
Abstract
Purpose
Distinct anatomical rearrangements of the gastrointestinal tract achieved by various types of bariatric surgery cause changes in nutrient intake and gut microbiota. The contribution of such gut microbiota changes to remission of type 2 diabetes (T2D) remains unclear.
Aim
We examined gut microbiota changes following banded Roux-en-Y gastric bypass (RYGB) or sleeve gastrectomy (SG) in a randomised study, in relation to T2D remission.
Materials and Methods
Whole-metagenome shotgun sequencing was carried out on paired stool samples at pre- and 1-year post-surgery collected from 44 participants with T2D randomised to banded Roux-en-Y gastric bypass (RYGB) or sleeve gastrectomy (SG). Taxonomic composition and predicted functional potential of the gut bacteria were identified using HUMANn2, and annotated using MetaCyc. Five-day dietary records (analysed using FoodWorks v8.0), body weight and diabetes status were recorded at both time points.
Results
RYGB participants had higher percentage excess weight loss than SG (p = 0.01), even though dietary intake was similar at 1-year post-surgery. Similar proportions achieved diabetes remission (HbA1c < 48 mmol/mol without medications) after either RYGB (68%) or SG (59%). RYGB resulted in increased abundances of Firmicutes and Proteobacteria, while SG resulted in increased Bacteroidetes. Pre-surgery, an increased abundance of Eubacteriaceae (p = 0.01) and Alistipes putredinis (p = 0.01) was observed in those who went on to remit from T2D post-surgery. Following surgery, Lachnospiraceae (p = 0.04) and Roseburia (p = 0.01) species were more abundant in those who had achieved T2D remission.
Conclusions
Specific stool bacterial taxa may signal likelihood of T2D remission after bariatric surgery which is potentially mediated by increases in Lachnospiraceae and Roseburia.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
Bariatric surgery is currently the only therapy that achieves long-term weight reduction, and has dramatic effects on both remission and prevention of type 2 diabetes (T2D) among those with severe obesity [1]. Both Roux-en-Y gastric bypass (RYGB) and sleeve gastrectomy (SG) achieve rapid and sustained weight loss of up to 70%, and remission of T2D in 70–85% of patients [2,3,4], with very few complications [3, 5]. While there are significant differences in the anatomical rearrangements with RYGB and SG [6, 7], the contribution of the associated changes in gut microbiota to the metabolic benefits arising after bariatric surgery is unclear [8].
Obesity and T2D have been reported to hold distinct gut microbial ‘signatures’ [9,10,11,12,13] that are very different to those of a heathy, non-obese population. Several studies to date have also described dramatic alterations to the gut microbiota following bariatric surgery [14,15,16,17,18,19,20,21,22,23,24,25,26,27]. However, due to the heterogeneity of these studies (in terms of surgery type, sample number, methods used and follow-up periods), exactly which of the many reported changes in the gut microbiota are causally linked to the metabolic benefits (e.g. diabetes remission) remains unclear.
Diet [28] and medication use [29] are major contributing factors to the composition of the gut microbiota [30, 31]. Following a change to low calorie diet, large and immediate alterations to the gut bacterial community have been observed [28, 32]. Given that bariatric surgery results in a significant reduction in caloric intake as well as alterations in dietary macronutrient composition, this may represent a confounding factor. Further to this, many glucose-lowering and anti-acid medications (metformin, proton pump inhibitors) have also been shown to alter the gut bacterial community [33,34,35,36,37,38]. Therefore, it is vital to address diet and medication use before and after surgery, in order to accurately tease out surgery-specific changes to the gut microbiota.
In this study, we wished to identify whether there were surgery-specific changes in gut microbiota among obese people with T2D randomised to either SG or banded RYGB and whether there were common taxa and gut microbiota functional capacity changes among those who achieved T2D remission, irrespective of surgery type. We also wished to identify whether there were any baseline gut microbial predictors of T2D remission following surgery.
Methods
Ethics
The trial ‘Sleeve gastrectomy versus Roux-en-Y gastric bypass for type 2 diabetes and morbid obesity: double-blind randomised clinical trial’ [39] is registered with the Australian New Zealand Clinical Trials Registry (ACTRN12611000751976) and received approval from the New Zealand regional ethics committee (NTY/11/07/082). All participants gave written informed consent. This RCT commenced in September 2011 and recruitment completed in October 2014. Five-year data collection is currently ongoing.
Randomised Controlled Trial
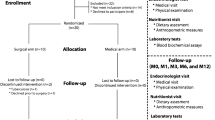
One hundred and fourteen obese participants with T2D were randomised to either RYGB or SG as part of a double-blind RCT with the primary outcome of diabetes remission, defined as HbA1c < 6.5% (< 48 mmol/mol) without the use of diabetes medication at 5 years [39].
Inclusion criteria for patients were: age between 20 and 56 years, a body mass index (BMI) of 35–65 kg/m2 and T2D diagnosis duration at least 6 months. Patients were excluded from participating in the trial if they had a BMI greater than 65 kg/m2, type 1 diabetes, were pregnant, or were previously diagnosed with cancer, severe lung, kidney, liver, or heart disease, due to increased risk of complications during bariatric surgery in these particular situations. Patients were assessed 2 days prior to surgery and at 12 months following surgery. A detailed protocol has been published previously [39].
Very Low Calorie Diet and Surgery
Prior to surgery, participants were prescribed a very low calorie diet (VLCD) for 2 weeks to reduce hepatic steatosis and the risks associated with laparoscopic abdominal surgery. The diet consisted of non-starch vegetables and three daily servings of Optifast (Nestle, Vevey, Switzerland), each of which contained approximately 200 cal.
SG was performed starting 2 cm proximal to the pylorus using serial applications of a stapler device over a 36Fr oro-gastric bougie. RYGB was created with a gastric pouch formed around a 32Fr orogastric tube and a 100-cm Roux limb anastomosed to this pouch (gastrojejunostomy). A 50-cm biliopancreatic limb was then anastomosed in a side-to-side stapled fashion to the distal Roux limb. A 6.5-cm silastic ring was then secured around the gastric pouch approximately 2 cm above the gastrojejunostomy, representing a banded RYGB.
Post-operative Assessments for T2D Status
All pharmacological agents for diabetes were stopped at the time of surgery. Post-surgery, pharmacological treatment for diabetes was restarted if mean post-operative capillary glucose exceeded 12 mmol/L [39]. Postoperatively, all participants were prescribed a twice daily multivitamin: either Band Buddies (NutriChew, Brisbane, Australia; each tablet containing 300-mg elemental calcium and 500-IU vitamin D3) or Centrum 50+ (Pfizer New Zealand, Auckland, New Zealand; each tablet containing 200-mg elemental calcium and 600-IU vitamin D3).
Diabetes remission at 1 year was defined as HbA1c < 6.5% (48 mmol/mol) without glucose lowering therapies. Percent excess weight loss (%EWL) was calculated as the difference between initial BMI and final BMI, divided by the difference between the initial BMI and a target BMI of 25 kg/m2. Dietary intake data was collected from food diaries in which participants were instructed to record everything they ate and drank (including supplements) for 5 days immediately prior to the pre-surgery and 1-year study assessment visits. Dietary nutrient analysis was conducted using FoodWorks version 8.0 (Xyris software, Highgate Hill, Melbourne, Australia). Items were coded into protein, total fat, saturated fat, carbohydrate, fibre and estimated total caloric intake in kilocalories.
Faecal Samples and DNA Isolation
Two days prior to each assessment (pre-surgery and 1 year following surgery) faecal samples were self-collected as follows: a plastic collection tray was placed in the toilet bowl; participants produced their sample into the collection container (without urinating); using gloves, a small piece of stool was scooped into the faecal collection tube using the scoop on the lid of the faecal collection tube, sealed and placed into a larger container filled with water, sealed again and frozen immediately in the home freezer (− 20 °C). On the day of the appointment, samples were transported (frozen) to the laboratory where they were stored (− 80 °C) until DNA extraction. DNA was extracted using the Qiagen QIamp DNA stool mini kit (according to the manufacturer’s instructions).
DNA Sequencing and Library Construction
Whole genome shotgun sequencing was performed on extracted genomic DNA using an Illumina HiSeq 2000 platform. Paired-end reads were trimmed using trimmomatic (v0.33, min length 50 bp), and human contaminants were removed using python scripts (remove_contaminants.py). Microbial taxa and microbiota functional profiles were identified using HUMANn2, which incorporates both MetaPhlan 2.0 for species and taxonomic identification, and HUMANn for metabolic pathway identification using the MetaCyc database (https://MetaCyc.org) [40]. All taxa and pathway data were reported as relative abundances in each sample. Processed sequence data (to remove human sequences) is available on NCBI Sequence Read Archive (https://www.ncbi.nlm.nih.gov/sra) project number PRJNA588805.
Statistical Analyses
Alpha diversity was calculated using both Shannon and Simpson indices. One-way ANOVA and Student’s t test were used to identify significant differences between alpha diversity in each group (surgery group or remitters vs non-remitters).
Permutational multivariate analysis of variance (PERMANOVA) was undertaken to identify differences in the variables of time point, sex, surgery type, BMI (at baseline) and HbA1c (both as a continuous variable and with a categorical variable of ‘remitted’ or ‘not remitted’ defined as outlined above).
Linear discriminant analysis (LDA) effect size (LEfSe) is a computational method used specifically for biomarker discovery within high-dimensional class (group) comparisons. LEfSe identifies features (bacterial species and genetic pathways) that most likely explain the differences observed between groups (surgery type, before or after surgery and T2D status) by coupling standard tests for statistical significance with additional tests encoding biological consistency. Finally, the effect size generated by LEfSe provides an estimation of the magnitude of the observed change due to each of the groups specified [41].
Here we used LEfSe to identify statistically significant differences in relative abundances of gut microbiota and metabolic pathways between pre- and post-operative (1 year) RYGB or SG patients, as well as between those who remitted from T2D and those who did not, 1-year post-surgery. A default cutoff value of LDA > 2.0 was used in all tests. Mann-Whitney tests were used to confirm significant differences between taxa or pathways identified by LEFSe. A p value < 0.05 was considered statistically significant.
Results
Participant Characteristics
Of the 114 participants in the randomised controlled trial who received either RYGB or SG surgery, 44 participants (including 14 whose samples were analysed previously [21]) provided faecal samples at both baseline and 1 year (Supplementary Fig. 1). Age, BMI, HbA1c, T2D medications and T2D status did not differ significantly by treatment group at baseline (Table 1). There was significantly greater %EWL 1-year post-operatively after RYGB compared with SG (p = 0.01). T2D remission was achieved at 1 year in 68% of those in the RYGB group and 59% in the SG group. The number of participants on insulin therapy and/or metformin treatment decreased at 1 year following both types of surgery (Table 1). Nutrient intake in kcal, protein, carbohydrate, total fat, saturated fat and fibre is outlined in Table 1. The total caloric intake increased 1 year after surgery following both RYGB and SG, when compared with the VLCD consumed 4 weeks prior to surgery. There were no significant differences between calorie intake and diet macronutrient composition between the surgery groups at baseline or 1 year following surgery.
Surgery-Specific Changes to the Gut Microbiota Composition and Functional Capacity
We observed no difference in alpha diversity from baseline to 1 year post-operatively for all 44 samples, or at 1 year following either RYGB (n = 22), or SG (n = 22).
We identified both time point (pre- vs post-surgery) and sex as significantly associated with microbiome taxonomic composition using PERMANOVA (p = 0.023 for time point and p = 0.041 for sex). We therefore adjusted for time point and sex, where possible, in subsequent analyses.
Using LEfSe [41], we observed small phylum level differences from baseline to 1 year following each type of surgery (Fig. 1). RYGB surgery was associated with an increased abundance of the phyla Firmicutes and Proteobacteria at 1-year post-surgery (Fig. 1a), whereas, SG surgery was associated with an increased abundance of Bacteroidetes post-surgery (Fig. 1b).

Surgery specific differences in the gut bacteria from pre-surgery (0, red) to 1 year post surgery (1, green) following RYGB (a) and SG (b). Red areas/bars indicate those taxa that were most abundant pre-surgery. Green areas/bars indicate those taxa that were more abundant at 1 year
To determine if the predicted functional capacity of the gut microbial composition changed as a result of either RYGB and or SG, we used LEfSe to identify changes from baseline to 1 year within each surgery type. LEfSe identified no predicted metabolic functions to be differentially abundant for either surgery type from baseline to 1 year post-surgery.
In a cross-sectional analysis at 1 year post-surgery, LEfSe analysis identified only a single order—Enterobacteriales (phylum = Proteobacteria) that was significantly more abundant (LDA = 4.21, p = 0.0084) in patients within the RYGB surgery group (Fig. 2) when compared with the SG group. We did not identify any differentially abundant functional capacity features of the gut microbiome for the same cross sectional analysis of RYGB vs SG at 1-year post-surgery.

Differentially expressed taxa at 1 year after RYGB versus SG. a Red bar indicates the single differentially abundant taxa between RYGB and SG 1 year after surgery. b Individual box plot of Enterobacteriales
Baseline Gut Microbial Composition and Functional Capacity Differences Among those with Subsequent Diabetes Remission Vs Non-diabetes Remission
LEfSe identified four bacterial taxa that were more abundant in the pre-surgery faeces of those who remitted from T2D at 1 year (Fig. 3a). Both the family Eubacteriaceae (phylum Firmicutes) and the species Alistipes putredinis (phylum Bacteroidetes) were significantly more abundant (p = 0.01 (Eubacteriaceae), p = 0.009 (Alistipes putredinis)) in the pre-surgery gut only in those who went on to remit from T2D following surgery (Fig. 3b). This was true irrespective of surgery type. Importantly, the abundance of these Eubacteriaceae and Alistipes putredinis taxa did not correlate with duration of T2D pre-surgery (Supplementary Fig. 2).

Differentially abundant taxa in the baseline gut separated by T2D status a identified by LEfSe, and b Individual box plots of Eubacteriaceae and Alistipes putredinis by surgery type and T2D status. Red bars indicate taxa that were more abundant pre-surgery in those with persisting T2D at 1-year, and green bars indicate taxa that were more abundant pre-surgery in those who remitted from T2D at 1 year. R = remitted (green bars), NR = non-remitted (red bars). Squares indicate whole cohort, triangles indicate SG and circles indicate RYGB
LEfSe was used to identify predicted metabolic pathways that were significantly different between T2D remitters and non-remitters. At baseline, those who went on to remit at 1 year had significantly increased guanosine nucleotide degradation and n10-formyl-tetrahydrofolate biosynthesis, a component of purine base synthesis (Fig. 5).
Gut Microbial Composition and Functional Capacity Among T2D Remitters Vs Non-remitters 1 Year Following Surgery
LEfSe analysis identified the family Lachnospiraceae (phylum Firmicutes) (p = 0.036) and the genus Roseburia (phylum Firmicutes) (p = 0.0047) as being more abundant 1-year post-surgery in diabetes remitters, regardless of surgery type (Figs. 4 and 5). At 1 year post-surgery, increased purine biosynthesis was observed only in those with persistent T2D (Fig. 6). However, this was not significant after post hoc analyses (Mann-Whitney, p = 0.93).

Differentially abundant taxa in the gut microbiota 1-year post surgery, separated by T2D status a identified by LEfSe, and b individual box plots of identified taxa by surgery type and T2D status. Red bars indicate taxa that were more abundant in those with persisting T2D at 1-year, and green bars indicate taxa that were more abundant in those who remitted from T2D at 1 year. R = remitted, NR = non-remitted. Squares indicate whole cohort, triangles indicate SG and circles indicate RYGB

Differentially abundant metabolic pathways in the baseline gut microbiota separated by T2D status a identified by LEfSe, and b individual box plots of identified taxa by surgery type and T2D status. Red bars indicate pathways that were more abundant pre-surgery in those with persisting T2D at 1-year, and green bars indicate pathways that were more abundant pre-surgery in those who remitted from T2D at 1 year. R = remitted, NR = non-remitted. Squares indicate whole cohort, triangles indicate SG and circles indicate RYGB

Differentially abundant pathway in 1 year post surgery, separated by T2D status a identified by LEfSe, and b Individual box plots of identified taxa by surgery type and T2D status. Red bars indicate pathways that were more abundant in those with persisting T2D at 1-year. R = remitted, NR = non-remitted. Squares indicate whole cohort, triangles indicate SG and circles indicate RYGB
Discussion
We have demonstrated that the gut microbiota of those receiving RYGB became enriched in Firmicutes, while the gut microbiota of those receiving SG surgery became enriched in Bacteroidetes, as compared with the baseline gut microbiota. However, these taxonomic changes did not result in any significant difference in overall predicted functional capacity changes of the gut microbiota by type of surgery. Importantly, irrespective of the type of surgery, we identified that Lachnospiraceae and Roseburia taxa increased among those who achieved diabetes remission. We also identified higher levels of Alistipes putredinis species and Eubacteriaceae (family) in baseline stool samples of those who subsequently achieved diabetes remission after either type of bariatric surgery.
Given the comparable T2D remission observed between these two types of bariatric surgery, it is unlikely that surgery-specific changes in gut microbiota will be causal to the common metabolic benefits observed. Indeed, decreases in the relative abundance of Firmicutes after SG and increases in Bacteroidetes and Proteobacteria after RYGB have been reported in two systematic reviews of the changes to the gut microbiota following bariatric surgery [8, 42]. At 1 year after surgery, patients who had banded RYGB surgery had higher Enterobacteriales (Proteobacteria) levels than SG patients, consistent with earlier reports of changes following non-banded RYGB at time points ranging from 3 months to 9.4 years [14, 16,17,18, 20, 26].
We did not see the expected increase in alpha diversity of the gut microbiota from baseline to 1-year post-surgery, in either surgery type, nor was increased alpha diversity associated with T2D remission status. However, the baseline gut microbiota was analysed while patients had been using VLCD in preparation for surgery in which all had markedly reduced their daily caloric intake, which is a lower energy but more nutritionally balanced than a typical obesogenic diet. It has been documented that low caloric intake significantly increases the diversity and richness of the gut bacteria [19, 32]. Therefore, this reduced caloric intake at baseline could have ameliorated possible increases in diversity which may have been seen if we had compared gut microbiota changes after surgery to baseline gut microbiota while on usual diet (prior to VLCD).
The baseline gut microbiota from people who remitted from T2D 1-year post-surgery had greater abundances of Alistipes putredinis species and Eubacteriaceae (family), detected while on VLCD. Alistipes species of the phylum Bacteroidetes are commonly ingested via plant sources, and are known butyrate producers [43]. Since participants were on a VLCD diet for 4 weeks before surgery, consisting only of Optifast shakes, accompanied by non-starch vegetables, it is possible that the higher abundance of Alistipes sp. among subsequent T2D remitters reflects pre-operative adherence to dietary advice, which in turn may predict post-operative dietary adherence promoting T2D remission. Alternatively, this could be a marker of gut dysbiosis that responds best to bariatric surgery, as Alistipes putredinis has also been associated with general gut dysbiosis [44, 45], increased inflammation and irritable bowel syndrome [46]. Some other bariatric studies to date investigating microbiome changes have reported an increased abundance in these species at 6 months [17] and 12 months [20] following RYGB, although they have not looked specifically for gut microbial biomarkers of diabetes remission pre-operatively.
Eubacteriaceae is a core component of a normal healthy gut [47,48,49], hence, may be enriched at baseline among those more likely to remit from T2D. Eubacteriaceae are more commonly shown to be abundant in healthy populations [47], as compared with those of IBS [50]. Interestingly, when the disease subsides in those patients, levels of Eubacteriaceae are shown to increase [50]. Eubacteriaceae has also been shown to be low in abundance in children with gut dysbiosis typically associated with autism spectrum disorder [51, 52], as well as in adults with type 1 diabetes [53]. Although we cannot explain the reason for the observed increase of Eubacteriaceae in the pre-surgery gut among those who went on to remit from T2D after surgery, the observation is consistent with a ‘healthier starting point’ pre-surgery.
Baseline gut microbiota of those who subsequently remitted from T2D had increased capacity for guanosine nucleotide degradation and n10-formyl-tetrahydrofolate biosynthesis. Exogenous purine degradation occurs in the small intestine [54], and includes the conversion of purines (guanine and adenine) to uric acid. This degradation has been known to be primarily carried out by Firmicutes including Bacillus and Enterobacteriaceae [55]. Notably, our data not only showed an increase in the ability to degrade guanine in the pre-surgery gut in those who go on to remit from T2D at 1 year, and also showed an increased abundance of Enterobacteriaceae, indicating this predicted increase in purine degradation in these participants.
The composition of the 1-year post-surgery gut microbiota differed by T2D status, with an increased abundance of the family Lachnospiraceae and the genus Roseburia, both of the Firmicutes phyla, in patients who remitted from T2D. Increased Roseburia levels were also observed only in those achieving T2D remission at 1 year in the earlier pilot analysis (n = 14) [21]. A gut bacterial community abundant in Firmicutes, specifically Lachnospiraceae and Roseburia species is generally deemed to be ‘a healthy gut signature’ [56]. In fact Lachnospiraceae itself is commonly used as a probiotic to promote gut and intestinal health [57]. Inflammatory conditions including Crohn’s disease, ulcerative colitis, IBS and T2D have all been found to show low abundance of both Roseburia and Lachnospiraceae [56, 58], as compared with higher abundances in non-disease or ‘healthy’ people. These specific taxa (Lachnospiraceae and Roseburia) changes have not been observed following dietary weight loss alone [19] but increased Firmicutes following VLCD for 12 weeks show an inverse correlation to resulting body weight. This suggests that the changes we observed among those remitting from T2D after bariatric surgery are similar to those seen after weight loss achieved by dietary means. While we cannot ascertain causality for these taxa in the remission of T2D remission following surgery, evaluating their metabolic benefits as part of probiotic supplementation would be worth testing.
It is important to note that cessation of metformin or other diabetes medications were not related to the changes to the taxonomic composition and functional potential of the gut bacteria seen. Previous studies [33, 34] have shown that metformin use (for as little as 4 weeks) for the treatment of T2D is associated with an increase in genes and pathways related to bacterial environmental response, drug resistance and carbohydrate metabolism, especially enhanced ability to produce short chain fatty acids (SCFAs). Explicitly determining how these gut microbial changes occurring after bariatric surgery contribute to T2D remission requires further work including the application of other mixed-omics approaches [59, 60] that are tailored to analyse the functionality of the gut microbiome, as well as faecal microbial transplant studies and cell-based and animal studies.
Limitations
Firstly, we acknowledge that our sample size was limited. However, it is similar to other bariatric cohorts that have been analysed using shotgun sequencing [16, 18, 19]. Secondly, baseline gut microbiota analysis was conducted while all participants were on the same VLCD at the pre-surgery assessment, so we cannot be sure that the baseline microbial predictors of T2D remission will still hold true if conducted while on usual diet. Nonetheless, such a gut microbial predictor taken while on VLCD, if validated, could still be useful for prognostic purposes before a final decision for surgery prioritisation is made. Finally, as with other studies evaluating changes to gut microbiota following bariatric surgery, we cannot be certain as to which of these are directly due to surgical rearrangement of the gut and which are due to accompanying changes in diet, medication and comorbidities.
Conclusions
In this work, we have identified family, genus and species level differences in the gut microbiota, specifically related to T2D remission status after two types of commonly used bariatric surgery. With further validation of the predictive capacity of certain faecal taxa linked with subsequent T2D remission, in larger cohorts and by using germ-free mice faecal transfer experiments to examine the diabetes remission potential of other taxa, these could be valuable in developing probiotics targeting diabetes remission.
Abbreviations
- ANOVA:
-
Analysis of variance
- BMI:
-
Body mass index
- GI:
-
Gastrointestinal
- LDA:
-
Linear discriminant analysis
- LEfSe:
-
Linear discriminant analysis effect size
- PERMANOVA:
-
Permutational multivariate analysis of variance
- RCT:
-
Randomised controlled trial
- RYGB:
-
Roux-en-Y gastric bypass
- SCFA:
-
Short chain fatty acids
- SG:
-
Sleeve gastrectomy
- T2D:
-
Type 2 diabetes
- VLCD:
-
Very low calorie diet
References
Buchwald H, Estok R, Fahrbach K, et al. Weight and type 2 diabetes after bariatric surgery: systematic review and meta-analysis. Am J Med. 2009;122(3):248–256.e5.
Vidal J, Ibarzabal A, Romero F, et al. Type 2 diabetes mellitus and the metabolic syndrome following sleeve gastrectomy in severely obese subjects. Obes Surg. 2008;18:1077–82.
Lee W-J, Chong K, Ser K-H, et al. Gastric bypass vs sleeve gastrectomy for type 2 diabetes mellitus: a randomized controlled trial. Arch Surg. 2011;146(2):143–8.
Murphy R, Clarke MG, Evennett NJ, et al. Laparoscopic sleeve gastrectomy versus banded Roux-en-Y gastric bypass for diabetes and obesity: a prospective randomised double-blind trial. Obes Surg. 2018;28(2):293–302.
Schauer PR, Kashyap SR, Wolski K, et al. Bariatric surgery versus intensive medical therapy in obese patients with diabetes. N Engl J Med. 2012;366(17):1567–76.
Wolfe BM, Kvach E, Eckel RH. Treatment of obesity: weight loss and bariatric surgery. Circ Res. 2017;118(11):1844–55.
Pories WJ. Bariatric surgery: risks and rewards. J Clin Endocrinol Metab. 2008;93:s89–96.
Davies N, O’Sullivan JM, Plank LD, et al. Altered gut microbiome after bariatric surgery and its association with metabolic benefits: a systematic review. Surg Obes Relat Dis. 2019;15:656–65.
Turnbaugh PJ, Gordon JI. The core gut microbiome, energy balance and obesity. J Physiol. 2009;587(17):4153–8.
Turnbaugh PJ, Hamady M, Yatsunenko T, et al. A core gut microbiom in obese and lean twins. Nature. 2009;457(32089):480–4.
Duncan SH, Lobley GE, Holtrop G, et al. Human colonic microbiota associated with diet, obesity and weight loss. Int J Obes (Lond). 2008;32(11):1720–4.
Wang Y, Luo X, Mao X, et al. Gut microbiome analysis of type 2 diabetic patients from the Chinese minority ethnic groups the Uygurs and Kazaks. PLoS One. 2017;12(3):1–15.
Qin J, Li Y, Cai Z, et al. A metagenome-wide association study of gut microbiota in type 2 diabetes. Nature. 2012;490(7418):55–60.
Zhang H, DiBaise JK, Zuccolo A, et al. Human gut microbiota in obesity and after gastric bypass. Proc Natl Acad Sci U S A. 2009;106(7):2365–70.
Furet J-P, Kong L-C, Tap J, et al. Differential adaptation of human gut microbiota to bariatric surgery-induced weight loss: links with metabolic and low-grade inflammation markers. Diabetes. 2010;59(12):3049–57.
Graessler J, Qin Y, Zhong H, et al. Metagenomic sequencing of the human gut microbiome before and after bariatric surgery in obese patients with type 2 diabetes: correlation with inflammatory and metabolic parameters. Pharmacogenomics J. 2013;13(6):514–22.
Kong LC, Tap J, Aron-Wisnewsky J, et al. Gut microbiota after gastric bypass in human obesity: increased richness and associations of bacterial genera with adipose tissue genes. Am J Clin Nutr. 2013;98(1):16–24.
Tremaroli V, Karlsson F, Werling M, et al. Roux-en-Y gastric bypass and vertical banded gastroplasty induce long-term changes on the human gut microbiome contributing to fat mass regulation. Cell Metab. 2015;22(2):228–38.
Damms-Machado A, Mitra S, Schollenberger AE, et al. Effects of surgical and dietary weight loss therapy for obesity on gut microbiota composition and nutrient absorption. Biomed Res Int. 2015;2015:1–12.
Palleja A, Kashani A, Allin KH, et al. Roux-en-Y gastric bypass surgery of morbidly obese patients induces swift and persistent changes of the individual gut microbiota. Genome Med. 2016;8(1):67.
Murphy R, Tsai P, Jullig M, et al. Differential changes in gut microbiota after gastric bypass and sleeve gastrectomy bariatric surgery vary according to diabetes remission. Obes Surg. 2017;27(4):917–25.
Sanmiguel CP, Jacobs J, Gupta A, et al. Surgically induced changes in gut microbiome and hedonic eating as related to weight loss. Psychosom Med. 2017;79(8):880–7.
Federico A, Dallio M, Tolone S, et al. Gastrointestinal hormones, intestinal microbiota and metabolic homeostasis in obese patients: effect of bariatric surgery. In Vivo. 2016;30(3):321–30.
Patrone V, Vajana E, Minuti A, et al. Postoperative changes in fecal bacterial communities and fermentation products in obese patients undergoing bilio-intestinal bypass. Front Microbiol. 2016;7(FEB):200.
Medina DA, Pedreros JP, Turiel D, et al. Distinct patterns in the gut microbiota after surgical or medical therapy in obese patients. PeerJ. 2017;5(6):3443.
Ilhan ZE, DiBaise JK, Isern NG, et al. Distinctive microbiomes and metabolites linked with weight loss after gastric bypass, but not gastric banding. ISME J. 2017;11(9):2047–58.
Chen H, Qian L, Lv Q, et al. Change in gut microbiota is correlated with alterations in the surface molecule expression of monocytes after Roux-en-Y gastric bypass surgery in obese type 2 diabetic patients. Am J Transl Res. 2017;9(3):1243–54.
Walker AW, Ince J, Duncan SH, et al. Dominant and diet-responsive groups of bacteria within the human colonic microbiota. ISME J. 2011;5(2):220–30.
Imhann F, Vich Vila A, Bonder MJ, et al. The influence of proton pump inhibitors and other commonly used medication on the gut microbiota. Gut Microbes. 2017;8(4):351–8.
Scott KP, Gratz SW, Sheridan PO, et al. The influence of diet on the gut microbiota. Pharmacol Res. 2013;69(1):52–60.
De Filippo C, Cavalieri D, Di Paola M, et al. Impact of diet in shaping gut microbiota revealed by a comparative study in children from Europe and rural Africa. Proc Natl Acad Sci U S A. 2010;107(33):14691–6.
Heinsen FA, Fangmann D, Müller N, et al. Beneficial effects of a dietary weight loss intervention on human gut microbiome diversity and metabolism are not sustained during weight maintenance. Obes Facts. 2017;9(6):379–91.
Wu H, Esteve E, Tremaroli V, Khan MT, Caesar R, Mannerås-holm L, et al. Metformin alters the gut microbiome of individuals with treatment-naive type 2 diabetes , contributing to the therapeutic effects of the drug. Nat Med. 2017;23(7):850–8.
Forslund K, Hildebrand F, Nielsen T, et al. Disentangling type 2 diabetes and metformin treatment signatures in the human gut microbiota. Nature. 2015;528(7581):262–6.
Mardinoglu A, Boren J, Smith U. Confounding effects of metformin on the human gut microbiome in type 2 diabetes. Cell Metab. 2016;23(1):10–2.
Seto CT, Jeraldo P, Orenstein R, et al. Prolonged use of a proton pump inhibitor reduces microbial diversity: implications for Clostridium difficile susceptibility. Microbiome. 2014;2(1):42.
Imhann F, Bonder MJ, Vich Vila A, et al. Proton pump inhibitors affect the gut microbiome. Gut. 2015; https://doi.org/10.1136/gutjnl-2015-310376.
Jackson MA, Goodrich JK, Maxan ME, et al. Proton pump inhibitors alter the composition of the gut microbiota. Gut. 2016;65(5):749–56.
Murphy R, Evennett NJ, Clarke MG, et al. Sleeve gastrectomy versus Roux-en-Y gastric bypass for type 2 diabetes and morbid obesity: double-blind randomised clinical trial protocol. BMJ Open. 2016;6(7):e011416.
Caspi R, Billington R, Fulcher CA, et al. The MetaCyc database of metabolic pathways and enzymes. Nucleic Acids Res. 2018;46(D1):D633–9.
Segata N, Waldron L, Ballarini A, et al. Metagenomic microbial community profiling using unique clade-specific marker genes. Nat Methods. 2012;9(8):811–4.
Luijten JCHBM, Vugts G, Nieuwenhuijzen GAP, et al. The importance of the microbiome in bariatric surgery : a systematic review. Obes Surg. 2019;29:2338–49.
Vital M, Karch A, Pieper DH. Colonic butyrate-producing communities in humans: an overview using omics data. msystems. 2017;2(6):1–18.
Jandhyala SM, Talukdar R, Subramanyam C, et al. Role of the normal gut microbiota. World J Gastroenterol. 2015;21(29):8787–803.
Remely M, Hippe B, Zanner J, et al. Gut microbiota of obese, type 2 diabetic individuals is enriched in Faecalibacterium prausnitzii, Akkermansia muciniphila and Peptostreptococcus anaerobius after weight loss. Endocr Metab Immune Disord Drug Targets. 2016;16:99–106.
Saulnier DM, Riehle K, Mistretta T, et al. Gastrointestinal microbiome signatures of pediatric patients with irritable bowel syndrome. Gastroenterology. 2011;141(5):1782–91.
Jackson DN, Theiss AL. Gut bacteria signaling to mitochondria in intestinal inflammation and cancer. Gut Microbes. 2019;0(0):1–20.
Arrieta M, Finlay BB, Rawls J, et al. The commensal microbiota drives immune homeostasis. Front Immunol. 2012;3(March):1–6.
Lewis DA, Brown R, Williams J, et al. The human urinary microbiome ; bacterial DNA in voided urine of asymptomatic adults. Front Cell Infect Microbiol. 2013;3(August):1–14.
Papa E, Docktor M, Smillie C, et al. Non-invasive mapping of the gastrointestinal microbiota identifies children with inflammatory bowel disease. PLoS One. 2012;7(6):e39242.
De Angelis M, Piccolo M, Vannini L, et al. Fecal microbiota and metabolome of children with autism and pervasive developmental disorder not otherwise specified. PLoS One. 2013;8(10):1–18.
Zhang Y, Li S, Gan R, et al. Impacts of gut bacteria on human health and diseases. Int J Mol Sci. 2015;16:7493–519.
Vaarala O. Gut microbiota and type 1 diabetes. Rev Diabet Stud. 2013;9(4):251–9.
Maiuolo J, Oppedisano F, Gratteri S, et al. Regulation of uric acid metabolism and excretion. Int J Cardiol. 2016;213:8–14.
Vogels GD, Van Der Drift C. Degradation of purines and pyrimidines by microorganisms. Bacteriol Rev. 1976;40(2):403–68.
Frank DN, St Amand AL, Feldman RA, et al. Molecular-phylogenetic characterization of microbial community imbalances in human inflammatory bowel diseases. Proc Natl Acad Sci U S A. 2007;104(34):13780–5.
Dahiya DK, Renuka, Dangi AK, Shandilya UM, Puniya AK, Shukla P. Microbiome and metabolome in diagnosis, therapy and other strategic applications. 2019. 417–424 p.
Million M, Tomas J, Wagner C, et al. New insights in gut microbiota and mucosal immunity of the small intestine. Hum Microbiome J. 2018;7–8(January):23–32.
Mchardy IH, Goudarzi M, Tong M, et al. Integrative analysis of the microbiome and metabolome of the human intestinal mucosal surface reveals exquisite inter-relationships. Microbiome. 2013;1(17):1–19.
Sberro H, Fremin BJ, Zlitni S, et al. Large-scale analyses of human microbiomes reveal thousands of small, novel genes. Cell. 2019;178(5):1245–1259.e14.
Acknowledgements
We would like to acknowledge the following people for their generous contribution to this trial and manuscript: Michael Booth, Bronwen Jones, Hisham Hammodat and Michael G. Clarke.
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Conflict of Interest
The authors declare that they have no conflict of interest.
Ethical Approval Statement
All procedures performed in studies involving human participants were in accordance with the ethical standards of the institutional and/or national research committee and with the 1964 Helsinki declaration and its later amendments or comparable ethical standards.
Informed Consent Statement
Informed consent was obtained from all individual participants included in the study.
Additional information
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Electronic supplementary material
ESM 1
(DOCX 71 kb).
Rights and permissions
About this article

Cite this article
Davies, N., O’Sullivan, J.M., Plank, L.D. et al. Gut Microbial Predictors of Type 2 Diabetes Remission Following Bariatric Surgery. OBES SURG 30, 3536–3548 (2020). https://doi.org/10.1007/s11695-020-04684-0
Published:
Issue Date:
DOI: https://doi.org/10.1007/s11695-020-04684-0