
Abstract
Laboratory studies demonstrated that nymphs of Thyanta perditor (F.) completed their development on seed heads of wheat (Triticum aestivum L.) and barley (Hordeum vulgare L.), and on immature and mature seeds of black jack (Bidens pilosa L.), whereas no nymphs passed to the third instar when fed with leaves of any plant evaluated. At adult emergency, fresh body weight of females and males was greater on seed heads of wheat and barley (range 59–77 mg) compared to adults reared on immature and mature seeds of black jack (36–51 mg). Survivorship of adults after 40 days post eclosion on wheat seed heads, on green bean (Phaseolus vulgaris L.) pods, and on maize (Zea mays L.) seeds was > 80%, and was lower on barley seed heads (50%) and on soybean (Glycine max Merr.) pods (< 20%). For wild plants, on mature seeds and on black jack plant, the survivorship was higher than 50%. For reproduction, the most suitable diets were maize seeds, and wheat and barley seed heads (cultivated), and black jack (wild) plants. Twenty-eight days post eclosion, fresh body weight increased on all cultivated plants (26–62%), and less on the wild black jack plant (16%); on immature seeds and inflorescences of black jack, and on immature fruits of milkweed (Euphorbia heterophylla L.), adults lost weight (− 10, − 26, and − 17%, respectively).
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
Stink bugs (Pentatomidae) are mostly phytophagous and polyphagous on several cultivated plants of agricultural importance (Panizzi et al. 2000). The genus Thyanta encompass species commonly found on soybean (Glycine max Merr.), wheat (Triticum aestivum L.), and cotton (Gossypium hirsutum L.), among other crops. Thyanta custator (F.) and T. custator acerra McAtee (Reay-Jones 2010, 2014; Suh et al. 2013) are the most commonly referred species in the Nearctic (US), while in the Neotropics (Brazil), the red-shouldered stink bug, Thyanta perditor (F.), is the most common species, found mainly on wheat (Perez et al. 1980) and on soybean (Panizzi and Herzog 1984) crop plants.
Several stink bug species pests in the Neotropics, such as Dichelops melacanthus (Dallas) (Chocorosqui and Panizzi 2008), Nezara viridula (L.) (Panizzi and Meneguim 1989), Euschistus heros (F.) (Costa et al. 1998; Malaguido and Panizzi 1999), and Piezodorus guildinii (Westwood) (Panizzi and Smith 1977; Panizzi et al. 2002), had their biology evaluated on different cultivated and on wild plants. However, T. perditor, although considered a potential pest, has been little investigated and information on its nymphal and adult biology on cultivated and on wild plants is scarce. The few studies found in the literature report its damage to wheat plants (Ferreira and Silveira 1991), and notes on its biology on black jack (Bidens pilosa L.), soybean (Panizzi and Herzog 1984), and wheat (Perez et al. 1980).
In this study, we report results obtained in laboratory bioassays, in which we evaluated the performance of nymph and adult T. perditor feeding on cultivated and on wild plants. The cultivated plants chosen were soybean, maize (Zea mays L.) wheat, barley (Hordeum vulgare L.), and green bean (Phaseolus vulgaris L.), and the wild plants chosen were the weeds black jack, milkweed (Euphorbia heterophylla L.), signal grass (Brachiaria plantaginea Link), and flax-leaf fleabane (Conyza bonariensis L.). This plant selection was done based on the presence of T. perditor on these plants or on its potential to colonize them. Therefore, we tested the hypothesis that these cultivated and wild plants may or may not support the nymphal and adult biology of T. perditor.
Materials and methods
Stink bug colony and plants
Adults and nymphs of T. perditor were field-collected on black jack plants at the Embrapa National Wheat Research Center Field Experimental Station located in Passo Fundo, RS, Brazil (28°15′ S, 52°24′ W). Bugs were taken to the Laboratory of Entomology, where a colony was established. They were placed into plastic cages (25 × 20 × 20 cm) lined with filter paper, and kept in a walk-in chamber at 25 ± 1 °C temperature, 65 ± 10% relative humidity, and photoperiod of 14L:10D hours.
As food source, black jack plants carrying all reproductive structures (inflorescence, immature and mature seeds) were provided, which were replaced twice a week. Deposited egg masses were collected daily from the rearing cages and placed inside Petri dishes (9.0 × 1.5 cm) containing a fresh green bean pod (Phaseolus vulgaris L.) to obtain nymphs. Nymphs were used for the nymphal biology tests. Nymphs were also allocated in plastic cages (25 × 20 × 20 cm) and fed with a mixture of green bean pods, raw shelled peanuts (Arachis hypogaea L.), and branches of black jack containing inflorescence, immature and mature seeds, to obtain adults for the adult biology tests.
Plants of wheat cv. BRS Parrudo, barley cv. BRS Cauê, and soybean cv. BRS 284 and of black jack, milkweed, signal grass, and flax-leaf fleabane were obtained in the greenhouse. Seeds were placed in pots (5 L) containing a mixture of prepared soil every 2 weeks to grow the plants. These plants were cultivated during June to November 2016. Fresh green bean pod and maize (immature seed head) of unknown cultivars were obtained in local grocery stores.
Nymphal biology on cultivated and on wild plants
On the first day of the second instar (first instar stays grouped and do not feed), nymphs were carefully collected from the colony using a fine paint brush and placed into Petri dishes (9.0 × 1.5 cm) lined with filter paper. A wet cotton placed on a plastic lid (2 cm diameter) was used to provide water. The following plants were used as food source (10 nymphs/food) and replaced twice per week: cultivated—wheat, barley, soybean, green bean, and maize; wild—black jack, milkweed, signal grass, and flax-leaf fleabane.
During the vegetative plant stage, we evaluated from cultivated plants, leaves of maize, soybean, barley, and wheat, and from wild plants, leaves of black jack, signal grass, milkweed, and flax-leaf fleabane. During the reproductive plant stage, we evaluated from cultivated plants, immature seed heads of wheat and barley, immature pods of soybean and green bean, and immature seeds of maize. From the wild plants, we tested inflorescences, and immature and mature seeds of black jack, immature fruits of milkweed, immature seeds of signal grass, and mature seeds of flax-leaf fleabane.
The Petri dishes were placed randomly in an environmental chamber maintained at 25 ± 1 °C and 65 ± 5% RH and photoperiod of 14L:10D. Daily observations were made on nymph survivorship and development. From these observations, we calculated nymphal developmental time (for each instar and from second instar to adult) and percent mortality during each instar and from second instar to adult. At adult emergence, fresh body weight was measured using an electronic balance (Sartorius model BP210S).
Adult biology on cultivated and on wild plants
The adult performance was evaluated on five cultivated plants (wheat, barley, soybean, green bean, and maize) and on two wild plants (black jack and milkweed). The following treatments were used: cultivated plants—immature seed heads of wheat and barley, immature pods of soybean and green bean, and immature seeds of maize; wild plants—plants of black jack (carrying vegetative and reproductive structures), inflorescences, immature seeds, and mature seeds of black jack, and immature fruits of milkweed.
On the day of emergence, adults obtained from the laboratory colony were separated, weighed, and placed in pairs (female + male) in plastic boxes (11 × 11 × 3.5 cm) lined with filter paper and added a food source (n = 10 pairs per food treatment); for the treatment with black jack plant, a plastic cage (25 × 20 × 20 cm) was used. Wet cotton placed on a plastic lid (2 cm diameter) was used to provide water. The boxes were placed randomly in an environmental chamber maintained at 25 ± 1 °C and 65 ± 5% RH and photoperiod of 14L:10D.
At the end of 40 days of adult life, survivorship, fecundity (i.e., number of eggs and egg masses), and egg fertility (i.e., egg hatchability) were recorded. The parameters calculated were percent of ovipositing females, number of egg masses and eggs per female deposited, and percent of egg hatched. In addition, adults (females and males) had their fresh body weight, 28 days post eclosion, measured which was used to calculate percent of weight gain.
Statistical analysis
Data were previously analyzed using the Bartlett test to check for homogeneity of variances (P < 0.05), and then transformed to log(x) when necessary to attend the pre-requisites of analysis of variance (ANOVA). Nymphal and adult biology data were analyzed using ANOVA, and means were compared using the Tukey test (P < 0.05), except for comparison of the female’s body weight at the first day of the adult life, where means between barley (immature seeds head) and black jack (mature seeds) were compared using Student’s t test. All statistical analyses were carried out using R program version 3.3.1 (R Development Core Team 2016).
Results
Nymphal biology on cultivated and on wild plants
When fed during the vegetative stage of cultivated and wild plants, T. perditor nymphs were unable to complete their development (no nymphs reached the third instar). Nymphs survived on average from 4.7 to 5.1 days in the presence of leaves of cultivated plants (wheat = 5.1 ± 0.7 days, maize = 4.8 ± 0.4 days, barley = 4.8 ± 0.5 days, and soybean = 4.7 ± 0.5 days), and from 2.9 to 5.4 days on wild plants (signal grass = 5.4 ± 0.5 days, milkweed = 4.9 ± 0.5 days, flax-leaf fleabane = 4.5 ± 0.4 days, and black jack = 2.9 ± 0.3 days). On water only, nymphs survived for 4.5 ± 0.3 days.
When fed during the reproductive stage of cultivated plants, T. perditor nymphs were able to complete their development when fed on immature seed heads of barley (60%) and wheat (30%), but not on immature seeds of maize or on immature pods of soybean and of green bean (Table 1). On wild plants, T. perditor nymphs completed their development only on immature (40%) and on mature (70%) seeds of black jack. On mature seeds of flax-leaf fleabane, on inflorescences of black jack, on immature seeds of signal grass, and on immature fruits of milkweed, nymphs did not complete development (Table 1).
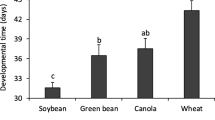
In general, the higher mortality was observed during the second instar, except on immature seeds of black jack where the fifth instar had highest mortality (Table 1). Even though some differences were observed during each instar, the total nymphal developmental time was not significantly different among those fed on immature seed heads of barley and wheat for cultivated plants, and on immature and mature seeds of black jack for wild plants (ranged on average from 31 to 34.3 days) (Table 1). On immature seeds of maize, immature pods of soybean and green bean, immature seeds of signal grass, and immature fruits of milkweed, no nymphs pass to the 3rd instar.
At the first day of the adult life, fresh body weight of females and males showed statistical differences when fed on cultivated and on wild plants on which nymphs completed development. Both females and males presented a significantly higher fresh body weight when fed on immature seed heads of the cultivated plants (barley and wheat; range 59–77 mg) compared to wild plants (immature and mature seeds of black jack; range 35.5–51 mg). On immature seeds of black jack, males presented the lowest body weight, whereas on mature seeds of this weed plant they remained in an intermediate position (Fig. 1).

Fresh body weight (mg ± SE) of males and females Thyanta perditor at the first day of the adult life feeding on reproductive structures of cultivated (barley immature seed heads, n = 6; wheat immature seed heads, n = 3) and wild plants (black jack mature seeds, n = 7; black jack immature seeds, n = 4) in laboratory conditions. Means (± SE) followed by the same letter among foods in each sex do not differ significantly (Tukey’s test, P < 0.05), except for female’s body weight, where means of barley (immature seed heads) and black jack (mature seeds) were compared using Student’s t test (t value = 15.634, df = 7, P value < 0.001). *Data were not considered for statistical analyses because only one female was obtained
Adult biology on cultivated and on wild plants
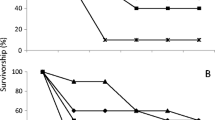
Survivorship of females and males T. perditor up to 40 days of life was variable according to the food source utilized. On the cultivated plants, on wheat seed heads, green bean pods, and maize seeds, both females and males presented over 80% of survivorship after 40 days (Fig. 2a–c); however, the survivorship of females and males decreased over time on barley seed heads (50%) and mostly on soybean pods (20 and 10%, respectively) (Fig. 2d, e).

Survivorship (%) of adult females and males Thyanta perditor (n = 10) up to 40 days post eclosion feeding on cultivated plants in laboratory conditions. Wheat: immature seed heads (a), green bean: immature pods (b), maize: immature seeds (c), barley: immature seed heads (d), and soybean: immature pods (e)
On the other hand, on all wild plants, the survivorship of both genders decreased over time. Passed 40 days, females and males fed on black jack plants (Fig. 3a) and males fed on mature seeds of black jack (Fig. 3b) showed survivorship higher or equal to 50%. On immature seeds and on inflorescences of black jack, and immature fruits of milkweed, survivorship decreased more sharply, in which more than 50% of males and females were dead after 20 days, and none reached day 40 (Fig. 3c–e).

Survivorship (%) of adult females and males Thyanta perditor (n = 10) up to 40 days post eclosion feeding on wild plants in laboratory conditions. Black jack: plants (a), black jack: mature seeds (b), black jack: immature seeds (c), black jack: inflorescences (d), and milkweed: immature fruits (e)
For the reproductive performance of T. perditor, the percentage of females that oviposited varied from 30 to 100% among cultivated plants and from 0 to 100% among wild plants. On soybean pods, only 30% of females laid eggs; however, on the other cultivated plants (green bean pods, seed heads of wheat and barley, and maize seeds) the majority (> 70%) of females oviposited (Table 2). On wild plants, all females laid eggs when fed on black jack plants; however, on mature and on immature seeds of black jack, only 40 and 10% of females laid eggs, respectively; on inflorescences of black jack and on immature fruits of milkweed, no females laid eggs (Table 2).
Regarding the fecundity of T. perditor, the number of egg masses/female was significantly higher when they were fed on maize seeds, and wheat and barley seed heads on cultivated plants; on the wild plants—black jack—fecundity was also significantly higher (over 2 × more egg masses/female) compared to the remaining food sources (Table 2). As a result of this higher fecundity, the total number of eggs per female was also significantly higher (range 136–207 eggs) on these referred food sources; on the remaining food sources, the number of eggs/female was significantly lower (range 9–54 eggs) (Table 2).
Egg hatchability showed variability among the foods provided; on plants where females laid the highest number of eggs (maize seeds, wheat and barley seed heads, and black jack plants), it ranged from 56 to 68%. Percentage values of egg hatchability tended to be higher on green bean pods and on immature and mature seeds of black jack (> 75%); however, few eggs were laid on these foods (Table 2).
Fresh body weight of adult T. perditor after 28 days presented significant differences among foods. On cultivated plants, the fresh body weight did not show significant differences among them (range 85–98 mg). Percentage gain weight was higher on maize seeds (62%), followed by wheat seed heads (43%), soybean pods (39%), green bean pods (34%), and barley seed heads (26%) (Fig. 4). On wild plants, fresh body weight was, in general, smaller, except for black jack plants (on average 86 mg), which did not differ significantly from the cultivated plants. Body weight decreased significantly on mature seeds of black jack (on average 77 mg), and achieved the lowest values on inflorescences and immature seeds of black jack, and on immature fruits of milkweed (< 58 mg). Percentage weight gain occurred only on mature seed of black jack (23%) and on black jack plants (16%), whereas on immature seeds and inflorescences of black jack and on immature fruits of milkweed, adults lost weight (− 10, − 26, and − 17%, respectively) (Fig. 4).

Fresh body weight (mg ± SE) of (female + male) Thyanta perditor after 28 days of the adult life feeding on cultivated (wheat, n = 19; green bean = 19; maize = 18; soybean = 6; barley = 17) and on wild (black jack plants, n = 17; mature seeds = 12; immature seeds = 8; inflorescences = 5; milkweed immature seeds = 1) plants in laboratory conditions. Means (± SE) followed by the same letter are not significantly different (Tukey’s test, P < 0.05). *Data were not considered for statistical analyses because only one individual survived. In parentheses, percentage of body weight gain in the period
Discussion
Thyanta perditor, as the majority of pentatomid pests of crops of economic importance (Panizzi 1997), feeds preferably on seeds, as reported by Ferreira and Silveira (1991) on wheat plants. In our study, this feeding preference for reproductive structures was clearly demonstrated by the fact that T. perditor nymphs did not developed on vegetative structures (leaves) of both the cultivated and wild plants tested. In fact, nymphs survived for about 5 days only on these leaves before perishing, similar to what happened when they took water only. Even though T. perditor nymphs did not survive when fed only on vegetative structures (leaves), Tomacheski and Panizzi (2018) observed that T. perditor adults had preference for seedlings of soybean and maize compared to other cultivated plants, and for the weed plants, milkweed and black jack were preferred, when compared to both cultivated and wild plants.
Therefore, on vegetative stage of cultivated and on wild plants, T. perditor may exploit them as shelter and/or as plant juice source for rehydration, which is known to occur in other stink bug species, such as Edessa meditabunda (F.) (Lucini and Panizzi 2016), which feed preferably on plant stems (Silva et al. 2012), and P. guildinii (Westwood) (Lucini et al. 2016), which feed preferably on seeds (Panizzi and Smith 1977). Much probably, the nutrient concentration in leaves of cultivated and wild plants, therefore, is not enough to allow T. perditor nymphs to complete their development.
The fact that nymphs of T. perditor did survive on seed heads of barley and wheat in our laboratory tests explains in part the presence of adults on these plants in the field. However, their relatively low survivorship, as compared to other species of phytophagous pentatomids (see review in Panizzi 1997), may explain the fact that they are seldom observed on seed heads of cereal plants. Therefore, most adults observed on the seed heads might have developed as nymphs elsewhere. On the other cultivated Poaceae (immature seeds of maize) and Fabaceae (immature pods of soybean and green bean), no nymphs survived, demonstrating their unsuitability as food sources for T. perditor nymphs.
On wild plants, nymphs did survive and complete development on black jack (immature and mature seeds), confirming previous data on T. perditor nymphal biology on this food source (Panizzi and Herzog 1984). However, on our study, we add novel information on the relation of the bug with black jack, demonstrating that nymphs develop better on mature than on immature seeds, and that, although observed in the field, nymphs do not complete development when fed exclusively on black jack inflorescences. The other three remaining weed plants tested (flax-leaf fleabane mature seed, signal grass immature seed, and milkweed immature fruit) were unsuitable foods for nymphs.
The higher fresh body weight attained at adult emergency on seed heads of barley and wheat, and on mature seeds of black jack shows that, despite the high nymph mortality, the ones that are able to reach adulthood, originate bigger adults. The adult survivorship, reproduction, and weight gain data demonstrate that, in general, food sources suitable for nymphs were also suitable for adults. However, some drastic differences were observed. For example, all nymphs died when fed on immature seeds of maize; adult reproduction, however, was highest on this diet.
Maize seedlings may be attacked by stink bugs, such as the so-called in Brazil green-belly stink bugs, D. melacanthus (Dallas) (Chocorosqui and Panizzi 2008) and D. furcatus (F.) (Panizzi et al. 2015), and by the Neotropical brown stink bug, E. heros (F.) (Quintela et al. 2006). Moreover, Tomacheski and Panizzi (2018) reported the preference of T. perditor adults for maize seedlings compared to wheat and barley seedlings; however, it was considered an unexpected result, because T. perditor is not reported to occurs on maize, whereas on wheat it does (Smaniotto and Panizzi 2015). On maize seed heads, few reports are found in the literature on true bugs exploring this as a food source: the cosmopolitan southern green stink bug, N. viridula (L.) in Louisiana (USA) (Negron and Riley 1987), and the coreid, Leptoglossus zonatus (Dallas) in Brazil (Panizzi 1989).
Apparently, adult pentatomids and other true bugs are unable to identify maize seed heads as a rich and nutritious food source or are unable to reach the seeds covered by the maize seed head husk. Also, the fact that nymphs died when fed on exposed immature maize seeds while adults reached the highest reproduction suggest that the nymphal development and adult reproduction syndromes apparently require different nutrients and/or nutrients on different concentrations. Clearly, additional investigations should be conducted to confirm these data.
Interesting to note was the fact that on black jack immature seeds, both, nymphs and adults, did not perform well. The majority of nymphs did not complete development, adults (males and females) did not reach day 40 and few reproduced, and they lost weight after 28 days. On mature seeds, however, the opposite was observed, i.e., the majority of nymphs completed development, both males and females reached day 40, reproduction was high, and they gained weight after 28 days. In fact, in natural conditions, nymphs are exclusively observed feeding on mature seeds of black jack. Apparently immature seeds do not have all nutrients needed for nymph and adult to perform well or may present toxic allelochemicals that cause detrimental effects compared to mature seeds.
These laboratory studies coupled with field observations allow to conclude that the stink bug T. perditor may eventually reach pest status on the winter cereals of southern Brazil, such as wheat and barley. Among the wild plants, black jack is the most common host plant, while the other weed plants tested play a minor role on its biology.
References
Chocorosqui VR, Panizzi AR (2008) Nymphs and adults of Dichelops melacanthus (Dallas) (Heteroptera: Pentatomidae) feeding on cultivated and non-cultivated host plants. Neotrop Entomol 37:353–360
Costa MLM, Borges M, Vilela EF (1998) Reproductive biology of Euschistus heros (F.) (Heteroptera: Pentatomidae). An Soc Entomol Bras 27:559–568
Ferreira E, Silveira PM (1991) Grain yield losses due to Thyanta perditor (Hemiptera: Pentatomidae) in wheat (Triticum aestivum L.). An Soc Entomol Bras 20:165–171
Lucini T, Panizzi AR (2016) Waveform characterization of the soybean stem feeder Edessa meditabunda (F.) (Hemiptera: Heteroptera: Pentatomidae): overcoming the challenge of wiring pentatomids for EPG. Entomol Exp Appl 158:118–132
Lucini T, Panizzi AR, Backus EA (2016) Characterization of an EPG waveform library for redbanded stink bug, Piezodorus guildinii (Hemiptera: Pentatomidae) on soybean plants. Ann Entomol Soc Am 109:198–210
Malaguido AB, Panizzi AR (1999) Nymph and adult biology of Euschistus heros (Hemiptera: Pentatomidae) and its abundance related to planting date and phenological stages of sunflower. Ann Entomol Soc Am 92:424–429
Negron JF, Riley TJ (1987) Southern green stink bug, Nezara viridula (Heteroptera: Pentatomidae) feeding on corn. J Econ Entomol 80:666–669
Panizzi AR (1989) Desempenho de ninfas e adultos de Leptoglossus zonatus (Hemiptera: Coreidae) em diferentes alimentos. An Soc Entomol Bras 18:375–389
Panizzi AR (1997) Wild hosts of pentatomids: ecological significance and role in their pest status on crops. Annu Rev Entomol 42:99–122
Panizzi AR, Herzog DC (1984) Biology of Thyanta perditor (Hemiptera: Pentatomidae). Ann Entomol Soc Am 77:646–650
Panizzi AR, Meneguim AM (1989) Performance of nymphal and adult Nezara viridula on selected alternate host plants. Entomol Exp Appl 50:215–223
Panizzi AR, Smith JG (1977) Biology of Piezodorus guildinii: oviposition, development time, adult sex ratio, and longevity. Ann Entomol Soc Am 70:35–39
Panizzi AR, McPherson JE, James DG, Javahery M, McPherson RM (2000) Stink bugs (Pentatomidae). In: Schaefer CW, Panizzi AR (eds) Heteroptera of economic importance. CRC Press, Boca Raton, p 421–474
Panizzi AR, Cardoso SR, Chocorosqui VR (2002) Nymph and adult performance of the small green stink bug, Piezodorus guildinii (Westwood) on lanceleaf crotalaria and soybean. Braz Arch Biol Technol 45:53–58
Panizzi AR, Agostinetto A, Lucini T, Smaniotto LF, Pereira PRVS (2015) Manejo integrado dos percevejos barriga-verde, Dichelops spp. em trigo. Documentos 114. Embrapa Trigo, Passo Fundo
Perez CA, Souza JLF, Nakano O (1980) Observações sobre a biologia e hábitos do percevejo Thyanta perditor (F.) (Hemiptera-Pentatomidae) em planta de trigo. Solo 72:61–62
Quintela ED, Francisco AJ, Ferreira SB, Oliveira LFC, Lemes ACO (2006) Efeito do tratamento de sementes com inseticidas químicos sobre danos de percevejos fitófagos e sobre a lagarta do cartucho no milho. Circ Téc 76. Embrapa Arroz e Feijão, Santo Antônio de Goiás
R Development Core Team (2016) R: a language and environment for statistical computing. R Foundation for Statistical Computing, Vienna
Reay-Jones FPF (2010) Spatial and temporal patterns of stink bugs (Hemiptera: Pentatomidae) in wheat. Environ Entomol 39:944–955
Reay-Jones FPF (2014) Spatial distribution of stink bugs (Hemiptera: Pentatomidae) in wheat. J Insect Sci 14:1–22
Smaniotto LF, Panizzi AR (2015) Interactions of selected species of stink bugs (Hemiptera: Heteroptera: Pentatomidae) from leguminous crops with plants in the Neotropics. Fla Entomol 98:7–17
Silva FAC, Silva JJ, Depieri RA, Panizzi AR (2012) Feeding activity, salivary amylase activity and superficial damage to soybean seed by adult Edessa meditabunda (F.) and Euschistus heros (F.) (Hemiptera: Pentatomidae). Neotrop Entomol 41:386–390
Suh CPC, Westbrook JK, Esquivel JF (2013) Species of stink bugs in cotton and other row crops in the Brazos River Bottom of Texas. Southwest Entomol 38:561–570
Tomacheski JF, Panizzi AR (2018) Seasonal abundance of Thyanta perditor (F.) (Heteroptera: Pentatomidae) and its preference among cultivated and non-cultivated plants. Rev Agropec Catar 31 (in press)
Acknowledgements
This study was partially supported by a National Council of Research and Technology of Brazil (CNPq) Grant Number 471517/2012-7 to A. R. P., and by a Scholarship from CNPq (National Council for Scientific and Technological Development) of Brazil to J. F. T. We also thank the Embrapa Unit at Passo Fundo, RS, for support. Approved by the Publication Committee of the Embrapa Wheat, Passo Fundo, RS, Brazil under Number 21205.001716/2018-35.
Author information
Authors and Affiliations
Corresponding author
Additional information
Handling Editor: Heikki Hokkanen.
Rights and permissions
About this article

Cite this article
Tomacheski, J.F., Panizzi, A.R. & Lucini, T. Nymphal and adult biology of the red-shouldered stink bug, Thyanta perditor (F.), on cultivated and on wild plants. Arthropod-Plant Interactions 13, 91–98 (2019). https://doi.org/10.1007/s11829-018-9627-8
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s11829-018-9627-8