
Abstract
We analyzed the effect of a 6-week aerobic exercise training program on the in vivo macrophage reverse cholesterol transport (RCT) in human cholesteryl ester transfer protein (CETP) transgenic (CETP-tg) mice. Male CETP-tg mice were randomly assigned to a sedentary group or a carefully supervised exercise training group (treadmill 15 m/min, 30 min sessions, five sessions per week). The levels of plasma lipids were determined by enzymatic methods, and the lipoprotein profile was determined by fast protein liquid chromatography (FPLC). CETP activity was determined by measuring the transfer rate of 14C-cholesterol from HDL to apo-B containing lipoproteins, using plasma from CETP-tg mice as a source of CETP. The reverse cholesterol transport was determined in vivo by measuring the [3H]-cholesterol recovery in plasma and feces (24 and 48 h) and in the liver (48 h) following a peritoneal injection of [3H]-cholesterol labeled J774-macrophages into both sedentary and exercise trained mice. The protein levels of liver receptors were determined by immunoblot, and the mRNA levels for liver enzymes were measured using RT-PCR. Exercise training did not significantly affect the levels of plasma lipids or CETP activity. The HDL fraction assessed by FPLC was higher in exercise-trained compared to sedentary mice. In comparison to the sedentary group, a greater recovery of [3H]-cholesterol from the injected macrophages was found in the plasma, liver and feces of exercise-trained animals. The latter occurred even with a reduction in the liver CYP7A1 mRNA level in exercised trained animals. Exercise training increased the liver LDL receptor and ABCA-1 protein levels, although the SR-BI protein content was unchanged. The RCT benefit in CETP-tg mice elicited by exercise training helps to elucidate the role of exercise in the prevention of atherosclerosis in humans.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
Regular exercise is independently associated with a reduced incidence of coronary heart disease and mortality by improving the cardiovascular risk profile [1]. In fact, a negative correlation between physical activity level and the carotid intima media thickness has been shown in aerobically exercised individuals [2].
Ramachandran et al. [3] demonstrated that regular, aerobic exercise reduces pre-existing atherosclerotic lesions in low density lipoprotein (LDL) receptor knockout mice, and more recently, Matsumoto et al. [4] showed that in low-density lipoprotein-receptor-deficient mice, regular exercise training prevents aortic valve sclerosis by numerous mechanisms, including preservation of endothelial integrity, reduced inflammation and oxidative stress, and inhibition of the osteogenic pathway. It has also been reported that a progressive aerobic exercise training protocol improves the apo E knockout mouse survival rate, an effect that was ascribed to the stabilization of atherosclerotic lesions and prevention of plaque rupture [5]. These events are most likely related to the role of aerobic exercise in improving high density lipoprotein (HDL) plasma levels, as well as antioxidant and anti-inflammatory defenses in the arterial wall [6].
Through the reverse cholesterol transport (RCT) process, HDL mediates the cholesterol removal from macrophages and peripheral cells and its delivery to the liver, where it is secreted into the bile and feces. The cholesteryl ester transfer protein (CETP) transfers esterified cholesterol from HDL to apo B-containing lipoproteins (LP) that are ultimately removed by the hepatic LDL receptor (B-E), LRP and E receptors. There are no consistent data in the literature regarding the effect of aerobic exercise training on the RCT, and most of the results were drawn from in vitro analyses that may not reflect the whole animal system in vivo. In this regard, an interplay of actions involving CETP, lecithin cholesterol acyltransferase (LCAT), ABC transporters (ABCA-1 and ABCG-1) and scavenger receptor class B, type I (SR-BI) could dictate the flow of cholesterol to the liver [7].
In the present study, we tested the hypothesis that aerobic exercise training can improve the in vivo RCT by modulating proteins and enzymes involved in the cholesterol flow to the liver. For this purpose, we used CETP transgenic C57BL/6 mice (CETP-tg), an animal model that simulates the RCT in humans. We assessed the effect of a 6 week aerobic training program on the redistribution of radiolabeled cholesterol into the plasma, liver and feces following the intraperitoneal injection of macrophages enriched with [3H] cholesterol. We also investigated the hepatic protein levels of receptors involved in lipid metabolism, such as SR-BI, ATP-binding cassette transporter A-1 (ABCA-1), liver X receptor (LXR) and LDL receptor, and the mRNA of enzymes that convert cholesterol in bile acids (CYP7A1 and CYP27A). We found that aerobic exercise training improves the macrophage RCT. Exercise-trained mice had higher levels of macrophage-derived 3H-cholesterol in their plasma, liver and feces. The enhanced protein levels of hepatic ABCA-1 and LDL receptor contributed to greater HDL plasma concentration and cholesterol flow into the liver, respectively. The RCT benefit seen in exercise-trained CETP-tg mice helps to elucidate the role of exercise in the prevention of atherosclerosis in humans.
Materials and Methods
Animals and Aerobic Exercise Protocol
Two-month-old male transgenic mice homozygous for human CETP (human natural promoter-driven CETP transgenic mice, CETP-tg; line 5203; back-crossed on a C57BL/6 background for 10 generations) that originated in Dr. AR Tall′s laboratory and were kindly provided by Dr. HCF Oliveira (University of Campinas, São Paulo, Brazil) were fed a pelleted commercial chow ad libitum (Nuvilab-Nuvital, São Paulo, Brazil) with free access to water. Animals were housed in conventional housing at 22 ± 2 °C with a 12 h light/dark cycle. Protocols were approved by the Institutional Animal Care and Research Advisory Committee (Hospital das Clinicas of the Faculty of Medical Sciences, University of São Paulo-CAPPesq # 773/06) according to the US National Institutes of Health Guidelines. Animal care was provided in accordance with the procedures outlined in the Guide for the Care and Use of Laboratory Animals [8], and experiments were performed under adherence to the American College of Sports Medicine (ACSM) animal care standards. CETP-tg animals were subjected to a 6 week monitored aerobic exercise training protocol performed on a treadmill (WEG CFW-08, São Carlos, Brazil) at 15 m/min during 30 min sessions five times a week. A control group was kept sedentary for the same amount of time. Exercise sessions were carried out in the late afternoon and were supervised by the same investigator (DDFM Rocco).
LDL Isolation and Acetylation
Procedures with humans conformed to the Declaration of Helsinki and all participants signed an informed written consent form previously approved by The Ethical Committee for Human Research Protocols of the Hospital of the Faculty of Medical Sciences, University of São Paulo (CAPPesq # 773/06). Low density lipoprotein (LDL) was isolated from healthy plasma donors by sequential ultracentrifugation (d = 1.019−1.063 g/mL) and was further purified by discontinuous gradient ultracentrifugation. After dialysis against PBS, the protein content was determined using the Lowry procedure [9], and acetylation was performed with acetic anhydride according to Basu et al. [10]. After extensive dialysis against ethylene diamine tetra acetic phosphate-buffered saline (EDTA–PBS), acetylated LDL (AcLDL) was kept sterile at 4 °C under nitrogen atmosphere and was used within a month.
Measurement of the In Vivo RCT
J-774 macrophages were grown in an RPMI 1640 culture medium supplemented with 10% fetal bovine serum (FBS), labeled with 5 μ Ci/mL [3H]-cholesterol and enriched with 50 μg/mL of AcLDL for 48 h, as previously described [11]. Cells were then removed from plaques by trypsin treatment followed by inactivation of the enzyme by adding RPMI plus FBS. The cells were then spun down by centrifugation at 4 °C at 1,500 rpm and rediluted in a small volume of PBS. Cell viability, assessed by exclusion with Trypan blue, was greater than 98%, and an aliquot was placed in a beta counter (Beckman model; Palo Alto, USA) to determine the radioactivity. The intracellular distribution of cholesterol fractions was determined by thin-layer chromatography and consisted of 96 and 4% in the free and esterified forms, respectively.
Forty-eight hours after the last bout of exercise, [3H]-cholesterol-labeled J-774 foam cells (3.5 × 106 cells; ~80,000 dpm in 100 μL of PBS) were intraperitoneally injected into the exercise-trained and sedentary mice, which were individually housed in metabolic cages with free access to food and water. Samples of blood from the tail vein and feces were taken after 24 and 48 h. Plasma radioactivity was immediately determined after spinning the blood at 1,500 rpm for 20 min at 4 °C. The mice were euthanized 48 h after the injection. Their organs (liver, spleen, lung, heart, kidneys and adrenal glands) were subsequently removed, carefully washed with cold 0.9% NaCl solution, gently dried with filter paper and weighed.
The feces and organs were stored at −70 °C until further analyses. Total feces and organs or liver aliquots were homogenized with a 2:1 (v/v) mixture of chloroform/methanol [12] and stored at 4 °C overnight for lipid extraction. The lipid layer was drawn and evaporated under nitrogen flow, and the radioactivity was determined. Results are expressed as percentages of total dose (dpm) injected per gram of sample (liver or feces) or plasma volume (mL). The radioactivity recovery in heart, lung, spleen, kidney and adrenal glands was too low and represented less than 1% of the total radioactivity injected (data not shown).
Measurement of Hepatic Cholesterol Content
Lipids were extracted from the liver, and total cholesterol was determined from a supernatant aliquot using the enzymatic method (Roche Diagnóstica, SP, Brazil), and the protein concentration by using the Lowry technique [9].
Plasma Lipid Analysis
Plasma (100 μL) from each mouse was analyzed by fast protein liquid chromatography (FPLC) gel filtration on two Superose-6 columns. The cholesterol concentration in all LP fractions was measured with a Cobas Mira using an enzymatic colorimetric kit (Roche Diagnostic, Brazil).
CETP Activity Determination
CETP activity was measured as the transfer of [14C]-cholesteryl oleate from HDL to VLDL and LDL using LP isolated from pools of human normolipidemic individuals and plasma from CETP-tg mice as a source of CETP as previously described [13].
Western Blotting Analysis
Protein lysates were obtained from frozen hepatic tissue from exercise-trained and sedentary CETP-tg mice. Tissue was homogenized in a Polytron (MA099 Potter Unit, Marcone Equip., Sao Paulo, Brazil) with a buffer containing 20 mM Hepes, 150 mM NaCl, 10% glycerol, 1% triton, 1 mM EDTA, 1.5 mM MgCl2 and protease inhibitors. The supernatant was obtained after centrifugation, and an aliquot was dissolved in SDS-glycerol. Equal amounts of sample protein were applied to a polyacrylamide gel. SR-BI, ABCA-1, LXR and the LDL receptor protein levels were determined using anti-SR-BI (1:1,000), anti-ABCA-1 (1:1,000), anti-LXRα/β (1:1,000) (Novus Biologicals, Inc., Littleton, CO, USA), and anti-LDL receptor (M-20):sc-11826, (1:1,000) (Santa Cruz Biotechnology Inc, USA) followed by incubation with HRP-conjugated antibody and ECL reaction (Super Signal West Pico Chemiluminescent substrate, Pierce, Rockford, IL, EUA). Protein membrane stripping was performed by rinsing with 0.8 mM NaOH. The difference between the bands was analyzed in pixels using the JX-330 Color Image Scanner (Sharp®) and ImageMaster software (Pharmacia Biotech). The results are expressed as arbitrary units. β-actin protein levels (anti β-actin 1:1,000, Fitzgerald Industries International, Inc., Concord, MA) were used as a control, and Ponceau staining of nitrocellulose membranes was implemented to assure equal protein loading.
CYP7A1 and CYP27A mRNA Expression in the Liver
At the final period of the study, the liver was removed and immediately frozen in liquid nitrogen. mRNA from liver samples (~50 mg) was extracted with 1 mL of Trizol (Invitrogen Life Technologies, Carlsbad, CA, USA). Reverse transcription reactions were performed with 2 μg of RNA using TaqMan Reverse Transcription Reagents (Applied Biosystems, Foster City, CA). Real time-PCR was performed on an Mx3000P QPCR System (Stratagene, La Jolla, CA) using the Brilliant SYBR Green QPCR Master Mix (Stratagene, catalogue no. 600548). The following primers were used: β actin sense 5′-TGGAGAGCACCAAGACAGACA-3′ and antisense 5′-TGCCGGAGTCGACAATGAT-3′; CYP7A1 sense 5′-AGCAACTAAACAACCTGCCAGTACTA-3′ and antisense 5′-GTCCGGATATTCAAGGATGCA-3′; CYP27A sense 5′-GGAGGGCAAGTACCCAATAAGA-3′ and antisense 5′-TGCGATGAAGATCCCATAGGT-3′.
Determination of Apo A-I in the Peritoneal Fluid
Mouse peritoneal cavities were rinsed with PBS, and the fluid was collected in order to measure the apo A-I content. The sample volume was reduced using Amicon filters (Millipore, USA), and the same amount of protein was submitted to immunoblot analysis as described above using anti apo A-I Ab 1:1,000 (Santa Cruz Diag, USA).
Statistical Analysis
Statistical analyses were performed using GraphPad Prism 4.0 software (GraphPad Prism, Inc., San Diego, CA). A non-paired Student’s t test was used to compare differences between groups. Summary data are reported as mean values ± standard error or mean values ± standard deviation as indicated. A p value <0.05 was considered statistically significant.
Results
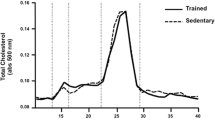
CETP-tg C57BL/6 male mice were submitted to a 6 week period of supervised aerobic exercise training. Body weight, total plasma cholesterol and triacylglycerols were not modified by exercise training (Table 1). As shown in Fig. 1, the VLDL-cholesterol and LDL-cholesterol plasma profile was not modified by the aerobic exercise training, although the HDL-cholesterol fraction área was higher in the trained animals compared to the sedentary animals in the final period of the protocol. Plasma CETP activity was similar in sedentary and trained mice at the beginning of the protocol and did not change after the exercise training period. Moreover, CETP plasma activity was not different between the sedentary and trained groups when the initial and final periods were compared (Table 1).

Plasma lipoprotein profile of aerobically trained and sedentary CETP-tg mice. Plasma lipoproteins were isolated by fast protein liquid chromatography (FPLC), and total cholesterol was determined in all fractions using an enzymatic colorimetric kit. Sedentary (dashed line) and trained (filled line) CETP-tg mice
The [3H]-cholesterol content was analyzed in plasma and feces at 24 and 48 h and in the liver at 48 h following the intraperitoneal injection of J774 macrophages enriched with AcLDL and radiolabeled cholesterol. A higher percentage of [3H]-cholesterol was recovered from the plasma (Fig. 2a), liver (Fig. 2b) and feces (Fig. 2c) of the trained mice when compared to the sedentary animals.

Macrophage-derived [3H]-cholesterol recovery in a plasma, b liver and c feces from aerobically trained and sedentary CETP-tg mice. J774 macrophages enriched with acetylated LDL and [3H]-cholesterol were injected into the peritoneal cavities of sedentary and trained CETP-tg mice. a Plasma and c feces were collected after 24 and 48 h of the injection, and b the liver was removed after 48 h. After lipid extraction, the radioactivity was determined, and the recovery of [3H]-cholesterol is expressed as the percentage of injected dose/mL of plasma or percentage of injected dose/mg of tissue or feces. Data are expressed as mean values ± standard error
As compared to the sedentary group, the increase in radiolabeled cholesterol in the liver of exercise-trained mice was accompanied by a 50% increase in protein levels of the hepatic LDL receptor (Fig. 3). In contrast, exercise training did not raise the SR-BI hepatic level (Fig. 3a). No changes were observed in the protein levels of LXR in the liver of exercise-trained animals (Fig. 3a). The hepatic ABCA-1 protein level was increased 100% in exercise-trained animals (Fig. 3a). CYP7A1 mRNA levels were reduced in exercise-trained CETP-tg mice, whereas their CYP27A mRNA levels were not influenced by exercise training (Fig. 3b). Total cholesterol content in the liver (mg of cholesterol/g of organ; mean ± SE) was not different between trained (43.3 ± 9.9; n = 6) and sedentary (40.4 ± 1.38; n = 6) animals.

LDL receptor, scavenger receptor class B type 1 (SR-BI), liver X receptor (LXR) and ATP binding cassette transporter (ABCA-1) protein levels and CYP7A1 and CYP27A mRNA in the livers of aerobically trained and sedentary CETP-tg mice. a liver samples from aerobically trained and sedentary CETP-tg mice were homogenized and dissolved in SDS-buffer. Equal amounts of protein lysates were applied to a 10% polyacrylamide gel and submitted to electrophoresis and immunoblotting with anti-LDL receptor Ab (1:1,000), anti-SR-BI Ab (1:1,000), anti-LXR Ab (1:1,000) and anti-ABCA-1 Ab (1:1,000). Following incubation with secondary Ab conjugated with HRP, bands were visualized after ECL reaction. Each lane represents one animal sample. Data are expressed as mean values ± standard deviation. b CYP7A1 and CYP27A mRNA was determined by RT-PCR as described in Material and Methods
The apo A-I content in peritoneal fluid as assessed by immunoblot was similar before and after exercise training (Fig. 4). Thus, the increase in the RCT after exercise training could not be ascribed to changes in apo A-I content.

Content of apo A-I in peritoneal fluid from aerobically trained and sedentary CETP-tg mice. Mouse peritoneal cavities were rinsed with PBS, and the fluid was collected. Protein was submitted to immunoblot analysis with anti apo A-I Ab 1:1,000
Discussion
The importance of regular exercise for the prevention and treatment of chronic diseases is widely acknowledged [1]. Aerobic exercise improves LP metabolism and prevents the development of cardiovascular disease. Apart from the well-documented favorable effects of regular exercise on cardiovascular risk factors, it has been suggested that physical training may involve additional mechanisms that ameliorate atherosclerosis.
HDL is known for its anti-atherogenic properties that rely on its ability to support the cholesterol efflux from cholesterol-loaded arterial macrophages. Previous investigations focused on the benefits of exercise on lipoprotein lipase activity, including the mediation of triacylglycerol-rich LP metabolism and the increased levels of plasma HDL cholesterol [14, 15]. Nonetheless, it is unclear whether exercise plays a role in the RCT system in vivo. In this study, we used an experimental model designed to evaluate in vivo RCT and found that a 6 week carefully controlled aerobic exercise training program accelerates the RCT in CETP-tg mice.
CETP-tg mice are a useful tool for investigating the contribution of CETP to RCT. CETP is known to play a major role in RCT in humans. In huCETP-tg mice, the uptake of apo B-containing LP by the LDL receptor offers an additional route for the delivery of plasma esterified cholesterol to the liver. Accordingly, the increased RCT observed in the exercise-trained mice in this study may be ascribed to the enhanced protein levels of the LDL receptor in the liver. Elevated levels of hepatic LDL receptor mRNA have previously been shown in aerobic trained mice [16, 17], although the protein content has never been reported. In our investigation, exercise in the presence of CETP increased the uptake of LDL cholesterol by the liver and ABCA-1 protein levels, which may have prevented to liver cholesterol content from increasing by enhancing the export of cholesterol. In addition, by increasing the HDLc fraction as assessed by FPLC, exercise training may have contributed to a greater removal of cell cholesterol, despite a slight reduction in CYP7A1 mRNA levels.
SR-BI plays a major role in the final step of the RCT system. Although macrophage SR-BI could not be related to an improvement in RCT efficiency [18], SR-BI protein levels in the liver are a positive regulator of macrophage RCT [19]. In fact, hepatic SR-BI is directly related to the secretion of cholesterol into the bile and to the reduction of atherosclerosis in animal models with low levels of HDL in plasma [20, 21]. Wei et al. [16] have described an enhancement of the hepatic SR-BI mRNA levels in wild-type C57BL/6 mice submitted to a 2 week aerobic training program. Surprisingly, the liver SR-BI protein levels did not change after aerobic exercise training in our study. This is likely due to a greater cholesterol influx mediated by the LDL receptor.
In huCETP-tg mice, a direct role of CETP in accelerating the tissue uptake of esterified cholesterol (EC) was demonstrated [22, 23]. Although improved HDL-EC removal and increased cholesterol content in the liver have been seen in CETP-tg mice, the excretion of biliary lipids and fecal bile acid have not been consistently found in these animals when compared to wild-type mice [24]. In CETP-tg mice, the in vivo RCT from macrophage to feces was not modified [25]. Nonetheless, the hypothesis that CETP facilitates RCT is strengthened by evidences related to mice expressing CETP alone [26] or combined with knock-outs of LDL or SR-BI receptors. In these animals, CETP elicited macrophage RCT despite a reduction in HDL [27], and SR-BI-deficient mice were protected against diet-induced atherosclerosis [28]. On the other hand, the treatment of hamsters with torcetrapib, a drug that suppresses the CETP activity, increased the appearance of cholesterol originating from peritoneally injected macrophages in plasma and feces [26].
Increased levels of CYP27A mRNA have been described by Wilund et al. [17] in mice submitted to a 12 week aerobic exercise training program. In addition, voluntary wheel running increased fecal bile acid and cholesterol output and reduced intestinal cholesterol absorption in mice [29]. In our study, the CYP7A1 mRNA levels were reduced but CYP27A mRNA was not modified by exercise training, suggesting that the greater amount of radiolabeled cholesterol found in feces could be attributed to the increased excretion of cholesterol instead of bile acids. A recent paper from Yasuda et al. [30] showed that the activation of RCT resulted in a higher excretion of cholesterol, but not bile acids, in the feces of LXR agonist-treated mice. A limitation of our study was the fact that we did not measure the intestinal cholesterol absorption that could affect RCT in vivo as occurred in mice treated with ezetimibe [31]. Also, Temel et al. [32] used genetic and surgical models of biliary cholesterol insufficiency to demonstrate that a non-classical hepatobiliary pathway can contribute to RCT independently of biliary sterol secretion.
While this manuscript was under preparation, Meissner et al. [33] published a paper showing that the in vivo RCT, assessed by the same methodology used here, was not modified by voluntary wheel running in wild-type C57BL-6 mice compared to sedentary animals. Differences between their report and our results are likely ascribed to: (1) the presence of CETP in our model and its role in the alternative pathway of cholesterol disposal via the LDL receptor; (2) their results were obtained in free running wheel mice, which cannot assure equal training intensities along the protocol period, as opposed to the regular and carefully supervised exercise training protocol that was utilized in our study.
Although CETP activity was not changed after exercise training in our study, we should bear in mind that the CETP-mediated HDL-EC uptake by the liver is independent of the plasma activity responsible for the exchange of EC and TAG between LP [22]. Nonetheless, divergent data have been published regarding the role of CETP inhibitors like torcetrapib on this process [26]. Zhou et al. [34] suggested that the hepatic acquisition of EC from HDL mediated by CETP occurs independently of SR-BI, LDL-receptor related protein (LRP) and possibly the LDL receptor.
Controversial results have been published regarding the effect of exercise on the CETP mass and activity, which could be ascribed to the different methodologies used to measure CETP, as well as to variations in exercise intensity and frequency and the measurement of acute or chronic effects of exercise [14, 15]. Moreover, the efficiency of aerobic exercise in modulating the lipid profile is related to gender, age, insulin resistance, weight loss and genetic polymorphisms of CETP, apo E and apo A-I [36–38].
Exercise training did not modify the protein levels of LXR in CETP-tg mice. On the other hand, ABCA-1 was greatly enhanced following training, and this could have contributed to the higher levels of HDLc observed in exercise-trained CETP-tg mice in the final period of the study. ABCA-1 in the liver is known to contribute to HDL formation and plasma HDL cholesterol levels. Adenovirus-targeted protein levels of ABCA-1 in the liver increased HDL plasma levels in mice [39, 40]. On the other hand, an 80% reduction in plasma HDL cholesterol levels was observed in mice with a targeted inactivation of hepatic ABCA-1 [41], and ABCA-1 has been shown to play a critical role in HDL catabolism by the liver [42].
Systemic and macrophage ABCA-1 are positive regulators of macrophage RCT in vivo [18, 35]. This is consistent with our findings. An enhancement in ABCA-1 mRNA levels in the livers isolated from exercise-trained rats has also been described [43]. Furthermore, Butcher et al. [44] reported an increased expression of ABCA-1 and ABCG-1 mRNA in leukocytes isolated from trained subjects compared to sedentary ones.
In conclusion, aerobic exercise training improves RCT from macrophages in CETP-tg mice by increasing the concentration of HDL in plasma and by distinctly modulating the protein levels of liver receptors involved in the uptake of EC, which may contribute to the prevention and regression of atherosclerosis as previously reported by others [3, 5, 45]. With regards to the role of CETP in lipid metabolism in humans, the findings of our study shed light on an important effect of exercise on the RCT system. Taking into account that exercise has many effects on lipid and LP metabolism, the exact mechanisms by which exercise favorably modulates the lipid transport should be further investigated.
Abbreviations
- ABC:
-
Transporters ABCA-1 and ABCG-1
- AcLDL:
-
Acetylated LDL
- Apo AI:
-
Apolipoprotein AI
- Apo E:
-
Apolipoprotein E
- B-E:
-
Low density lipoprotein receptor
- CETP:
-
Cholesteryl ester transfer protein
- CYP7A1:
-
7 alpha hydroxylase
- CYP27A:
-
27 alpha hydroxylase
- EC:
-
Esterified cholesterol
- EDTA-PBS:
-
Ethylene diamine tetra acetic phosphate-buffered saline
- FPLC:
-
Fast protein liquid chromatography
- HDL:
-
High density lipoprotein
- LCAT:
-
Lecithin cholesterol acyltransferase
- LDL:
-
Low density lipoprotein
- LRP:
-
LDL-receptor related protein
- LXR:
-
Liver X receptor
- LP:
-
Lipoprotein
- RCT:
-
Reverse cholesterol transport
- SR-BI:
-
Scavenger receptor class B type I
- VLDL:
-
Very low density lipoprotein
References
Lee IM, Hsieh CC, Paffenbarger RS Jr (1995) Exercise intensity and longevity in men. The Harvard Alumni Study. JAMA 273:1179–1184
Nordstrom CK, Dwyer KM, Merz NB, Shircore A, Dwyer JH (2003) Leisure time physical activity and early atherosclerosis: the Los Angeles atherosclerosis study. Am J Med 115:19–25
Ramachandran S, Penumetcha M, Merchant NK, Santanam N, Rong R, Parthasarathy S (2005) Exercise reduces preexisting atherosclerotic lesions in LDL receptor knock out mice. Atherosclerosis 178:33–38
Matsumoto Y, Adams V, Jacob S, Mangner N, Schuler G, Linke A (2010) Regular exercise training prevents aortic valve disease in low-density lipoprotein-receptor-deficient mice. Circulation 21:759–767
Napoli C, Williams-Ignarro S, Nigris F, Lerman LO, D’Armiento FP, Crimi E, Byrns RE, Casamassimi A, Lanza A, Gombos F, Sica V (2006) Physical training and metabolic supplementation reduce spontaneous atherosclerotic plaque rupture and prolong survival in hypercholesterolemic mice. PNAS 13:10479–10484
Meilhac O, Ramachandran S, Chiang K, Santanam N, Parthasarathy S (2001) Role of arterial wall antioxidant defense in beneficial effects of exercise on atherosclerosis in mice. Arterioscler Thromb Vasc Biol 21:1681–1688
Wang X, Rader DJ (2007) Molecular regulation of macrophage reverse cholesterol transport. Curr Opin Cardiol 22:368–372
Institute of Laboratory Animal Research CoLS (1996) National Research Council guide for the care and use of laboratory animals. In: Council NR (ed). National Academy Press, Washington (DC), p 124
Lowry OH, Rosebrough NJ, Farr Al, Randall RJ (1951) Protein measurement with folin phenol reagent. J Biol Chem 193:265–275
Basu SK, Goldstein JL, Anderson RGW, Brown MS (1976) Degradation of cationized low density lipoprotein and regulation of cholesterol metabolism in homozygous hypercholesterolemia fibroblasts. Proc Natl Acad Sci 73:3178–3182
Zhang Y, Zanotti I, Reilly MP, Glick JM, Rothblat GH, Rader DJ (2003) Overexpression of apolipoprotein A-1 promotes reverse transport of cholesterol from macrophages to feces in vivo. Circulation 108:661–663
Folch J, Lees M, Stanley S (1957) A simple method for the isolation and purification of total lipids from animal’s tissues. J Biol Chem 226:497–509
Escolà-Gil JC, Julve J, Marzal-Casacuberta A, Ordóñez-Llanos J, González-Sastre F, Blanco-Vaca F (2001) ApoA-II expression in CETP transgenic mice increases VLDL production and impairs VLDL clearance. J Lipid Res 42:241–248
Gupta AK, Ross EA, Myers JN, Kashyap ML (1993) Increased reverse cholesterol transport in athletes. Metabolism 42:684–690
Olchawa B, Kingwell BA, Hoang A, Schneider L, Miyazaki O, Nestel P, Sviridov D (2004) Physical fitness and reverse cholesterol transport. Arterioscler Thromb Vasc Biol 24:1087–1091
Wei C, Penumetcha M, Santanam N, Liu GY, Garelnabi M, Parthasarathy S (2005) Exercise might favor reverse cholesterol transport and lipoprotein clearance: potential mechanism for its anti-atherosclerotic effects. Biochim Biophys Acta 1723:124–127
Wilund KR, Feeney LA, Tomayko EJ, Chung HR, Kim K (2008) Endurance exercise training reduces gallstone development in mice. J Appl Physiol 104:761–765
Wang X, Collins HL, Ranalletta M, Fuki IV, Billheimer JT, Rothblat GH, Tall AR, Rader DJ (2007) Macrophage ABCA1 and ABCG1, but not SR-BI, promote reverse cholesterol transport in vivo. J Clin Invest 117:2216–2224
Zhang Y, Da Silva JR, Reilly M, Billheimer JT, Rothblat GH, Rader DJ (2005) Hepatic expression of scavenger receptor class B type I (SR-BI) is a positive regulator of macrophage reverse cholesterol transport in vivo. J Clin Invest 115:2870–2874
Mardones P, Quiñones V, Amigo L, Moreno M, Miquel JF, Schwarz M, Miettinen HE, Trigatti B, Krieger M, VanPatten S, Cohen DE, Rigotti A (2001) Hepatic cholesterol and bile acid metabolism and intestinal cholesterol absorption in scavenger receptor class B type I-deficient mice. J Lipid Res 42:170–180
Kozarsky KF, Donahee MH, Glick JM, Krieger M, Rader DJ (2000) Gene transfer and hepatic overexpression of the HDL receptor SR-BI reduces atherosclerosis in the cholesterol-fed LDL receptor-deficient mouse. Arterioscler Thromb Vasc Biol 20:721–727
Gauthier A, Lau P, Zha X, Milne R, McPherson R (2005) Cholesteryl ester transfer protein directly mediates selective uptake of high density lipoprotein cholesteryl esters by the liver. Arterioscler Thromb Vasc Biol 25:2177–2184
Vassiliou G, McPherson R (2004) Role of cholesteryl ester transfer protein in selective uptake of high density lipoprotein cholesteryl esters by adipocytes. J Lipid Res 5:1683–1693
Harada LM, Amigo L, Cazita PM, Salerno AG, Rigotti AA, Quintão EC, Oliveira HC (2007) CETP expression enhances liver HDL-cholesteryl ester uptake but does not alter VLDL and biliary lipid secretion. Atherosclerosis 191:313–318
Rotllan N, Calpe-Berdiel L, Guillaumet-Adkins A, Süren-Castillo S, Blanco-Vaca F, Escolà-Gil JC (2008) CETP activity variation in mice does not affect two major HDL antiatherogenic properties: macrophage-specific reverse cholesterol transport and LDL antioxidant protection. Atherosclerosis 196:505–513
Tchoua U, D'Souza W, Mukhamedova N, Blum D, Niesor E, Mizrahi J, Maugeais C, Sviridov D (2008) The effect of cholesteryl ester transfer protein overexpression and inhibition on reverse cholesterol transport. Cardiovascular Res 77:732–739
Tanigawa H, Billheimer JT, Tohyama J, Zhang Y, Rothblat G, Rader DJ (2007) Expression of cholesteryl ester transfer protein in mice promotes macrophage reverse cholesterol transport. Circulation 116:1267–1273
Harder C, Lau P, Meng A, Whitman SC, McPherson R (2007) Cholesteryl ester transfer protein (CETP) expression protects against diet induced atherosclerosis in SR-BI deficient mice. Arterioscler Thromb Vasc Biol 27:858–864
Meissner M, Havinga R, Boverhof R, Kema I, Groen AK, Kuipers F (2010) Exercise enhances whole-body cholesterol turnover in mice. Med Sci Sports Exerc 42:1460–1468
Yasuda T, Grillot D, Bilheimer JT, Briand F, Delerive P, Huet S, Rader DJ (2010) Tissue-specific liver X receptor activation promotes macrophage reverse cholesterol transport in vivo. Arterioscler Thromb Vasc Biol 30(4):781–786
Sehayek E, Hazen SL (2008) Cholesterol absorption from the intestine is a major determinant of reverse cholesterol transport from peripheral tissue macrophages. Arterioscler Thromb Vasc Biol 28:1296–1297
Temel RE, Sawyer JK, Yu L, Lord C, Degirolamo C, McDaniel A, Marshall S, Wang N, Shah R, Rudel LL, Brown JM (2010) Biliary sterol secretion is not required for macrophage reverse cholesterol transport. Cell Metab 12:96–102
Meissner M, Nijstad N, Kuipers F, Tietge UJ (2010) Voluntary exercise increases cholesterol efflux but not macrophage reverse cholesterol transport in vivo in mice. Nutr Metab 1:54
Zhou H, Li Z, Silver DL, Jiang XC (2006) Cholesteryl ester transfer protein (CETP) expression enhances HDL cholesteryl ester liver delivery, which is independently of scavenger receptor BI, LDL receptor related protein and possibly LDL receptor. Biochim Biophys Acta 1761:1482–1488
Calpe-Berdiel L, Rotllan N, Palomer X, Ribas V, Blanco-Vaca F, Escolà-Gil JC (2005) Direct evidence in vivo of impaired macrophage-specific reverse cholesterol transport in ATP-binding cassette transporter A1-deficient mice. Biochim Biophys Acta 1738:6–9
Couillard C, Després JP, Lamarche B, Bergeron J, Gagnon J, Leon AS, Rao DC, Skinner JS, Wilmore JH, Bouchard C (2001) Effects of endurance exercise training on plasma HDL cholesterol levels depend on levels of triglycerides: evidence from men of the Health, Risk Factors, Exercise Training and Genetics (HERITAGE) Family Study. Arterioscler Thromb Vasc Biol 21:1226–1232
Mukherjee M, Shetty KR (2004) Variations in high-density lipoprotein cholesterol in relation to physical activity and Taq 1B polymorphism of the cholesteryl ester transfer protein gene. Clin Genet 65:412–418
Sorlí JV, Corella D, Francés F, Ramírez JB, González JI, Guillén M, Portolés O (2006) The effect of the APOE polymorphism on HDL-C concentrations depends on the cholesterol ester transfer protein gene variation in a Southern European population. Clin Chim Acta 366:196–203
Basso F, Freeman L, Knapper CL, Remaley A, Stonik J, Neufeld EB, Tansey T, Amar MJ, Fruchart-Najib J, Duverger N, Santamarina-Fojo S, Brewer HB Jr (2003) J Role of the hepatic ABCA1 transporter in modulating intrahepatic cholesterol and plasma HDL cholesterol concentrations. J Lipid Res 44(2):296–302
Wellington CL, Brunham LR, Zhou S, Singaraja RR, Visscher H, Gelfer A, Ross C, James E, Liu G, Huber MT, Yang YZ, Parks RJ, Groen A, Fruchart-Najib J, Hayden MR (2003) Alterations of plasma lipids in mice via adenoviral-mediated hepatic overexpression of human ABCA1. J Lipid Res 44(2):1470–1480
Timmins JM, Lee JY, Boudyguina E, Kluckman KD, Brunham LR, Mulya A, Gebre AK, Coutinho JM, Colvin PL, Smith TL, Hayden MR, Maeda N, Parks JS (2005) Targeted inactivation of hepatic Abca1 causes profound hypoalphalipoproteinemia and kidney hypercatabolism of apoA-I. J Clin Invest 115(5):1333–1342
Singaraja RR, Stahmer B, Brundert M, Merkel M, Heeren J, Bissada N, Kang M, Timmins JM, Ramakrishnan R, Parks JS, Hayden MR, Rinninger F (2006) Hepatic ATP-binding cassette transporter A1 is a key molecule in high-density proprotein cholesteryl ester metabolism in mice. Arterioscler Thromb Vasc Biol 26(8):1821–1827
Ghanbari-Niaki A, Khabazian BM, Hossaini-Kakhak SA, Rahbarizadeh F, Hedayati M (2007) Treadmill exercise enhances ABCA1 expression in rat liver. Biochem Biophys Res Commun 361:841–846
Butcher LR, Thomas A, Backx K, Roberts A, Webb R, Morris K (2008) Low-intensity exercise exerts beneficial effects on plasma lipids via PPARgamma. Med Sci Sports Exerc 40:1263–1270
Okabe TA, Shimada K, Hattori M, Murayama T, Yokode M, Kita T, Kishimoto C (2007) Swimming reduces the severity of atherosclerosis in apolipoprotein E deficient mice by antioxidant effects. Cardiovasc Res 74:537–545
Acknowledgments
This work was supported by Fundação de Amparo à Pesquisa do Estado de Sao Paulo—FAPESP [07/50387-8 to MP, 06/52702-5 to DDFM Rocco, 07/56654-8 to LS Okuda and 09/53412-9 to RS Pinto] and by Conselho Nacional de Desenvolvimento Científico e Tecnológico-CNPq [870248/1197-9 to DDFMR]. The authors are indebted to Dr. Marisa Dolhnikoff and Ana L B Silveira from the Pathology Museum, Faculty of Medical Sciences, University of Sao Paulo for helping with microscope analysis, and to Walter Campestre (LIM-16) and Antonio dos Santos Filho (LIM-17) for caring for the animals. The authors are thankful to Fundação Faculdade de Medicina and Laboratórios de Investigação Médica (LIM). Results of the present study do not constitute endorsement by ACSM.
Conflict of interest
None.
Author information
Authors and Affiliations
Corresponding author
About this article
Cite this article
Rocco, D.D.F.M., Okuda, L.S., Pinto, R.S. et al. Aerobic Exercise Improves Reverse Cholesterol Transport in Cholesteryl Ester Transfer Protein Transgenic Mice. Lipids 46, 617–625 (2011). https://doi.org/10.1007/s11745-011-3555-z
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s11745-011-3555-z