
Abstract
Purpose of Review
The primary cilium is a non-motile microtubule-based organelle that senses a diverse range of extracellular signals. While recent studies highlight the importance of ciliary-dependent developmental signals, including Hedgehog, Wnt, and platelet-derived growth factor, it is not well understood whether and how bone morphogenetic protein (BMP) signaling, a key regulator of skeletogenesis, is involved in cilia-related bone developmental aspects and in the etiology of skeletal disorders.
Recent Findings
Increasing evidence suggests that osteoblast- or osteocyte-specific deletion of ciliary proteins leads to diverse skeletal malformations, reinforcing the idea that primary cilia are indispensable for regulating bone development and maintenance. Furthermore, it became evident that ciliary proteins not only contribute to ciliogenesis but also orchestrate cellular trafficking.
Summary
This review summarizes the current understanding of ciliary proteins in bone development and discusses the potential role of BMP signaling in primary cilia, enabling us to unravel the potential pathogenesis of skeletal ciliopathies.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
Bone is a dynamic tissue responsible for the architectural framework of the body that supports other connective tissues and protects internal organs in vertebrates. Bone also exerts important functions as an internal organ, such as calcium and phosphate storage and bone marrow support. Like other tissues, bone is being constantly remodeled. The highly coordinated physiological remodeling process involves bone resorption by osteoclasts and subsequent bone formation by osteoblasts. Osteocytes, the bone matrix-embedded descendants of osteoblasts, are also known to contribute to this process [1]. Whereas bone remodeling primarily occurs to repair damage and to allow adaptation to the mechanical environment in a site-specific manner, it also occurs to maintain mineral homeostasis in a non-site-specific manner [2]. In order to orchestrate bone remodeling, bone cells need to recognize the extracellular environment. However, the precise mechanisms to sense mechanical and chemical changes have yet to be elucidated.
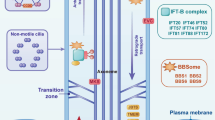
The primary cilium is a non-motile microtubule-based organelle present on the surface of most mammalian cell types. It is observed primarily in quiescent or differentiated cells [3], although certain differentiated cell lineages such as lymphocytes, hepatocytes, adipocytes, and skeletal muscle cells lack primary cilia [4]. Primary cilia are known to transduce a diverse range of extracellular signals into cellular responses via various signaling pathways (e.g. Hedgehog [Hh], Wnt, and platelet-derived growth factor [PDGF]) that regulate cell proliferation, differentiation, migration, and polarity, and are thereby crucial in tissue morphogenesis [5,6,7,8,9]. Although the mechanisms involved in the formation of primary cilia are unclear, there is increasing evidence suggesting that cilia formation and functions depend on a specialized protein transport process known as intraflagellar transport (IFT), which is mediated by a large multiprotein complex [10]. Because proteins are not synthesized within the cilia, they must be transported from the cytoplasm to build and maintain the primary cilia [4]. Any defect in the formation, maintenance, or functioning of primary cilia results in ciliopathies, a class of genetic multi-organ diseases that share clinical features and affect nearly every major tissue, including the kidney, brain, retina, liver, and bone. Their broad phenotypic spectrum highlights the critical role of primary cilia in development and homeostasis [11]. One of the remarkable phenotypes of ciliopathies is abnormal bone development, observed in Jeune syndrome, short-rib polydactyly, orofacial digital syndrome, and Ellis-van Creveld syndrome [12].
Because of the emerging interest in primary cilia in the field of skeletal development and disease, it is critical to understand their role in bone development and maintenance. This review summarizes recent advances in the understanding of the function of ciliary proteins in bone cells. Among growth factor signaling pathways, BMP is known as one of the most important signaling axes during bone development, maintenance, and disease, and its potential roles in ciliogenesis are also discussed.
Primary Cilia and Cell Cycle
Recent studies have suggested that primary cilium formation and cell cycle progression may reciprocally influence one another, although the exact mechanism remains to be elucidated [3]. In normal cells, primary cilia form during the quiescent state (G0/G1 phase) and begin to disassemble as cells re-enter the cell cycle [13]. One of the key components that link primary cilia and cell cycle is the centrosome. The centrosome, where spindle fibers appear during the mitotic phase, migrates to the cell surface, whereupon the mother centriole becomes the basal body of primary cilia [14]. Primary cilia cannot be formed during the mitotic phase, presumably because the basal body needs to be released from the cell surface before being translocated to the nucleolus to act as a mitotic component. This indicates that the conversion between the centriole and basal body of primary cilia is not only reversible, but also cell cycle-dependent [14]. In fact, centrosome dynamics are precisely synchronized with the cell cycle, therefore ciliogenesis and cell cycle are thought to be mutually exclusive events [15]. Moreover, many proteins known to be important for centrosome functions are also required for ciliogenesis [16, 17]. Hence, inhibiting the activity of ciliary proteins can promote cell cycle progression (IFT88/Polaris), disrupt the cell cycle (IFT27), or have no effect (OFD2) [3]. These findings thus indicate diverse cilia-independent functions for these ciliary proteins (discussed later).
Primary Cilia in Bone Cells
Although the existence of primary cilia in osteoblasts was first reported in the 1970s [18, 19], their possible contribution to bone homeostasis has only been recently proposed [20]. The significance of primary cilia in bone development and disease is widely recognized, but their incidence in bone cells is still controversial. A recent study reported that only a small fraction of bone cells (4.0% of osteocytes and 4.6% of bone-lining cells) possess primary cilia [21•], which is in accordance with a previous study [18]. However, it has also been reported that the majority of rat osteocytes (94%) possess acetylated, tubulin-positive primary cilia [22]. Such discrepancies between the two reports can be attributed to differences in the detection methods, animal species, location, or age of the animals analyzed.
Even though osteocytes are vital for regulating bone homeostasis, these cells do not divide further and most likely remain in the G0/G1 phase in vivo. In addition, osteocyte-specific disruption of ciliary proteins such as Pkd1 (Pkd1 Dmp1-cre) shows considerable phenotypic alterations [23]. As a result, it has been speculated that the majority of osteocytes possess primary cilia in vivo, but further studies are required to confirm these speculations.
Recent studies have reported that primary cilia are also present in mesenchymal stem cells (MSCs), the progenitors of osteoblasts [21, 24, 25]. To investigate their functional significance in these cells, IFT88/Polaris siRNA was used to inhibit primary cilia formation in MSCs, inducing a significant decrease in mRNA levels of all three (i.e., osteoblast, chondrocyte, and adipocyte) lineage-specific transcription factors [24]. MSCs transfected with the IFT88/Polaris siRNA and cultured in both osteogenic and adipogenic media lost cell adhesion, suggesting that primary cilia are required for osteogenic and adipogenic differentiation [24].
Osteoclasts, on the other hand, are derived from hematopoietic lineage cells [26], which are believed to lack primary cilia [4, 27]. However, a recent study reported the presence of primary cilia in such hematopoietic lineages, including peripheral blood mononuclear cells and acute myeloid leukemia cell lines [28•]. Although their presence in osteoclasts is still controversial, osteocytic primary cilia might play a role in bone resorption. Receptor activator of nuclear factor κB ligand (RANKL) and its antagonist osteoprotegerin (OPG), which are secreted by osteocytes, directly regulate osteoclast differentiation, and it has been reported that primary cilia regulate the mechanical stress-induced RANKL/OPG expression in osteocytes [29], thereby suggesting that they indirectly regulate osteoclastogenesis.
Several studies have reported that disruption of IFT proteins (e.g., IFT80, IFT88/Polaris), IFT motor proteins (e.g. KIF3A), and other ciliary proteins (e.g. EVC, polycystin) results in impaired osteoblast differentiation and bone formation [10]. Targeted deletion of Pkd1 in osteoblasts (Pkd1 Oc-cKO) induces abnormalities in Runx2-mediated osteoblast development and increases adipogenesis in the bone marrow, resulting in osteopenia [30, 31]. Similarly, selective deletion of Kif3a in osteoblasts (Kif3a Oc-cKO) disrupts primary cilia formation and function, also resulting in osteopenia [32]. In addition, Kif3a-deficient osteoblasts exhibit a reduction in response to fluid flow shear stress in vitro and impaired cilia-related pathways, such as Hh and Wnt signaling [32]. Remarkably, while deletion of Pkd1 alone leads to skeletal abnormalities, abrogation of both Pkd1 and Kif3a compensates for the skeletal phenotype. This indicates that crosstalk between Pkd1 and Kif3a might play a counterbalancing role in bone formation through the differential regulation of osteogenesis and adipogenesis [30].
Targeted deletion of IFT80 in osteoblast precursor cells (IFT80 Osx-cKO) results in significantly decreased bone mass due to impaired osteoblast differentiation [33••]. Deletion of IFT80 not only blocks canonical Hh-Gli signaling, but also stimulates cilia-independent non-canonical Hh-Gαi-RhoA signaling, resulting in excess stress fibers and subsequent osteoblast differentiation. Inhibition of non-canonical Hh signaling disrupts the formation of stress fibers and enhances cilia formation in IFT80-deficient osteoblasts. Therefore, IFT80 regulates osteoblast differentiation by balancing canonical Hh-Gli signaling and non-canonical Hh-Gαi-RhoA signaling.
Primary Cilia as a Mechanosensor in Osteocytes
Osteocyte-specific deletion of Pkd1 (Pkd1 Dmp1-cre) results in impaired sensory functions and disrupts mechanical stress-regulated skeletal homeostasis [23]. A recent study demonstrated that mechanical loading stimulates the homing of bone marrow cells to the bone surface, enhances osteogenic differentiation, and increases bone mass. In response to mechanical stimulation, deletion of Kif3a in the recruited osteoblasts significantly decreased the amount of bone formation [34••]. In response to oscillatory fluid flow-induced shear stress, the osteoblast and osteocyte cell lines MC3T3-E1 and MLO-Y4 displayed upregulated gene expression for osteopontin (Opn) and prostaglandin-E2 (PGE2), respectively, both of which are known to respond well to mechanical stimuli [29]. However, treatment with chloral hydrate or gene silencing by IFT88/Polaris siRNA reduced the number of ciliary cells and downregulated Opn and PGE2 gene expression. Interestingly, unlike in kidney cells, primary cilium-mediated cellular responses are independent of Ca2+ flux and stretch-activated ion channels [29], and a recent report further confirmed that primary cilia are not Ca2+-responsive mechanosensors in many cell types [35].
Primary Cilia and BMP Signaling
Despite many studies showing the critical importance of ciliary proteins in bone cells, the underlying signaling pathways remain elusive. Most of the current studies focus on investigating the ciliary-dependent Hh, PDGF, and Wnt pathways and on clarifying the significance of these signals during skeletogenesis [9, 10]. However, it is not known whether BMP, one of the most critical factors regulating bone development, is involved in cilia-mediated signal transduction. While it still remains unclear if BMP signaling is transduced by primary cilia in bone cells, recent studies have highlighted a potential link between transforming growth factor-β (TGF-β) signaling and primary cilia in retinal pigment epithelial cells [36] and fibroblasts [8]. Since BMP belongs to the TGF-β super-family [37], it would be reasonable to predict that BMP signaling may also be regulated by primary cilia, but the mechanism of how BMP is exactly transduced by primary cilia remains to be determined. Of note, BMP receptors are subject to endocytosis and trafficking when signaling is activated, and this regulatory mechanism is critical for the fine-tuning of BMP signaling response [38]. Because TGF-β signaling is transduced by primary cilia via endocytosis [8], ciliary-dependent BMP signaling may be orchestrated by the endocytosis machinery, which is likely critical for normal bone development and homeostasis.
Meanwhile, BMP signaling may also be required for primary cilia formation in bone cells. For example, deficiency of Acvr1, the gene encoding BMP type I receptor ACVR1, in mouse embryonic fibroblasts has been shown to result in severe defects in the formation of nodal primary cilia [39]. BMP signaling via ACVR1 positively regulates phosphorylation of cyclin-dependent kinase inhibitor p27KiP1, which in turn maintains the cells in quiescence and induces the formation of primary cilia. Moreover, BMP7 has been shown to inhibit the proliferation of human anaplastic thyroid carcinoma cells via stabilization of p21CiP1 and p27KiP1 [40]. In accordance with these findings, several mouse models of polycystic kidney disease, such as transgenic mice expressing Cux1 and Cys1, showed downregulation of p27KiP1 stability and its subsequent cell proliferation [41, 42]. In murine embryonic fibroblasts, a recombinant BMP2 and BMP4/7 fusion protein induced cell cycle arrest and stabilization of p27KiP1, thereby arresting the cell cycle in the G0/G1 phase [43]. In addition, BMP4-induced G0/G1 arrest mediates osteoblast differentiation through increased expression of p21CiP1 and p27KiP1, which are associated with BMPR-IA/ALK3-specific Smad1/5 phosphorylation [44]. Furthermore, BMPs are known to stimulate osteoblastic differentiation from their precursors [45, 46]; this supports the notion that BMPs are involved in the regulation of cell cycle, which is presumably important for ciliogenesis in osteoblasts.
In an Oak Ridge polycystic kidney (ORPK) mouse model, non-ciliated aortic endothelial cells acquired the ability to transdifferentiate into osteogenic cells in a BMP-dependent manner [47]. In addition, endothelial cells lacking primary cilia are sensitized to undergo BMP-induced osteogenic differentiation along with the activation of β-catenin [47]. These results suggest that primary cilia play an important role in BMP signaling by integrating it with other signaling pathways such as Wnt. Unfortunately, the currently available data on the role of BMP signaling are very limited, and it is unclear whether BMP signals act as a “ciliogenic” growth factor or are actually transduced by primary cilia in bone cells. Further studies are necessary to clarify these aspects, and the molecular pathogenesis of BMP-related disorders, including fibrodysplasia ossificans progressiva, [48•] will hopefully be fully uncovered in the future.
Cilium-Independent Functions of Ciliary Proteins
Recent studies have revealed that ciliary proteins are also found at certain non-cilia sites, where they have discrete roles in cellular function [49]. Interestingly, the disruption of each ciliary component results in a different phenotype, which suggests that these components are involved in processes other than cilium formation and functions. Phenotypic differences of ciliary protein disruption in bone cells are summarized in Table 1.
We have recently reported that a neural crest-specific deletion of IFT20 in mice compromises ciliogenesis and collagen intracellular trafficking, leading to osteopenia in the craniofacial region. These IFT20-mutant mice exhibited normal osteogenic differentiation potential, but the process of mineralization was severely attenuated due to the dysfunctional transport of procollagen from the endoplasmic reticulum (ER) to the Golgi apparatus. Hence, these data suggest that IFT20 is crucial for regulating not only ciliogenesis but also intracellular collagen transport [50••]. Additionally, previous studies demonstrated that non-ciliated T lymphocytes express several IFT proteins, as well as the IFT-dependent kinesin-2 motors, which are required for interaction of T cells with antigen-presenting cells and subsequent cytokine exocytosis [51]. In the non-ciliated T lymphocytes, IFT20 and other IFT proteins associate with both the centrosome and the Golgi apparatus, with a similar localization as that seen in ciliated cells [52]. These studies clearly demonstrated that IFT proteins are involved not only in cilia assembly/disassembly, but also in non-ciliary functions such as exocytosis.
Accumulating evidence indicates that many ciliary proteins exhibit more than one function in osteoblasts [50••] and in other cell types as well [49, 51, 53,54,55]. The contradicting findings based on the disruption of each ciliary protein are then most likely due to the multifunctional role and localization pattern of these proteins. Although ciliopathies comprise a group of genetic disorders associated with the disruption of cilium functions or defective expression of ciliary proteins, they display different phenotypes in distinct organs and at different stages of life. Therefore, the true etiology of ciliopathies may require re-evaluation and further investigation.
Conclusion
It has become increasingly evident that primary cilia act in a diverse range of cellular events involved in bone development and maintenance. A better understanding of cilia function can provide insights into the extracellular niche and environment in bone cells. The complexity of skeletal ciliopathies is due to the fact that ciliary proteins not only contribute to ciliogenesis but are also involved in mitosis and cellular trafficking. Therefore, it may be important to revise the notion that the phenotypes of skeletal ciliopathies are simply attributable to altered ciliary-dependent signals.
In this review, we highlighted a possible link between BMP, ciliogenesis and ciliary-dependent BMP signaling (Fig. 1). The functional output of BMP signaling relies on extensive crosstalk with other signaling pathways, including Hh, Wnt, Notch, mammalian target of rapamycin (mTOR), and receptor tyrosine kinases, which have been reported to be coordinated by the cilia/centrosome axis [56]. Defining the underlying mechanisms of BMP and primary cilia will contribute to the understanding of the interactions of BMP and other developmental signals critical for controlling bone development. Further studies are necessary to clarify the cilium-dependent and cilium-independent mechanisms of action of ciliary proteins, and to understand the precise pathogenesis of skeletal ciliopathies.

References
Papers of particular interest, published recently, have been highlighted as: •Of importance••Of major importance
Bonewald LF. The amazing osteocyte. J Bone Miner Res. 2011;26(2):229–38.
Burr DB. Targeted and nontargeted remodeling. Bone. 2002;30(1):2–4.
Santos N, Reiter JF. Building it up and taking it down: the regulation of vertebrate ciliogenesis. Developmental dynamics : an official publication of the American Association of Anatomists. 2008;237(8):1972–81.
Sanchez I, Dynlacht BD. Cilium assembly and disassembly. Nat Cell Biol. 2016;18(7):711–7.
Oh EC, Katsanis N. Cilia in vertebrate development and disease. Development. 2012;139(3):443–8.
Eggenschwiler JT, Anderson KV. Cilia and developmental signaling. Annu Rev Cell Dev Biol. 2007;23(1):345–73.
Berbari NF, O'Connor AK, Haycraft CJ, Yoder BK. The primary cilium as a complex signaling center. Curr Biol. 2009;19(13):R526–35.
Clement CA, Ajbro KD, Koefoed K, Vestergaard ML, Veland IR, Henriques de Jesus MP, et al. TGF-beta signaling is associated with endocytosis at the pocket region of the primary cilium. Cell Rep. 2013;3(6):1806–14.
Mourao A, Christensen ST, Lorentzen E. The intraflagellar transport machinery in ciliary signaling. Curr Opin Struct Biol. 2016;41:98–108.
Yuan X, Serra RA, Yang S. Function and regulation of primary cilia and intraflagellar transport proteins in the skeleton. Ann N Y Acad Sci. 2015;1335(1):78–99.
Novarino G, Akizu N, Gleeson JG. Modeling human disease in humans: the ciliopathies. Cell. 2011;147(1):70–9.
Yuan X, Yang S. Cilia/Ift protein and motor-related bone diseases and mouse models. Frontiers in bioscience (Landmark edition). 2015;20:515–55.
Tucker RW, Pardee AB, Fujiwara K. Centriole ciliation is related to quiescence and DNA-synthesis in 3t3-cells. Cell. 1979;17(3):527–35.
Kobayashi T, Dynlacht BD. Regulating the transition from centriole to basal body. J Cell Biol. 2011;193(3):435–44.
Nigg EA, Stearns T. The centrosome cycle: centriole biogenesis, duplication and inherent asymmetries. Nat Cell Biol. 2011;13(10):1154–60.
Gupta GD, Coyaud E, Goncalves J, Mojarad BA, Liu Y, Wu Q, et al. A dynamic protein interaction landscape of the human centrosome-cilium interface. Cell. 2015;163(6):1484–99.
Conduit PT, Wainman A, Raff JW. Centrosome function and assembly in animal cells. Nat Rev Mol Cell Biol. 2015;16(10):611–24.
Tonna EA, Lampen NM. Electron microscopy of aging skeletal cells. I. Centrioles and solitary cilia. J Gerontol. 1972;27(3):316–24.
Matthews JL, Martin JH. Intracellular transport of calcium and its relationship to homeostasis and mineralization: an electron microscope study. Am J Med. 1971;50(5):589–97.
Whitfield JF. Primary cilium—is it an osteocyte’s strain-sensing flowmeter? J Cell Biochem. 2003;89(2):233–7.
• Coughlin TR, Voisin M, Schaffler MB, Niebur GL, McNamara LM. Primary cilia exist in a small fraction of cells in trabecular bone and marrow. Calcif Tissue Int. 2015;96(1):65–72. This study demonstrated that only 4% of osteoblasts/osteocytes express primary cilia, indicating that cilia may act in a selected cell population
Uzbekov RE, Maurel DB, Aveline PC, Pallu S, Benhamou CL, Rochefort GY. Centrosome fine ultrastructure of the osteocyte mechanosensitive primary cilium. Microsc Microanal. 2012;18(6):1430–41.
Xiao Z, Dallas M, Qiu N, Nicolella D, Cao L, Johnson M, et al. Conditional deletion of Pkd1 in osteocytes disrupts skeletal mechanosensing in mice. FASEB journal: official publication of the Federation of American Societies for Experimental Biology. 2011;25(7):2418–32.
Tummala P, Arnsdorf EJ, Jacobs CR. The role of primary cilia in mesenchymal stem cell differentiation: a pivotal switch in guiding lineage commitment. Cell Mol Bioeng. 2010;3(3):207–12.
Hoey DA, Tormey S, Ramcharan S, O'Brien FJ, Jacobs CR. Primary cilia-mediated mechanotransduction in human mesenchymal stem cells. Stem Cells. 2012;30(11):2561–70.
Udagawa N, Takahashi N, Akatsu T, Tanaka H, Sasaki T, Nishihara T, et al. Origin of osteoclasts—mature monocytes and macrophages are capable of differentiating into osteoclasts under a suitable microenvironment prepared by bone marrow-derived stromal cells. Proceedings of the National Academy of Sciences of the United States of America. 1990;87(18):7260–4.
Finetti F, Paccani SR, Rosenbaum J, Baldari CT. Intraflagellar transport: a new player at the immune synapse. Trends Immunol. 2011;32(4):139–45.
• Singh M, Chaudhry P, Ramsingh G, Merchant A. Presence of primary cilia in human hematopoietic system. Blood. 2015;126(23):4762. This study demonstrated the presence of primary cilia in the hematopoietic system for the first time. The results implicate loss of primary cilia in the development of hematological malignancies through deregulated Hh signaling
Malone AM, Anderson CT, Tummala P, Kwon RY, Johnston TR, Stearns T, et al. Primary cilia mediate mechanosensing in bone cells by a calcium-independent mechanism. Proceedings of the National Academy of Sciences of the United States of America. 2007;104(33):13325–30.
Qiu N, Cao L, David V, Quarles LD, Xiao Z. Kif3a deficiency reverses the skeletal abnormalities in Pkd1 deficient mice by restoring the balance between osteogenesis and adipogenesis. PLoS One. 2010;5(12):e15240.
Xiao Z, Zhang S, Cao L, Qiu N, David V, Quarles LD. Conditional disruption of Pkd1 in osteoblasts results in osteopenia due to direct impairment of bone formation. J Biol Chem. 2010;285(2):1177–87.
Qiu N, Xiao Z, Cao L, Buechel MM, David V, Roan E, et al. Disruption of Kif3a in osteoblasts results in defective bone formation and osteopenia. J Cell Sci. 2012;125(Pt 8):1945–57.
•• Yuan X, Cao J, He X, Serra R, Qu J, Cao X, et al. Ciliary IFT80 balances canonical versus non-canonical hedgehog signalling for osteoblast differentiation. Nat Commun. 2016;7:11024. This study revealed that IFT80 is required for osteoblast differentiation by balancing the canonical Hh-Gli and non-canonical Hh-Galphai-RhoA pathways
• Chen JC, Hoey DA, Chua M, Bellon R, Jacobs CR. Mechanical signals promote osteogenic fate through a primary cilia-mediated mechanism. FASEB journal : official publication of the Federation of American Societies for Experimental Biology. 2016;30(4):1504–11. This study demonstrated that mechanical loading stimulates the homing of bone marrow cells to the bone surface and increases bone mass. In response to mechanical loading, deletion of Kif3a in the recruited osteoblasts significantly decreases the amount of bone formation
Delling M, Indzhykulian AA, Liu X, Li Y, Xie T, Corey DP, et al. Primary cilia are not calcium-responsive mechanosensors. Nature. 2016;531(7596):656–60.
Lindbæk L, Warzecha CB, Koefoed K, Mogensen JB, Schmid F, Pedersen LB, et al. Coordination of TGFβ/BMP signaling is associated with the primary cilium. Cilia. 2015;4(Suppl 1):P17.
Kawabata M, Imamura T, Miyazono K. Signal transduction by bone morphogenetic proteins. Cytokine Growth Factor Rev. 1998;9(1):49–61.
Ehrlich M. Endocytosis and trafficking of BMP receptors: regulatory mechanisms for fine-tuning the signaling response in different cellular contexts. Cytokine Growth Factor Rev. 2016;27:35–42.
Komatsu Y, Kaartinen V, Mishina Y. Cell cycle arrest in node cells governs ciliogenesis at the node to break left-right symmetry. Development. 2011;138(18):3915–20.
Franzen A, Heldin NE. BMP-7-induced cell cycle arrest of anaplastic thyroid carcinoma cells via p21(CIP1) and p27(KIP1). Biochem Biophys Res Commun. 2001;285(3):773–81.
Alcalay NI, Sharma M, Vassmer D, Chapman B, Paul B, Zhou J, et al. Acceleration of polycystic kidney disease progression in cpk mice carrying a deletion in the homeodomain protein Cux1. Am J Physiol Renal Physiol. 2008;295(6):F1725–34.
Cadieux C, Harada R, Paquet M, Cote O, Trudel M, Nepveu A, et al. Polycystic kidneys caused by sustained expression of Cux1 isoform p75. J Biol Chem. 2008;283(20):13817–24.
Thomas DM, Johnson SA, Sims NA, Trivett MK, Slavin JL, Rubin BP, et al. Terminal osteoblast differentiation, mediated by runx2 and p27KIP1, is disrupted in osteosarcoma. J Cell Biol. 2004;167(5):925–34.
Chang SF, Chang TK, Peng HH, Yeh YT, Lee DY, Yeh CR, et al. BMP-4 induction of arrest and differentiation of osteoblast-like cells via p21 CIP1 and p27 KIP1 regulation. Mol Endocrinol. 2009;23(11):1827–38.
Chen G, Deng C, Li YP. TGF-beta and BMP signaling in osteoblast differentiation and bone formation. Int J Biol Sci. 2012;8(2):272–88.
Wu M, Chen G, Li Y-P. TGF-β and BMP signaling in osteoblast, skeletal development, and bone formation, homeostasis and disease. Bone Research. 2016;4:16009.
Sanchez-Duffhues G, de Vinuesa AG, Lindeman JH, Mulder-Stapel A, DeRuiter MC, Van Munsteren C, et al. SLUG is expressed in endothelial cells lacking primary cilia to promote cellular calcification. Arterioscler Thromb Vasc Biol. 2015;35(3):616–27.
• Wang RN, Green J, Wang Z, Deng Y, Qiao M, Peabody M, et al. Bone Morphogenetic Protein (BMP) signaling in development and human diseases. Genes Dis. 2014;1(1):87–105. This study demonstrated that non-ciliated aortic endothelial cells acquire the ability to transdifferentiate into osteogenic cells in a BMP-dependent manner, and that β-catenin–induced Slug is a key transcription factor controlling this process
Vertii A, Bright A, Delaval B, Hehnly H, Doxsey S. New frontiers: discovering cilia-independent functions of cilia proteins. EMBO Rep. 2015;16(10):1275–87.
•• Noda K, Kitami M, Kitami K, Kaku M, Komatsu Y. Canonical and noncanonical intraflagellar transport regulates craniofacial skeletal development. Proceedings of the National Academy of Sciences of the United States of America. 2016;113(19):E2589-97. This study demonstrated that neural crest-specific deletion of IFT20 compromises not only ciliogenesis but also intracellular transport of collagen, which leads to osteopenia in the facial region.
Finetti F, Paccani SR, Riparbelli MG, Giacomello E, Perinetti G, Pazour GJ, et al. Intraflagellar transport is required for polarized recycling of the TCR/CD3 complex to the immune synapse. Nat Cell Biol. 2009;11(11):1332–U163.
Follit JA, Tuft RA, Fogarty KE, Pazour GJ. The intraflagellar transport protein IFT20 is associated with the Golgi complex and is required for cilia assembly. Mol Biol Cell. 2006;17(9):3781–92.
Baldari CT, Rosenbaum J. Intraflagellar transport: it’s not just for cilia anymore. Curr Opin Cell Biol. 2010;22(1):75–80.
Finetti F, Paccani SR, Rosenbaum J, Baldari CT. Intraflagellar transport: a new player at the immune synapse. Trends Immunol. 2011;32(4):139–45.
Ocbina PJ, Eggenschwiler JT, Moskowitz I, Anderson KV. Complex interactions between genes controlling trafficking in primary cilia. Nat Genet. 2011;43(6):547–53.
Lamouille S, Connolly E, Smyth JW, Akhurst RJ, Derynck R. TGF-beta-induced activation of mTOR complex 2 drives epithelial-mesenchymal transition and cell invasion. J Cell Sci. 2012;125(Pt 5):1259–73.
Hart EK, Jinnin M, Hou B, Fukai N, Olsen BR. Kinesin-2 controls development and patterning of the vertebrate skeleton by Hedgehog-and Gli3-dependent mechanisms. Dev Biol. 2007;309(2):273–84.
Haycraft CJ, Zhang Q, Song B, Jackson WS, Detloff PJ, Serra R, et al. Intraflagellar transport is essential for endochondral bone formation. Development. 2007;134(2):307–16.
Liu B, Chen S, Cheng D, Jing W, Helms JA. Primary cilia integrate hedgehog and Wnt signaling during tooth development. J Dent Res. 2014;93(5):475–82.
Temiyasathit S, Tang WJ, Leucht P, Anderson CT, Monica SD, Castillo AB, et al. Mechanosensing by the primary cilium: deletion of Kif3A reduces bone formation due to loading. PLoS One. 2012;7(3):e33368.
Yuan X, Yang S. Primary cilia and intraflagellar transport proteins in bone and cartilage. J Dent Res. 2016;95(12):1341–9.
Acknowledgements
We gratefully acknowledge Patricia Fonseca for editorial assistance. This study was supported by a Grant-in-Aid for Scientific Research from the Japan Society for the Promotion of Science (26293407, 15KK0337, and 15K15704 to MK), and by a grant from the NIDCR/NIH (R00DE021054 to YK).
Author information
Authors and Affiliations
Corresponding authors
Ethics declarations
Conflict of Interest
Masaru Kaku and Yoshihiro Komatsu declare no conflict of interest.
Human and Animal Rights and Informed Consent
This article does not contain any studies with human or animal subjects performed by any of the authors.
Additional information
This article is part of the Topical Collection on Skeletal Development
Rights and permissions
About this article

Cite this article
Kaku, M., Komatsu, Y. Functional Diversity of Ciliary Proteins in Bone Development and Disease. Curr Osteoporos Rep 15, 96–102 (2017). https://doi.org/10.1007/s11914-017-0351-6
Published:
Issue Date:
DOI: https://doi.org/10.1007/s11914-017-0351-6