
Abstract
The halophyte Puccinellia tenuiflora efficiently restricts Na+ entry into roots and maintains a strong selective absorption (SA) capacity for K+ over Na+ to cope with salt stress. AKT1-type channel, an important plant K+ uptake protein, may be active in regulating SA in P. tenuiflora. Here, its encoding gene PtAKT1 was isolated from P. tenuiflora. To examine the function of PtAKT1 in salt resistance and selectivity for K+ over Na+, we analyzed the transcript patterns of PtAKT1 and Na+, K+ concentration in P. tenuiflora during varying concentrations of NaCl and KCl. Results displayed that both the transcription abundance of PtAKT1 in roots and the whole plant K+ concentration did not change with varying K+ concentrations in the medium (0-10 mM). When NaCl (25 or 150 mM) was present, transcript levels of PtAKT1 were down-regulated, accompanied by a decrease in the whole plant K+ concentration. However, under normal (5 mM) or low (0.1 mM) KCl condition, transcript levels of PtAKT1 in roots under severe salt stress (150 mM NaCl) were higher than that in mild salt condition (25 mM NaCl) within 6–24 h, so did SA during the whole treatment time (6–96 h). A strong positive correlation was existed between root PtAKT1 transcription abundance and SA values during 25 or 150 mM NaCl plus KCl. Consequently, it is proposed that PtAKT1 is a key factor in regulating selective absorption capacity for K+ over Na+ in P. tenuiflora during saline condition. Finally, we hypothesize a model that SOS1, HKT and SKOR synergistically regulate potassium and sodium transport systems in the xylem parenchyma, and subsequently modulates K+ absorption via AKT1, which is located in epidermis, cortex and endodermis. The model could also provide a likely evidence to the mechanism that SOS1 could protect AKT1 mediating K+ influx during salt stress.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
Salinity is a main abiotic factor limiting crop production worldwide (Flowers 2004). Excessive accumulation of Na+ in plants can damage cell membrane and inhibit enzyme activity of metabolism. Owing to the similar ionic hydrated radius between Na+ and K+, Na+ always compete the binding site of K+ channels and transporters, thus restricts K+ acquisition and induces imbalance of cytoplasmic cation, affecting many physiological and metabolic activities accordingly, for instance, enzyme system homeostasis and osmatic adjustment (Ding and Zhu 1997; Buschmann et al. 2000; Shabala and Cuin 2008; Horie et al. 2009). Maintaining a high K+/Na+ selectivity is important in most plants.
Puccinellia tenuiflora is a typical gramineous halophyte, which is broadly distributed in saline swamplands, with significant feeding and ecological values (Wang et al. 2009; Guo et al. 2012). Our previous researches proved that limiting Na+ flow into roots and possessing a powerful selectivity for K+ contributed to the salt resistance of P. tenuiflora (Wang et al. 2009; Guo et al. 2012). Recent research found that plasma membrane Na+/H+ antiporter PtSOS1 was a key factor in selective transport capacity for K+ over Na+ in P. tenuiflora in salt condition (Guo et al. 2012). However, the molecular mechanism of selective absorption (SA) capacity for K+ over Na+ in P. tenuiflora remains unclear. The research by Peng et al. (2004) demonstrated the ability of P. tenuiflora to absorb K+ under salt stress depended on K+ channels that inhibited by Cs+. A further study by Ardie et al. (2010) found that over-expression of PtAKT1 led to the rise of K+ content and decrease in Na+ accumulation in Arabidopsis thaliana. These researches indicate that AKT1-type channel is closely related to K+ uptake in roots and salt tolerance in P. tenuiflora.
AKT1 (ARABIDOPSIS K+ TRANSPORTER1), a Shaker family K+ channel, is crucial in plant potassium nutrition acquisition. In A. thaliana, most of the potassium absorbs through AKT1 and HAK5 (Hirsch et al. 1998; Gierth et al. 2005; Pyo et al. 2010). HAK5 accounts for a large portion of the high-affinity K+ absorption, and its activity is suppressed by NH4 + (Santa-Maria et al. 1997). AKT1 is a NH4 +-insensitive K+ uptake component, and mediates K+ absorption covering a large range of exogenous K+ concentrations, even at micromolar concentrations (Rubio et al. 2008). The gene encoding AKT1 was the first cloned plant inward-rectifying K+ channel gene from A. thaliana (Sentenac et al. 1992). AKT1 was located in epidermis, cortex, and endodermis of the root (Cao et al. 1995; Lagarde et al. 1996; Fuchs et al. 2005). In A. thaliana, AKT1 was activated by low K+ medium, but its transcript levels were not influenced by disparate adscititious K+ from the concentrations range of low to high, implying that the expression of AKT1 was possibly post-transcriptional regulation (Maathuis and Sanders 1996; Maathuis et al. 2003; Spalding et al. 1999; Hampton et al. 2004). In spite of the transcription abundance of OsAKT1 was not affected by external K+, OsAKT1 was down-regulated in Oryza sativa responding to salt treatment, in-line with the significant reduction of inward K+ currents in root protoplasts (Fuchs et al. 2005). Both transcripts and protein expression of AKT1-type K+ channel McAKT1 in ice plant (Mesembryanthemum crystallinum) were also enormously declined under saline condition (Su et al. 2001). These results indicated that AKT1 was inhibited by salt stress. However, AKT1 has been shown to possibly mediate Na+ influx. In a comparison of two rice varieties with different salt resistance, Golldack et al. (2003) showed higher OsAKT1 transcript levels in the sensitive cultivar IR29 than that in the tolerant cultivar Pokkali in response to Na+. Another research in rice showed that Na+ in leaf protoplasts of the salt-sensitive cultivar was decreased by approximately 50 % when the K+ channels inhibitors Cs+ and TEA+ were added, whereas the salt-tolerant variety was not influenced by Cs+ and TEA+ (Kader and Lindberg 2005). A recent study suggested that AtAKT1 is possibly a channel for K+ efflux from the root during salt stress (Nieves-Cordones et al. 2010). These results provided entirely different viewpoints about AKT1 under salt stress. Therefore, the role of AKT1 in P. tenuiflora needs to be further revealed.
To address the role of AKT1 in salt tolerance, we first isolated and characterized PtAKT1 gene from P. tenuiflora. Second, we investigated gene expression patterns and ion concentrations of potassium and sodium in P. tenuiflora under different KCl and NaCl treatments. The results suggested the important role of AKT1 in selective absorption for K+ over Na+ in P. tenuiflora.
Materials and methods
Plant culture and treatments
Seeds’ germination and seedlings’ growth conditions of P. tenuiflora were performed according to the methods in Wang et al. (2009). One-month-old seedlings were transferred to the modified Hoagland nutrient solution deprived of KNO3 for 7 days and 5 mM KNO3 was substituted by 2.5 mM NH4NO3. Then the plants were treated as follows: (i) 0, 0.1, 0.5, 1, 5 or 10 mM KCl was added separately for 48 h; (ii) 0.1 or 5 mM KCl plus 25 or 150 mM NaCl for 0, 6, 12, 24, 48, 72 and 96 h, respectively.
Cloning of PtAKT1 and sequence analysis
Total RNA was isolated from roots under 150 mM NaCl for 48 h using the Trizol Kit (Sangon, China) (Wu et al. 2011; Yuan et al. 2015). Reverse transcription was performed using MMLV-RTase (Sangon, China). A segment of PtAKT1 was amplified with primers P1 and P2 (Table S1). PCR fragment was ligated to the pUCm-T vector (Sangon, China) after purification and then sequenced by Sangon. The 5′- and 3′-cDNA were acquired using the kit of RNA ligase mediated rapid amplification of 5′- and 3′-cDNA Ends (RLM-RACE, Invitrogen, USA) according to the manufacturer’s instructions, 5′- and 3′-ends of PtAKT1 were amplified using the special primers P3, P4, P5 and P6, respectively (Table S1). The full-length of the PtAKT1 cDNA was obtained by assembling these fragments. The final sequence was analyzed as described by Guo et al. (2012).
Semi-quantitative RT-PCR
Primers P7 and P8 (Table S1) were used to test the transcript level of PtAKT1. PtACTIN (GenBank accession No. FJ545641) was used as the reference gene for RT-PCR experiments. Specific primers of PtACTIN t were A1 and A2 (Table S1). The PCR procedure was given below: 2 min at 94 °C; 30 cycles of 30 s at 94 °C, 25 s at 52 °C (PtAKT1)/50 s at 56 °C (PtACTIN), 40 s at 72 °C. PCR products were separated on 1.0 % agarose gels.
Determination of Na+ and K+ concentration
Roots of P. tenuiflora were soaked for 8 min in icy calcium chloride to swap the apoplastic ion and shoots were washed by distilled water to remove surface salts (Wang et al. 2007). Each plant was cut into roots and shoots before they dried off. Put dried samples into 100 mM glacial acetic acid and bath over 90 °C for 2 h to extract cation. Na+ and K+ concentrations were determined with a flame spectrophotometer (Model 410, Sherwood Scientific Ltd, UK).
Calculation of SA value and statistical analysis
Selective absorption (SA) capacity for K+ over Na+ was calculated in accordance with the equation given below: SA = (K+/Na+ in whole plant)/(K+/Na+ in medium) (Wang et al. 2002, 2004). Statistical analysis was performed according to Guo et al. (2012).
Results
Molecular cloning and primary structure of PtAKT1
The full-length cDNA of PtAKT1 was reconstituted from P. tenuiflora by PCR, 5′- and 3′-RACE, it was 3100 bp long with a 5′-UTR of 74 bp nucleotides, a predicted ORF of 2688 bp nucleotides, and a 3′-UTR of 338 bp. The deduced polypeptide was 895 amino acids long with an estimated molecular mass of 100.8 kDa and a theoretical isoelectric point of 8. 3.
Hydrophobicity analysis of the deduced polypeptide showed that PtAKT1 had 6 predicted trans-membrane segments (TM 1–TM 6), a putative cyclic nucleotide binding domain (cNBD) from amino acids residues 409–498, ankyrin (ANK) repeat sequences from residues 572–700, and the pore domain (262–285) that included the characteristic GYGD/E-motif in K+ channels (Fig. 1). Multiple sequence alignment showed that there was high identity between PtAKT1 and known-plant AKT1-type channels, especially HvAKT1 or TaAKT1 (88 %). Phylogenetic analysis of plant AKT1-type K+ channels indicated that the family was divided into two branches, and PtAKT1 had closer relationships with monocotyledons (Fig. 2). Therefore, the results showed that PtAKT1 isolated from P. tenuiflora encoded an AKT1-type K+ channel.

Sequence alignment of PtAKT1 with other AKT1 from higher plants. Sources of AKT1 and their GenBank accession numbers are as follows: HvAKT1 (Hordeum vulgare, DQ465992) and TaAKT1 (Triticum aestivum, AF207745). The sequences were aligned with DNAMAN 6.0 software. The six putative trans-membrane domains (TM 1–TM 6) and other domains (pore, cNMP binding domain, and ankyrin repeats domain) are underlined, respectively

Phylogenetic tree of PtAKT1 (framed with red line) with related sequences in plants. The tree was constructed by the neighbor-joining method. The proteins are as follows: HvAKT1 (Hordeum vulgare, DQ465992), TaAKT1 (Triticum aestivum, AF207745), OsAKT1 (Oryza sativa, AY065970), ZmAKT1 (Zea mays, NM_001112010), SbAKT1 (Sorghum bicolor, XM002458189), LlAKT1 (Lilium longiflorum, EF397611), AtAKT1 (Arabidopsis thaliana, AK317729), NtAKT1 (Nicotiana tabacum, AB196790), McAKT1 (Mesembryanthemum crystallinum, AF267753), ZxAKT1 (Zygophyllum xanthoxylum, GQ857474), PeAKT1 (Populus euphratica, EU554630), RcAKT1 (Ricinus communis, XM_002529327), VvAKT1 (Vitis vinifera, FR669116), LeAKT1 (Lycopersicon esculentum, X96390), StAKT1 (Solanum tuberosum, X86021), DcAKT1 (Daucus carota, AJ697979) and EcAKT1 (Eucalyptus camaldulensis, AF197332)
Expression of PtAKT1 and whole plant K+ accumulation in P. tenuiflora treated with KCl
The expression levels of PtAKT1 were great higher in roots than that in shoots (Fig. 3a, b). However, no significant difference was found in the transcript abundance of PtAKT1 among various concentrations of external KCl (0–10 mM) for 48 h both in roots and shoots (Fig. 3b). The results suggested that PtAKT1 was preferentially expressed in the root tissue and its expression level was not affected by external K+ concentrations. Furthermore, additional KCl (0.1–10) had no impact on whole plant K+ accumulation as well (Fig. 3c).

Expression of PtAKT1 and whole plant K+ concentration in P. tenuiflora under different concentrations of KCl (0, 0.1, 0.5, 1, 5 and 10 mM) for 48 h. a Semi-quantitative RT-PCR analysis of PtAKT1 in 4-week-old plants exposed to K+-free nutrient solution for 7 days and then treated with KCl (0, 0.1, 0.5, 1, 5 and 10 mM) for 48 h. b The relative expression level of PtAKT1 (related to ACTIN) in shoots and roots. ACTIN was used as an internal control. Experiments were repeated at least three times to obtain similar results. c Whole plant K+ concentration in P. tenuiflora under different concentrations of KCl for 48 h. Ten plants were pooled in each replicate (n = 8). Values are mean ± SE and bars indicate SE (n = 3–8). Different letters indicate significant difference at P < 0.05 (Duncan’s test)
Expression of PtAKT1 in P. tenuiflora under 25 or 150 mM NaCl plus 0.1 or 5 mM KCl treatments
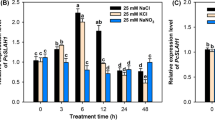
The tissue-specific expression analysis showed that the transcripts of PtAKT1 were detected primarily in roots under salt treatment (data not shown). With prolonging of salt stress, the transcript abundance of PtAKT1 was down-regulated sharply and then tended to stabilization after 6 h under 25 mM NaCl, whereas it was down-regulated gradually and tended to stabilization after 24 h under 150 mM NaCl (Fig. 4a, b). In addition, the expression of PtAKT1 at 150 mM NaCl plus various KCl was significantly higher than that at 25 mM NaCl plus various KCl during 48 h of salt stress, but there were no significant variations among the treatments from 48 to 96 h (Fig. 4b). The results indicated that the transcription abundance of PtAKT1 was restrained rapidly by mild salt treatment (25 mM NaCl), while it was inhibited slowly by severe salt stress (150 mM NaCl).

Time courses of PtAKT1 expression in roots of P. tenuiflora under 25 or 150 mM NaCl plus 0.1 or 5 mM KCl. a Semi-quantitative RT-PCR analysis of PtAKT1 in 4-week-old plants exposed to K+-free nutrient solution for 7 days and then treated with 25 or 150 mM NaCl plus 0.1 or 5 mM KCl over a 96-h period. b The relative expression level of PtAKT1 (related to ACTIN) in roots. ACTIN was used as an internal control. Experiments were repeated at least three times to obtain similar results. Values are mean ± SE (n = 3) and bars indicate SE
The whole plant Na+, K+ accumulation and the relationship between SA values and expression levels of PtAKT1 in P. tenuiflora under 25 or 150 mM NaCl plus 0.1 or 5 mM KCl treatments
The addition of 25 or 150 mM NaCl plus 0.1 or 5 mM KCl all significantly increased whole plant Na+ accumulation with prolonging of the treatments, while the magnitude of the former (25 mM NaCl plus various KCl) was remarkably lower than that of the latter (150 mM NaCl plus various KCl), and the Na+ concentration treated with 150 mM NaCl plus 0.1 mM KCl was higher than that under 150 mM NaCl plus 5 mM KCl (Fig. 5a). Whole plant K+ accumulation tended to decrease with the increase of stress duration, but no significant difference in the whole plant K+ accumulation was found among various treatments during 48–96 h (Fig. 5b).

Time courses of whole plant Na+ (a) and K+ (b) concentration in P. tenuiflora under 25 or 150 mM NaCl plus 0.1 or 5 mM KCl over a 96-h period. Ten plants were pooled in each replicate (n = 8). Values are mean ± SE (n = 8) and bars indicate SE
Moreover, selective absorption (SA) capacity for K+ over Na+ in roots were on a declining curve under all treatments. Either under high or low KCl conditions, SA values under 150 mM NaCl were significantly greater than that under 25 mM NaCl (Fig. 6a, b). Further analysis showed a significantly positive correlation between SA values and expression levels of PtAKT1 in roots under 25 or 150 mM NaCl plus various KCl in all treatment times (Fig. 6c, d).

Selective absorption (SA) capacity for K+ over Na+ in P. tenuiflora exposed to 25 or 150 mM NaCl plus 0.1 (a) or 5 mM KCl (b) and relationship between SA values and relative PtAKT1 expression levels in roots under 25 or 150 mM NaCl plus 0.1 (c) or 5 mM KCl (d) treatments for 6–96 h. Ten plants were pooled in each replicate (n = 8). Values are mean ± SE (n = 3–8) and bars indicate SE
Discussion
The deduced polypeptide sequence of PtAKT1 in the current study showed the typical features that included six trans-membrane segments (TM 1–TM 6) and the pore domain between TM 5 and TM 6, the TM 4 is supposed to be a voltage sensor (Chérel 2004). Similar to AKT1 in A. thaliana (Lee et al. 2007), OsAKT1 in O. sativa (Fuchs et al. 2005), TaAKT1 in wheat (Buschmann et al. 2000), and McAKT1 in M. crystallinum (Su et al. 2001), PtAKT1 possesses a pore domain between TM 5–TM 6 that included the characteristic GYGD/E-motif (Chérel 2004, Fig. 1). The long C-terminal region contained a cyclic nucleotide binding domain (CNBD) and ankyrin repeats, which are thought to be potentially involved in interacting between membrane proteins and cytoskeleton (Sentenac et al. 1992; Véry and Sentenac 2003). Furthermore, the PtAKT1 showed 88 % identity with the HvAKT1 from H. vulgare in amino acid sequence (Fig. 1). Phylogenetic analysis showed that PtAKT1 shared a closer relationship with monocotyledon AKT1-type K+ channels (Fig. 2). These results showed that PtAKT1 encoded an AKT1-type channel in P. tenuiflora.
In line with previous results from A. thaliana (Cao et al. 1995) and rice (Fuchs et al. 2005), we reported here that PtAKT1 expressed primarily in roots of P. tenuiflora (Fig. 3a, b). In A. thaliana, the steady-state level of AKT1 mRNA was not influenced by the concentrations changing of additive K+ (5 μM–5 mM) (Lagarde et al. 1996). The similar results were acquired with Northern blot and micro array (Maathuis and Sanders 1996; Maathuis et al. 2003; Spalding et al. 1999; Hampton et al. 2004). In P. tenuiflora, the transcript level of PtAKT1 was also unaffected by varying K+ concentration in the nutrient solution (Fig. 3b), suggesting that the regulation of external K+ to PtAKT1 could be a post-transcriptional process. Ardie et al. (2010) showed PtAKT1 was induced by K+ starvation, while external KCl was added after the treatment of depriving of K+ in the current study, therefore, our results did not conflict with the finding by Ardie et al. (2010). Furthermore, the whole plant K+ concentration remained unchanged with varying K+ in the medium (Fig. 3c), suggesting K+ uptake in P. tenuiflora covered a large range of external K+ concentrations. It is well known that AKT1 mediates both high- and low-affinity K+ uptake (Hirsch et al. 1998; Rubio et al. 2008). Therefore, PtAKT1 might be the leading component of K+ uptake in P. tenuiflora, especially with existence of NH4 +.
In our current study, the expression abundances of PtAKT1 were down-regulated by NaCl, either under normal (5 mM) or low (0.1 mM) K+ concentrations (Fig. 4a, b). Similar expression patterns in AKT1 were noted in M. crystallinum (Su et al. 2001), rice (Fuchs et al. 2005), A. thaliana (Kaddour et al. 2009) and barley (Boscari et al. 2009). In addition, the whole plant K+ concentrations showed a decrease trend during salt treatment (Fig. 5b). Similar results were found in rice using electrophysiological approach that the OsAKT1 in roots was down-regulated under salt treatment, together with significant reduction in the inward K+ currents (Fuchs et al. 2005). NaCl can depolarize the membrane potential, which results in K+ absorption via the K+ inward-rectifying channel is blocked (Shabala and Cuin 2008; Nieves-Cordones et al. 2010). It appeared that the down-regulation of AKT1 gene expression was likely to be attributed to Na+ entry to cytoplasm. In contrast, Ardie et al. (2010) showed external NaCl (300 mM) did not affect the transcript level of PtAKT1, differences in the results possibly due to different Na+ treatment conditions, on the one hand, 300 mM NaCl is likely to exceed the tolerant range of P. tenuiflora; on the other hand, addition of NaCl is on the basis of K+ starvation in our study, similarly, treatment with NaCl inhibited the expression of PtAKT1 that induced by K+ starvation in the previous research (Ardie et al. 2010).
Regulating plant K+ acquisition and maintaining the homeostasis of K+ and Na+ in protoplasts are especially important for plants to cope with salt stress. Wang et al. (2009) showed that limiting Na+ flow into P. tenuiflora roots and maintaining a powerful selectivity for K+ over Na+ were crucial to salt resistance of the plants. An augment in K+ content and a reduction in Na+ accumulation in transgenic plants of A. thaliana over-expressed with PtAKT1 provided a candidate who controls selective absorption (SA) capacity for K+ over Na+ in molecular aspect (Ardie et al. 2010). In the present study, although salt treatment inhibited the expression of PtAKT1, the transcript levels of PtAKT1 under severe salt treatment (150 mM NaCl) were dramatically higher than that under mild salt condition (25 mM NaCl) within 6–24 h (Fig. 4b). Similarly, either under normal (5 mM) or low (0.1 mM) KCl condition, the SA values in roots during 150 mM NaCl were significantly higher than that during 25 mM NaCl treatments (Fig. 6a, b). A significant positive correlation was found between root PtAKT1 expression levels and SA values under either 25 or 150 mM NaCl plus KCl treatments (Fig. 6c, d), indicating that PtAKT1 is very likely to be the key factor in maintaining selective absorption capacity for K+ over Na+ during saline conditions.
AKT1-type K+ channels exhibit a high selectivity for K+ (Bertl et al. 1997). Over-expression of PtAKT1 results in not only an augment in K+ content but also reduction in Na+ accumulation in A. thaliana (Ardie et al. 2010). An interesting study by Qi and Spalding (2004) proposed that AKT1 K+-acquisition system was protected by SOS1 (plasma membrane Na+/H+ antiporter) system from impairment by Na+, and SOS1 mutation inhibited K+ absorption in roots under salt stress. In fact, as a Na+ efflux transporter, SOS1 was deemed to be related to K+ nutrition in the original researches: Wu et al. (1996) and Ding and Zhu (1997) showed the defect of high-affinity K+ uptake in sos1 mutant. Meanwhile, SOS system may restrain Na+ transporter HKT (high-affinity K+ transporter) (Rus et al. 2001; Uozumi et al. 2000; Fuchs et al. 2005). Recently, we hypothesized a function model of SOS1 and HKT in sustaining the membrane integrity to regulate K+ and Na+ transport system (Guo et al. 2012). In the current work, we suggested that PtAKT1 regulated selective absorption capacity for K+ over Na+. However, the transcript levels of PtAKT1 were down-regulated by NaCl, and the phenomenon that the expression of PtAKT1 under severe salt condition were higher than that under mild salinity seemed to be difficult to explain. Based on all these results, we supplement the previous model (Guo et al. 2012) in this study and suggest a model to detail the role of AKT1 in maintaining selective absorption capacity for K+ over Na+ by roots (Fig. 7). During severe salt stress (Fig. 7a), Na+ in XPCs is excreted to extracellular space by SOS1 and then diffuses into xylem to ameliorate cytoplasmic impairment by Na+ (Shi et al. 2002). Na+ would then flow quickly to the aerial part by transpiration pull, and over-accumulated Na+ in leaves would certainly compromise the photosynthetic tissues (Pardo 2010). Sequestering a portion of Na+ is a strategy to alleviate the damage. Besides, HKT localized in XPCs could retrieve part of Na+ from xylem (Ren et al. 2005; Sunarpi et al. 2005; Byrt et al. 2007; Guo et al. 2012). The second pathway could depolarize membrane of XPCs and then induce K+ loading to the xylem by depolarization-activated SKOR (Stele K+ Outwardly Rectifying) and subsequently promotes K+ transport to shoots (Gaymard et al. 1998; Horie et al. 2009; Guo et al. 2012). Plentiful K+ transport to shoots, through feedback regulation, stimulates the expression of AKT1 located at epidermis, cortex and endodermis, promoting more K+ acquisition from environments, which can, on the one hand, provide adequate K+ nutrition for plants; and on the other hand, increase K+/Na+ ratio in cytoplasm and then mitigate the damage by toxic Na+. Under the mild salt condition (Fig. 7b), Na+ is loaded into xylem via SOS1 and then delivered to shoots since its accumulation do not exceed the capacity of the vacuoles compartmenting Na+ (Blumwald et al. 2000; Shi et al. 2002; Guo et al. 2012). Meanwhile, hydrion is pumped into XPCs by SOS1, resulting in acidification of the cells, which might notably down-regulate the activity of SKOR (Lacombe et al. 2000), thereby negatively regulating the expression of AKT1 and decreasing K+ uptake by roots. This might be the reason why the 25 mM Na+ showed higher repression than 150 mM Na+.

Schematic model for the function of AKT1 in maintaining selective absorption capacity for K+ over Na+ under various salt stress. a Under severe salt stress, the transport activities of HKT outweigh SOS1 at the membrane of XPCs and therefore, Na+ is retrieved from xylem (Guo et al. 2012), Na+ unloading into XPCs possibly depolarises their plasma membrane, which in turn could activate SKOR to load K+ into xylem (Wegner and De Boer 1997; Gaymard et al. 1998; Horie et al. 2009), then stimulates AKT1 to absorb and transport more K+ by feedback regulation. b Under mild salt condition, the activities of SOS1 overwhelm HKT (Guo et al. 2012), SOS1 loads Na+ to xylem directly and transfer it to shoots, at the same time, hydrion is pumped into XPCs by this Na+/H+ antiporter, causing decline in pH of XPCs, which might notably down-regulate the activity of SKOR (Lacombe et al. 2000; Shi et al. 2002), accordingly, that extensive K+ detained in XPCs might negatively regulate the expression of AKT1, reducing K+ uptake by roots. The size of the arrows indicate the capabilities of proteins
In conclusion, the results in this paper suggest that PtAKT1 maintain a strong selective absorption capacity for K+ over Na+ during various salinity. We hypothesize a model to expound the different effects of various salinity on the expressions of PtAKT1, and provide a likely explanation to how SOS1 protect the AKT1 mediating K+ influx during salt stress.
Author contribution statement
Conceived and designed the experiments: Qiang Guo and Suo-Min Wang. Performed the experiments: Qiang Guo, Pei Wang and Xiang-Rui Zhou. Analyzed the data: Qiang Guo and Pei Wang. Performed the analysis with constructive discussions: Pei Wang and Suo-Min Wang. Wrote the paper: Pei Wang, Qian Wang and Suo-Min Wang.
References
Ardie SW, Liu SK, Takano T (2010) Expression of the AKT1-type K+ channel gene from Puccinellia tenuiflora, PutAKT1, enhances salt tolerance in Arabidopsis. Plant Cell Rep 29:865–874
Bertl A, Reid JD, Sentenac H, Slayman CL (1997) Functional comparison of plant inward-rectifier channels expressed in yeast. J Exp Bot 48:405–413
Blumwald E, Aharon GS, Apse MP (2000) Sodium transport in plant cells. Biochim Biophys Acta 1465:140–151
Boscari A, Clément M, Volkov V, Golldack D, Hybiak J, Miller AJ, Amtmann A, Fricke W (2009) Potassium channels in barley: cloning, functional characterization and expression analyses in relation to leaf growth and development. Plant Cell Environ 12:1761–1777
Buschmann PH, Vaidyanathan R, Gassmann W, Schroeder JI (2000) Enhancement of Na+ uptake currents, time-dependent inward-rectifying K+ channel currents, and K+ channel transcripts by K+ starvation in wheat root cells. Plant Physiol 122:1387–1397
Byrt CS, Platten JD, Spielmeyer W, James RA, Lagudah ES, Dennis ES, Tester M, Munns R (2007) HKT1; 5-like cation transporters linked to Na+ exclusion loci in wheat, Nax2 and Kna1. Plant Physiol 143:1918–1928
Cao Y, Ward JM, Kelly WB, Ichida AM, Gaber RF, Anderson JA, Uozumi N, Schroeder JI, Crawford NM (1995) Multiple genes, tissue specificity, and expression dependent modulation contribute to the functional diversity of potassium channels in Arabidopsis thaliana. Plant Physiol 109:1093–1106
Chérel I (2004) Regulation of K+ channel activities in plants: from physiological to molecular aspects. J Exp Bot 55:337–351
Ding L, Zhu JK (1997) Reduced Na+ uptake in the NaCI-hypersensitive sosl mutant of Arabidopsis thaljana. Plant Physiol 113:795–799
Flowers TJ (2004) Improving crop salt tolerance. J Exp Bot 55:307–319
Fuchs I, Stölzle S, Ivashikina N, Hedrich R (2005) Rice K+ uptake channel OsAKT1 is sensitive to salt stress. Planta 221:212–221
Gaymard F, Pilot G, Lacombe B, Bouchez D, Bruneau D, Boucherez J, Michaux-Ferriere N, Thibaud JB, Sentenac H (1998) Identification and disruption of a plant shaker-like outward channel involved in K+ release into the xylem sap. Cell 94:647–655
Gierth M, Mäser P (2007) Potassium transporters in plants involvement in K+ acquisition, redistribution and homeostasis. FEBS Lett 581:2348–2356
Gierth M, Mäser P, Schroeder JI (2005) The potassium transporter AtHAK5 functions in K+ deprivation-induced high-affinity K+ uptake and AKT1 K+ channel contribution to K+ uptake kinetics in Arabidopsis roots. Plant Physiol 137:1105–1114
Golldack D, Quigley F, Michalowski CB, Kamasani UR, Bohnert HJ (2003) Salinity stress-tolerant and -sensitive rice (Oryza sativa L.) regulate AKT1-type potassium channel transcripts differently. Plant Mol Biol 51:71–81
Guo Q, Wang P, Ma Q, Zhang JL, Bao AK, Wang SM (2012) Selective transport capacity for K+ over Na+ is linked to the expression levels of PtSOS1 in halophyte Puccinellia tenuiflora. Funct Plant Biol 39:1047–1057
Hampton CR, Bowen HC, Broadley MR, Hammond JP, Mead A, Payne KA, Pritchard J, White PJ (2004) Cesium toxicity in A rabidopsis. Plant Physiol 136:3824–3837
Hirsch RE, Lewis BD, Spalding EP, Sussman MR (1998) A role for the AKT1 potassium channel in plant nutrition. Science 280:918–921
Horie T, Hauser F, Schroeder JI (2009) HKT transporter-mediated salinity resistance mechanisms in Arabidopsis and monocot crop plants. Trends Plant Sci 14:660–668
Kaddour R, Nasri N, M’rah S, Berthomieu P, Lachaâl M (2009) Comparative effect of potassium in K and Na uptake and transport in two accessions of Arabidopsis thaliana during salinity stress. C R Biol 332:784–794
Kader MA, Lindberg S (2005) Uptake of sodium in protoplasts of salt-sensitive and salt-tolerant cultivars of rice, Oryza sativa L. determined by the fluorescent dye SBFI. J Exp Bot 56:3149–3158
Lacombe B, Pilot G, Gaymard F, Sentenac H, Thibaud J-B (2000) pH control of the plant outwardly-rectifying potassium channel SKOR. FEBS Lett 466:351–354
Lagarde D, Basset M, Lepetit M, Conejero G, Gaymard F, Astruc S, Grignon C (1996) Tissue-specific expression of Arabidopsis AKT1 gene is consistent with a role in K+ nutrition. Plant J 9:195–203
Lee SC, Lan WZ, Kim BG, Li LG, Cheong YH, Pandey GK, Lu GH, Buchanan BB, Luan S (2007) A protein phosphorylation/dephosphorylation network regulates a plant potassium channel. Proc Natl Acad Sci USA 104:15959–15964
Maathuis FJM, Sanders D (1996) Mechanism of potassium absorption by higher plant roots. Physiol Plant 96:68–78
Maathuis FJM, Filatov V, Herzyk P, Krijger GC, Axelsen KB, Chen S, Green BJ, Li Y, Madagan KL, Sánchez-Fernández R, Forde BG, Palmgren MG, Rea PA, Williams LE, Sanders D, Amtmann A (2003) Transcriptome analysis of root transporters reveals participation of multiple gene families in the response to cation stress. Plant J 35:675–692
Nieves-Cordones M, Alemán F, Martínez V, Rubio F (2010) The Arabidopsis thaliana HAK5 K+ transporter is required for plant growth and K+ acquisition from low K+ solutions under saline conditions. Mol Plant 3:326–333
Pardo JM (2010) Biotechnology of water and salinity stress tolerance. Curr Opin Biotech 21:185–196
Peng YH, Zhu YF, Mao YQ, Wang SM, Su WA, Tang ZC (2004) Alkali grass resists salt stress through high [K+] and an endodermis barrier to Na+. J Exp Bot 55:939–949
Pyo YJ, Gierth M, Schroeder JI, Cho MH (2010) High-affinity K+ transport in Arabidopsis: AtHAK5 and AKT1 are vital for seedling establishment and postgermination growth under low-potassium conditions. Plant Physiol 153:863–875
Qi Z, Spalding EP (2004) Protection of plasma membrane K+ transport by the salt overlay sensitive Na+/H+ antiporter during salinity stress. Plant Physiol 136:2548–2555
Ren ZH, Gao JP, Li LG, Cai XL, Huang W, Chao DY, Zhu MZ, Wang ZY, Luan S, Lin HX (2005) A rice quantitative trait locus for salt tolerance encodes a sodium transporter. Nat Genet 37:1141–1146
Rubio F, Nieves-Cordones M, Alemán F, Martínez V (2008) Relative contribution of AtHAK5 and AtAKT1 to K+ uptake in the high-affinity range of concentrations. Physiol Plant 134:598–608
Rus A, Yokoi S, Sharkhuu A, Reddy M, Lee BH, Matsumoto TK, Koiwa H, Zhu JK, Bressan RA, Hasegawa PM (2001) AtHKT1 is a salt tolerance determinant that controls Na+ entry into plant roots. Proc Natl Acad Sci USA 98:14150–14155
Santa-Maria GE, Rubio F, Dubcovsky J, Rodriguez-Navarro A (1997) The HAK1 gene of barley is a member of a large gene family and encodes a high-affinity potassium transporter. Plant Cell 9:2281–2289
Sentenac H, Bonneaud N, Minet M et al (1992) Cloning and expression in yeast of a plant potassium ion transport system. Science 256:663–665
Shabala S, Cuin TA (2008) Potassium transport and plant salt tolerance. Physiol Plant 133:651–669
Shi H, Quintero FJ, Pardo JM, Zhu JK (2002) The putative plasma membrane Na+/H+ antiporter SOS1 controls long-distance Na+ transport in plants. Plant Cell 14:465–477
Spalding EP, Hirsch RE, Lewis DR, Qi Z, Sussman MR, Lewis BD (1999) Potassium uptake supporting plant growth in the absence of AKT1 channel activity: inhibition by ammonium and stimulation by sodium. J Gen Physiol 113:909–918
Su H, Golldack D, Katsuhara M, Zhao C, Bohnert HJ (2001) Expression and stress-dependent induction of potassium channel transcripts in the common ice plant. Plant Physiol 125:604–614
Sunarpi HT, Motoda J, Kubo M, Yang H, Yoda K, Horie R, Chan WY, Leung HY, Hattori K, Konomi M, Osumi M, Yamagami M, Schroeder JI, Uozumi N (2005) Enhanced salt tolerance mediated by AtHKT1 transporter-induced Na+ unloading from xylem vessels to xylem parenchyma cells. Plant J 44:928–938
Uozumi N, Kim EJ, Rubio F, Yamaguchi T, Muto S, Tsuboi A, Bakker EP, Nakamura T, Schroeder JI (2000) The Arabidopsis HKT1 gene homolog mediates inward Na+ currents in Xenopus laevis oocytes and Na+ uptake in Saccharomyces cerevisiae. Plant Physiol 122:1249–1259
Véry AA, Sentenac H (2003) Molecular mechanisms and regulation of K+ transport in higher plants. Annu Rev Plant Biol 54:575–603
Wang SM, Zheng WJ, Ren JZ, Zhang CL (2002) Selectivity of various types of salt-resistant plants for K+ over Na+. J Arid Environ 52:457–472
Wang SM, Zhao GQ, Gao YS, Tang ZC, Zhang CL (2004) Puccinellia tenuiflora exhibits stronger selectivity for K+ over Na+ than wheat. J Plant Nutr 27:1841–1857
Wang SM, Zhang JL, Flowers TJ (2007) Low affinity Na+ uptake in the halophyte Suaeda maritima. Plant Physiol 145:559–571
Wang CM, Zhang JL, Liu XS, Li Z, Wu GQ, Cai JY, Flowers TJ, Wang SM (2009) Puccinellia tenuiflora maintains a low Na+ level under salinity by limiting unidirectional Na+ influx resulting in a high selectivity for K+ over Na+. Plant Cell Environ 32:486–496
Wegner LH, De Boer AH (1997) Properties of two outward rectifying channels in root xylem parenchyma cells suggest a role in K+ homeostasis and long-distance signaling. Plant Physiol 115:1707–1719
Wu SJ, Ding L, Zhu JK (1996) SOS1, a genetic locus essential for salt tolerance and potassium acquisition. Plant Cell 8:617–627
Wu GQ, Xi JJ, Wang Q, Bao AK, Ma Q, Zhang JL, Wang SM (2011) The ZxNHX gene encoding tonoplast Na+/H+ antiporter from the xerophyte Zygophyllum xanthoxylum plays important roles in response to salt and drought. J Plant Physiol 168:758–767
Yuan HJ, Ma Q, Wu GQ, Wang P, Hu J, Wang SM (2015) ZxNHX controls Na+ and K+ homeostasis at the whole-plant level in Zygophyllum xanthoxylum through feedback regulation of the expression of genes involved in their transport. Ann Bot 115:495–507
Acknowledgments
Thanks are due to Professor Yi-wei Jiang from Purdue University, USA, for revising the manuscript. This work was supported by the National Basic Research Program of China (973 Program, Grant No. 2014CB138701), the National Natural Science Foundation of China (Grant No. 31170431), the Fundamental Research Funds for the Central Universities (Grant No. lzujbky-2013-239) and Scholarship Award for Excellent Doctoral Student granted by Lanzhou University.
Author information
Authors and Affiliations
Corresponding author
Additional information
Communicated by P. Sowinski.
P. Wang, Q. Guo contributed equally to this work.
Electronic supplementary material
Below is the link to the electronic supplementary material.
Rights and permissions
About this article

Cite this article
Wang, P., Guo, Q., Wang, Q. et al. PtAKT1 maintains selective absorption capacity for K+ over Na+ in halophyte Puccinellia tenuiflora under salt stress. Acta Physiol Plant 37, 100 (2015). https://doi.org/10.1007/s11738-015-1846-3
Received:
Revised:
Accepted:
Published:
DOI: https://doi.org/10.1007/s11738-015-1846-3