genus. Inoculation of wine with a commercial MLF starter culture could thus potentially reduce the incidence of biogenic amines compared to spontaneous MLF in wines by domination of indigenous bacteria, which may contain decarboxylase activity.
After MLF, wine is usually submitted to clarifying and stabilising treatments and is aged (traditionally in oak barrels) from a few months to more than a year. White wines (such as from Burgundy) and sparkling wines produced by traditional secondary fermentation methods (such as méthode champenoise) were the only wine styles traditionally aged in contact with fermentation lees. However, today, it is becoming a more common practice to age red wines on the lees to increase the organoleptic complexity and balance of the wine by release of compounds during yeast and bacterial autolysis (Hernández et al. 2006; Alcaide-Hidalgo et al. 2007b).
The objectives of this study were to evaluate whether the timing of inoculation for MLF could influence the final biogenic amine content in wines and prove advantageous over spontaneous MLF, and to determine the risk of biogenic amine formation during ageing on the yeast lees.
Materials and Methods
Vinification, Malolactic Fermentation Treatments and Microorganisms
Pinotage and Cabernet Sauvignon grapes purchased from commercial wine cellars in the Paarl region, South Africa over two vintages (2006 and 2007) were used for this study. All treatments were repeated in duplicate in the 2006 harvest season and in triplicate in the 2007 season.
After grapes were destemmed and crushed, the skins and free-run juice were separated and homogenised. Equal volumes of the homogenised free-run juice and equal weights of the homogenised skins were aliquoted to each treatment in 10-L plastic buckets. Sulphur dioxide (10 to 20 mg/L) was added to the must of all treatments.
Malolactic fermentation treatments were comprised of spontaneous malolactic fermentation (treatment 1), two treatments co-inoculated for MLF (treatments 2 and 3) and two treatments conventionally inoculated for MLF (treatments 4 and 5). In treatment 1, spontaneous MLF was performed by the indigenous LAB. Commercial MLF starter cultures (Lalvin VP41®, Lallemand, France and Viniflora® Oenos, Chr. Hansen, Denmark) were inoculated into the fermenting grape musts of treatments 2 and 3, respectively, 24 h after inoculation with the yeast starter culture (co-inoculation). Treatments 4 and 5 were inoculated with Lalvin VP41® and Viniflora® Oenos, respectively, after the completion of alcoholic fermentation (conventional inoculation). All commercial yeast and MLF starter cultures added to wine during small-scale vinifications were inoculated according to the instructions of the manufacturer at the maximum recommended dosage.
Alcoholic fermentation was performed by S. cerevisiae strain NT202 (Anchor Yeast, South Africa) at room temperature (20 to 25 °C). Diammonium phosphate (0.2 to 0.5 g/L) was added to wines as yeast nitrogen source. Fermentation progress was monitored daily with a Brix hydrometer. Grape skins were punched down once daily throughout alcoholic fermentation. Wines were pressed with a hydraulic basket press at the completion of alcoholic fermentation and transferred to 4.5-L glass bottles sealed with airlocks to complete MLF. MLF progress was monitored by measurements of malic acid and lactic acid concentrations every 7 to 14 days by FT–IR (WineScan FT120, FOSS Analytical, Denmark) in all treatments. At the completion of MLF (malic acid <0.3 g/L), 50 to 80 mg/L SO2 was added to all wines. Each of treatments 1 to 5 was aged in the presence and absence of fermentation lees, respectively. Each original treatment was split into two new ageing treatments as follows: Half of the wine was racked from the treatment for ageing in the absence of lees (AL). The remaining half, containing all the fermentation lees, was used for ageing in the presence of lees (PL). Wines were aged in 750-mL glass bottles with screw caps for 4 months at ±15 °C.
Representative samples for analyses of biogenic amines and microbial enumeration were drawn in sterile sample vials. Samples were taken of the grape must, after alcoholic fermentation and after the completion of MLF for both cultivars in 2006 and 2007. After the 4-month ageing period, wines were again sampled for the analyses of biogenic amines in both seasons.
Enumeration of Microorganisms
The wine microbiological status was monitored in the must and at the completion of alcoholic fermentation and MLF by plate counts of colonies formed (cfu/mL) on selective agar media, followed by confirmation of cell morphology under a light microscope. One hundred microlitres of must or wine, diluted in sterile water in a ten-fold dilution series, was plated on selective media. All media were purchased from Biolab, Merck, Wadeville, Gauteng (South Africa), unless otherwise indicated. All plates were incubated at 30 °C for 5 to 10 days, depending on the growth rate of the microorganism.
De Man, Rogosa and Sharpe (MRS) agar was used for the enumeration of Lactobacillus, Pediococcus and Leuconostoc species (De Man et al. 1960). O. oeni was enumerated on MRS supplemented with 20% v/v apple juice at pH 5.2, incubated anaerobically. MRS agar supplemented with 2% v/v ethanol was used to isolate acetic acid bacteria in wine. GYC agar (5% w/v glucose, 1% w/v yeast extract, 3% v/v CaCO3 and 2% w/v agar) was used to isolate acetic acid bacteria from grape must. Lysine agar medium (lysine with 0.01% v/v potassium lactate) was used to establish non-Saccharomyces yeasts counts and yeast peptone dextrose agar (1% w/v yeast extract, 2% w/v peptone, 2% w/v glucose) to establish the total yeast population. WL nutrient agar for the enumeration of Brettanomyces species was purchased from Merck, Darmstadt, Germany. The pH of this medium was adjusted to 5.0 and 100 mg/L p-coumaric acid and 50 mg/L cycloheximide (both Sigma–Aldrich, Germany) were added to increase the selectivity of this medium. Pimaricin (50 mg/L) (Actistab, Gistbrocades, Anchor Bio-Technologies, South Africa), kanamycin sulphate (25 mg/L) (Roche, South Africa) and chloramphenicol (30 mg/L) (Sigma–Aldrich) were added to all the selective media as required to inhibit the growth of yeast, acetic acid bacteria and LAB, respectively.
Biogenic Amine Analyses
The four main biogenic amines associated with wine were analysed during this study (histamine, tyrosine, putrescine and cadaverine). In 2006, wine samples were analysed for biogenic amines by high-performance liquid chromatography based on the method described by Alberto et al. (2002). Samples were diluted 10 times and filtered through a 0.22-μm syringe filter prior to derivatisation and column injection. The derivatising reagent used was 200 mg o-phtaldialdehyde (Sigma) dissolved in 9 mL methanol, 1 mL 0.1 M sodium tetra-borate (pH 10) and 160 μL 2-mercaptoethanol. Twenty-five microlitres of the diluted sample reacted with 25 μL of derivatising reagent for exactly 45 s, and 25 μL of this solution was injected immediately thereafter. The derivatisation process was automated by the use of an auto sampler. In 2007, the services of a different laboratory was used, and biogenic amines were subsequently quantified in this season by liquid chromatography mass spectrometry, according to the methods described by Millán et al. (2007) and Smit (2007).
Univariate Statistical Analysis
Repeated-measures ANOVA was performed on the experimental data to evaluate main effects for fermentation treatment and phase, as well as their interactive effects. The Tukey HSD test was used to determine the significant differences between group means. A significance level of 5% was used in all cases. Statistical analyses were performed using STATISTICA (StatSoft, Inc. (2006) version 7.1. www.statsoft.com).
Results and Discussion
Biogenic Amine Content of Musts and Wines
Putrescine was present in the grape musts of both cultivars and vintages, with higher levels in 2006 than in 2007, and higher levels in Cabernet Sauvignon than in Pinotage grapes. Other biogenic amines were only detected at low concentrations in the grape must in the 2007 vintage (Table 1).
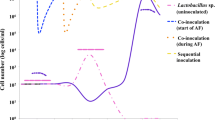
Figure 1 represents the concentrations of biogenic amines present in the different MLF treatments of the two cultivars and two vintages at three stages of winemaking, namely at the end of alcoholic fermentation, after MLF and after ageing for 4 months (PL or AL, respectively).

Histamine, putrescine and cadaverine produced in wine of two cultivars and two vintages. Biogenic amines were measured at the end of alcoholic fermentation, at the end of malolactic fermentation and after 4 months of ageing in the presence (PL) and absence (AL) of fermentation lees. Treatment 1 spontaneous MLF, Treatment 2 Co-inoculated MLF with VP41, Treatment 3 Co-inoculated MLF with Viniflora Oenos, Treatment 4 Conventional MLF with VP41, Treatment 5 Conventional MLF with Viniflora Oenos. Error bars indicate the means of treatment replicates
Histamine, putrescine and cadaverine were detected in the analysed wines. No tyramine was detected in any of the wines over both vintages. The concentrations of histamine produced at the end of MLF in Cabernet Sauvignon in 2006 in this study were between 3.4 and 6.7 mg/L. These amounts are of importance due to their trade implications, with acceptable upper limits in Europe ranging from 2 to 10 mg/L (Lehtonen 1996). The concentrations reached during the 2007 vintage were much lower than these recommended limits, although above the limit of quantification of our analytical method at 0.005 mg/L.
Relationship between MLF Inoculation Practices, Indigenous Microbial Population and Biogenic Amine Formation
In general, there were a few statistically significant differences in biogenic amine levels between different MLF inoculation practices. However, certain trends could be observed and will be discussed for each MLF treatment.
The different MLF inoculation scenarios had no impact on the growth of the commercial yeast strain or alcoholic fermentation speed. Non-Saccharomyces yeast strains (including Brettanomyces), acetic acid bacteria and Pediococcus strains were not detected in any of the wines and therefore did not contribute to biogenic amine formation.
The health condition of the grapes could have played a large role in the occurrence of indigenous decarboxylase-positive microorganisms and the physiological need to decarboxylate amino acids. For example, in Pinotage 2006, no Lactobacillus (or other indigenous) species were detected with selective plating once fermentations commenced (Table 2, End AF). Indigenous Lactobacillus species were present in the must of Cabernet Sauvignon 2006, increased in number and survived until the end of alcoholic fermentation. This higher incidence of indigenous LAB strains in Cabernet Sauvignon 2006 than in Pinotage 2006 could explain the higher incidence of histamine in this Cabernet Sauvignon, since the time of exposure to histidine decarboxylase-positive LAB was longer even in inoculated treatments. In Pinotage 2006 and 2007, Oenococcus could also only be detected at the end of alcoholic fermentation in the two treatments (treatments 2 and 3) that were co-inoculated together with alcoholic fermentation (no natural strains were present). In the Cabernet Sauvignon, indigenous O. oeni strains were present in the must and detected at the end of AF in all treatments, although at lower cell numbers in the spontaneous and conventional MLF treatments (not inoculated at this point) than the co-inoculated treatments.
It can be that biogenic amines may not be produced at significant concentrations, despite the presence of decarboxylase-positive bacteria, as also previously reported by Leitão et al. (2000). Decarboxylase activity and/or cell viability of LAB can be inhibited by wine conditions such as low pH and ethanol concentrations exceeding 12% v/v (Rollan et al. 1995; Leitão et al. 2000; Gardini et al. 2005). In particular, tyrosine decarboxylation never occurred in this study, and histamine was produced at very low concentrations in Pinotage 2006, 2007 and Cabernet Sauvignon 2007.
Examples of the correlation between biogenic amine formation and each MLF inoculation practice examined in this study are given below.
Spontaneous MLF
In the Pinotage 2006, histamine was only detected in one replicate of the spontaneous MLF treatment (treatment 1) after 4 months of ageing in the presence of yeast lees. In Cabernet Sauvignon 2006, the highest average histamine concentration was reached in the spontaneous MLF treatment (treatment 1), which was also the only treatment with residual histamine after 4 months of ageing with PL. Similarly, the highest histamine concentration was detected in Cabernet Sauvignon 2007, uninoculated (treatment 1) after MLF.
These observations indicate the potential variability and unreliability of spontaneous MLF in which the metabolic properties of the flora are generally unknown and may differ from one wine to the next (even between replicates). When no inoculation for MLF takes place, the wine is exposed to uncontrolled numbers of potential spoilage microbes, including decarboxylase-positive LAB, during the entire winemaking process. Spontaneous MLF therefore may be unpredictable and pose a larger risk of biogenic amine occurrence than in commercially inoculated MLFs. O. oeni starter cultures are specifically selected for their decarboxylase-negative status (Moreno-Arribas et al. 2003; Martín-Álvarez et al. 2006) and can thus greatly reduce the incidence of biogenic amine formation during MLF. Despite the widely held belief that organic wine production will increase the wholesomeness of wine due to its lower levels of added sulphur dioxide, its higher levels of biogenic amines (due to both low SO2 levels and spontaneous MLF) could pose a health risk to certain individuals, as also revealed in the study on quality red wines (including organic wine) by García-Marino et al. (2010).
Co-inoculated MLF
The concurrent inoculation of commercial strains of wine yeast and LAB in order to induce simultaneous alcoholic and malolactic fermentations can be a means to overcome potential inhibition of LAB in finished wines by high ethanol concentration, a shortage of nutrients and low temperatures. In our study, no antagonistic effects were observed during alcoholic fermentations that may have caused a delay in either alcoholic or malolactic fermentations.
For Cabernet Sauvignon 2007, the lowest average levels of histamine were found in co-inoculated treatments, when compared to spontaneous and conventional MLF. Similarly, the increase in cadaverine during ageing in this cultivar (2007) was also significantly lower in the co-inoculated treatments. In Cabernet Sauvignon 2006, the lowest putrescine levels were also found in co-inoculated treatments at the end of MLF. Due to the dominance of inoculated LAB cultures early on during alcoholic fermentation, contact between potential amino acid precursors in wine and spoilage (decarboxylase-positive) LAB is limited. O. oeni cell numbers in both cultivars and vintages indicate that co-inoculation successfully proceeded together with alcoholic fermentation and that MLF was indeed carried out and completed mainly by this species.
Inoculation with O. oeni starter cultures that are unable to produce biogenic amines and that may inhibit the growth and activity of decarboxylase-positive indigenous bacteria is a realistic option for the control of biogenic amines in wine. This has also been confirmed by other authors such as Marques et al. (2008) and Hernández-Orte et al. (2008). Inoculation with MLF starter cultures simultaneous with alcoholic fermentation may restrain these undesirable activities at an earlier stage than with conventional methods, decreasing the levels of biogenic amines in finished wine. In this study, it was shown that co-inoculation could reduce the incidence of biogenic amines in wine compared to conventional inoculation protocols. Similar results were obtained in a study by Van der Merwe (2007). It was also indicated in this work that in some cases the effect of co-inoculation on biogenic amine reduction may only be visible after a period of ageing (for example, histamine and cadaverine in Cabernet Sauvignon 2007, putrescine in Cabernet Sauvignon 2006).
Conventional MLF
In Cabernet Sauvignon 2007, the conventionally inoculated MLF treatments showed the highest means of histamine. Putrescine levels in Cabernet Sauvignon 2006 were also highest in conventional MLF treatments at the end of MLF, compared to co-inoculated and spontaneous treatments. Conventional inoculation also led to an increase in histamine after ageing on the lees, while all other treatments showed a decrease in histamine concentration.
Relationship between Ageing on the Yeast Lees and Risk of Biogenic Amine Formation
During our study, biogenic amines decreased during ageing with higher residual levels in treatments aged in the presence of yeast lees in a number of treatments. For example, histamine decreased in most cases during ageing in Cabernet Sauvignon of both vintages. In 2007, it was reduced more with AL than PL. Putrescine decreased during ageing in all inoculated treatments in Cabernet Sauvignon from both vintages, again with the reduction being more in AL than PL in 2007. Similarly, in Pinotage 2006 and 2007, putrescine decreased more in AL. Jiménez Moreno and Ancín Azpilicueta (2004) similarly reported that histamine reached a maximum concentration after the initial stages of ageing, where after it was progressively decomposed.
On the contrary, biogenic amine concentrations showed an increase in some treatments during ageing. The single replicate containing histamine in Pinotage 2006 was a treatment aged with lees. In Pinotage 2007, histamine concentrations were higher in all treatments after ageing compared to the end of MLF, with no difference attributed to the presence or absence of lees. However, it mostly appeared that in the case of biogenic amine increase during ageing, the increase was more in PL than in AL. Cadaverine increased during ageing in three of the four biological repeats; in Pinotage of both vintages, the increase was more significant with PL. Putrescine levels in Pinotage 2006 increased during ageing and were highest at the end of ageing and in PL. Our result obtained with Pinotage in 2006 is the only result that corresponds with the work done by Jiménez Moreno et al. (2003) and Jiménez Moreno and Ancín Azpilicueta (2004), who both reported that putrescine was accumulated and was not degraded during ageing.
In general, the frequency of biogenic amine occurrence in aged wines was higher in the presence of fermentation lees than in its absence in our study. Marques et al. (2008) and Alcaide-Hidalgo et al. (2007a) also found that wine storage on lees contributes to an increase in biogenic amine concentration. Hernández-Orte et al. (2008) reported an increase in biogenic amine production in wines during ageing for up to 6 months in oak barrels, particularly in treatments where spontaneous MLF had occurred (compared to inoculated MLF). In many cases in our study, biogenic amines also increased during ageing for 4 months. The reason for the initial increase of biogenic amines following the completion of MLF could be that SO2 added to the wine after MLF may not completely stop all biochemical reactions and enzyme activity (Coton et al. 1998; Lonvaud-Funel 2001). Another reason for increase in biogenic amines following MLF is the release of vitamins and nitrogen compounds such as proteins and polypeptides (that stimulate LAB activity) and free amino acids (that can act as biogenic amine precursors) due to yeast and bacterial autolysis and proteolytic activity of LAB (Manca de Nadra et al. 1997; Alcaide-Hidalgo et al. 2007a).
On the contrary, the decrease of some biogenic amines during ageing could presumable be due to amine oxidases present in some bacterial strains (Leuschner et al. 1998) that act by breaking down biogenic amines to an aldehyde, hydrogen peroxide and ammonia.
Conclusions
The conclusions of this work mainly confirm previous results of other authors who have studied the influence of winemaking conditions on biogenic amine production. The results obtained in this study confirm that all winemaking practices have the potential, when not controlled, to induce biogenic amine production. The two factors that posed the biggest risk in our study were the presence of high numbers of natural LAB (spontaneous MLF or delayed inoculation for MLF) and the ageing of wine on the yeast lees. Therefore, spontaneous MLF should be avoided to ensure good-quality wines, especially where the dominant flora consists of lactobacilli. It was also shown that co-inoculation compared to conventional inoculation (after alcoholic fermentation) reduced the risk due to early dominance over the natural flora. This would be applicable especially in high pH wines where the natural LAB numbers are higher and the species more diverse compared to low pH wines.
This study emphasises that controlled MLF and the timely separation of the fermentation lees after MLF are crucial factors to manage during the winemaking process to reduce the risk of biogenic amine production, thereby ensuring wine quality and safety.
References
Alberto, M. R., Arena, M. E., & Manca de Nadra, M. C. (2002). A comparative survey of two analytical methods for identification and quantification of biogenic amines. Food Control, 13, 125–129.
Alcaide-Hidalgo, J. M., Moreno-Arribas, M. V., Martín-Alvarez, P. J., & Polo, M. C. (2007). Influence of malolactic fermentation, postfermentative treatments and ageing with lees on nitrogen compounds of red wines. Food Chemistry, 103, 572–581.
Alcaide-Hidalgo, J. M., Moreno-Arribas, M. V., Polo, M. C., & Pueyo, E. (2007). Partial characterization of peptides from red wines. Changes during malolactic fermentation and ageing with lees. Food Chemistry, 107, 622–630.
Aredes Fernandez, P. A., Farias, M. E., & Manca de Nadra, M. C. (2010). Interaction between Oenococcus oeni and Lactobacillus hilgardii isolated from wine. Modification of available nitrogen and biogenic amine production. Biotechnology Letters, 32, 1095–1102.
Arena, M. E., & Manca de Nadra, M. C. (2001). Biogenic amine production by Lactobacillus. Journal of Applied Microbiology, 90, 158–162.
Constantini, A., Vaudano, E., Del Prete, V., Danei, M., & Garcia-Moruno, E. (2009). Biogenic amine production by contaminating bacteria found in starter preparations used in winemaking. Journal of Agricultural and Food Chemistry, 57, 10664–10669.
Coton, E., Rollan, G., Bertrand, A., & Lonvaud-Funel, A. (1998). Histamine-producing lactic acid bacteria in wines: Early detection, frequency, and distribution. American Journal of Enology and Viticulture, 49, 199–204.
De Man, J. C., Rogosa, M., & Sharpe, M. E. (1960). A medium for the cultivation of lactobacilli. Journal of Applied Bacteriology, 23, 130–135.
Du Toit, M., Engelbrecht, L., Lerm, E., & Krieger-Weber, S. (2011). Lactobacillus: The next generation of malolactic fermentation starter cultures—An overview. Food and Bioprocess Technology, 4, 876–906.
García-Marino, M., Trigueros, I., & Escribano-Bailón, T. (2010). Influence of oenological practices on the formation of biogenic amines in quality red wines. Journal of Food Composition and Analysis, 23, 455–462.
Gardini, F., Zaccarelli, A., Belletti, N., Faustini, F., Cavazza, A., Maruscelli, M., et al. (2005). Factors influencing biogenic amine production by a strain of Oenococcus oeni in a model system. Food Control, 16, 609–616.
Giusti, A. M., Bignetti, E., & Cannella, C. (2008). Exploring new frontiers in total food quality definition and assessment. From chemical to neurochemical properties. Food and Bioprocess Technology, 1, 130–142.
Glória, M. B. A., Watson, B. T., Simon-Sarkadi, L., & Daeschel, M. A. (1998). A survey of biogenic amines in Oregon Pinot noir and Cabernet Sauvignon wines. American Journal of Enology and Viticulture, 49, 279–282.
Granchi, L., Romano, P., Mangani, S., Guerrini, S., & Vincenzini, M. (2005). Production of biogenic amines by wine microorganisms. Bulletin de l’OIV, 78, 595–609.
Halász, A., Baráth, Á., Simon-Sarkadi, L., & Holzapfel, W. (1994). Biogenic amines and their production by microorganisms in food. Trends in Food Science & Technology, 5, 42–49.
Herbert, P., Cabrita, M. J., Ratola, N., Laureano, O., & Alves, A. (2005). Free amino acids and biogenic amines in wines and musts from the Alentejo region. Evolution of amines during alcoholic fermentation and relationship with variety, sub-region and vintage. Journal of Food Engineering, 66, 315–322.
Hernández, T., Estrella, I., Carlavilla, D., Martín-Álvarez, P. J., & Moreno-Arribas, M. V. (2006). Phenolic compounds in red wine subjected to industrial malolactic fermentation and ageing on lees. Analytica Chimica Acta, 563, 116–125.
Hernández-Orte, P., Lapeña, A. C., Peña-Gallego, A., Astrain, J., Baron, C., Pardo, I., et al. (2008). Biogenic amine determination in wine fermented in oak barrels. Factors affecting formation. Food Research International, 41, 697–706.
Jiménez Moreno, N., & Ancín Azpilicueta, C. (2004). Influence of wine turbidity on the accumulation of biogenic amines during ageing. Journal of the Science of Food and Agriculture, 84, 1571–1576.
Jiménez Moreno, N., Goñi, D. T., & Ancín Azpilicueta, C. (2003). Changes in amine concentrations during aging of red wine in oak barrels. Journal of Agricultural and Food Chemistry, 51, 5732–5737.
Lehtonen, P. (1996). Determination of amines and amino acids in wine – A review. American Journal of Enology and Viticulture, 47, 127–133.
Leitão, M. C., Teixeira, H. C., Barreto Crespo, M. T., & San Romão, M. V. (2000). Biogenic amines occurrence in wine: Amino acid decarboxylase and proteolytic activities expression by Oenococcus oeni. Journal of Agricultural and Food Chemistry, 48, 2780–2784.
Leuschner, R. G., Heidel, M., & Hammes, W. P. (1998). Histamine and tyramine degradation by food fermenting microorganisms. International Journal of Food Microbiology, 39, 1–10.
Lonvaud-Funel, A. (1999). Lactic acid bacteria in the quality improvement and depreciation of wine. Antonie Van Leeuwenhoek, 76, 317–331.
Lonvaud-Funel, A. (2001). Biogenic amines in wines: Role of lactic acid bacteria. FEMS Microbiology Letters, 199, 9–13.
Manca de Nadra, M. C., Farais, M. E., Moreno-Arribas, M. V., Pueyo, E., & Polo, M. C. (1997). Proteolytic activity of Leuconostoc oenos. Effect on proteins and polypeptides from white wine. FEMS Microbiology Letters, 150, 135–139.
Mangani, S., Geurrini, S., Granchi, L., & Vincenzini, M. (2005). Putrescine accumulation in wine: Role of Oenococcus oeni. Current Microbiology, 51, 6–10.
Marcobal, Á., Martín-Álvarez, P. J., Polo, C., Muñoz, R., & Moreno-Arribas, M. V. (2006). Formation of biogenic amines throughout the industrial manufacture of red wine. Journal of Food Protection, 69, 397–404.
Marques, A. P., Leitão, M. C., & San Romão, M. V. (2008). Biogenic amines in wines: Influence of oenological factors. Food Chemistry, 107, 853–860.
Martín-Álvarez, P. J., Marcobal, Á., Polo, C., & Moreno-Arribas, M. V. (2006). Influence of technological practices on biogenic amine contents in red wines. European Food Research and Technology, 222, 420–424.
Maynard, L. S., & Schenker, V. J. (1996). Monoamine-oxidase inhibition by ethanol in vitro. Nature, 196, 575–576.
Millán, S., Sampedro, M. C., Unceta, N., Goicolea, M. A., & Barrio, R. J. (2007). Simple and rapid determination of biogenic amines in wine by liquid chromatography-electronspray ionization ion trap mass spectrometry. Analytica Chimica Acta, 584, 145–152.
Moreno-Arribas, V., Polo, C. M., Jorganes, F., & Muñoz, R. (2003). Screening of biogenic amine production by lactic acid bacteria isolated from grape must and wine. International Journal of Food Microbiology, 84, 117–123.
Rollan, G. C., Coton, E., & Lonvaud-Funel, A. (1995). Histidine decarboxylase activity of Leuconostoc oenos 9204. Food Microbiology, 12, 455–461.
Ruiz, P., Izquierdo, P. M., Seseña, S., & Palop, M. L. (2010). Selection of autochthonous Oenococcus oeni strains according to their oenological properties and vinification results. International Journal of Food Microbiology, 137, 230–235.
Shalaby, A. R. (1996). Significance of biogenic amines to food safety and human health. Food Research International, 29, 675–690.
Smit, A. Y. (2007). Evaluating the influence of winemaking practices on biogenic amine production by wine microorganisms. MSc Thesis, Stellenbosch University, Private Bag X1, 7602 Matieland (Stellenbosch), South Africa.
Smit, A. Y., Du Toit, W. J., & Du Toit, M. (2008). Biogenic amines in wine: Understanding the headache. South African Journal of Enology and Viticulture, 29, 109–127.
Soufleros, E. H., Bouloumpasi, E., Zotou, A., & Loukou, Z. (2007). Determination of biogenic amines in Greek wines by HPLC and ultraviolet detection after dansylation and examination of factors affecting their presence and concentration. Food Chemistry, 101, 704–716.
Ten Brink, B., Damink, C., Joosten, H. M. L. J., & Huis in’t Veld, J. H. J. (1990). Occurrence and formation of biologically active amines in foods. International Journal of Food Microbiology, 11, 73–84.
Van der Merwe, H. (2007). The evaluation of malolactic fermentation starter cultures under South African winemaking conditions. MSc Thesis, Stellenbosch University, Private Bag X1, 7602 Matieland (Stellenbosch), South Africa
Wibowo, D., Eschenbruch, R., Davis, C. R., Fleet, G. H., & Lee, T. H. (1985). Occurrence and growth of lactic acid bacteria in wine: A review. American Journal of Enology and Viticulture, 36, 302–313.
Acknowledgement
The authors thank the National Research Foundation, Winetech and THRIP for financial support.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article
Cite this article
Smit, A.Y., du Toit, M. Evaluating the Influence of Malolactic Fermentation Inoculation Practices and Ageing on Lees on Biogenic Amine Production in Wine. Food Bioprocess Technol 6, 198–206 (2013). https://doi.org/10.1007/s11947-011-0702-8
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s11947-011-0702-8