
Abstract
Plantations have been widely established to improve ecosystem services and functioning. Black locust, Robinia pseudoacacia L. is a common, widely planted species to control soil erosion on the Loess Plateau. Previous studies have focused on economic values but the interactions between soil and plant carbon (C), nitrogen (N) and phosphorus (P) remain unknown. Investigating variations of soil, green and senesced leaf C, N and P levels in R. pseudoacacia along a latitudinal gradient is useful to understanding its ecological functions. The results show that soil C, N and senesced leaf N and P significantly decreased with an increase in latitude, but there were no significant changes in the senesced leaf C and soil P. The resorption efficiency of N was related with latitude and soil N levels, and the relation between green leaf N and soil N was significant. These relations suggest that soil N was the key in affecting green leaf N levels. At higher latitudes, senesced leaves had lower N levels associated with higher N resorption efficiency to maintain a stable N content in green leaves. With a decrease of soil N, R. pseudoacacia can enhance N resorption efficiency to meet the demand of growth. Thus, it is an important species for reforestation, especially in nutrient-poor environments.
Similar content being viewed by others

Explore related subjects
Discover the latest articles, news and stories from top researchers in related subjects.Avoid common mistakes on your manuscript.
Introduction
Vegetation restoration is widely implemented globally to improve land cover, reduce soil erosion, and enhance ecosystem services and functioning. Reforestation is regarded as an effective method for restoring vegetative cover. Previous studies showed that reforestation improved soil quality, site biodiversity, carbon fixation, and climate regulation (Sun et al. 2015; Zethof et al. 2019; Zheng et al. 2020). For example, after nearly 30 years of vegetation restoration, forest cover and vegetation index have increased on the Loess Plateau (Li et al. 2017, 2015). These increases in vegetation cover could be attributed to planting of Robinia pseudoacacia on the Loess Plateau.
R. pseudoacacia is a fast-growing species and widely planted although it is sometimes considered an invasive species (Vítková et al. 2017). R. pseudoacacia is a useful species for reforestation and ecological restoration, and is widely planted throughout China since the 1970s for soil and water conservation. Several studies have been carried out on improving soil moisture under R. pseudoacacia plantations (Zhang et al. 2003), reducing soil water depletion, enhancing root distribution (Zhong et al. 2006), improving soil microbial diversity (Zhang et al. 2008), and increasing soil nutrients (Ji et al. 2012; Qiu et al. 2010). The species has the ability of nitrogen fixation by bacteria through root nodules and is easily propagated (Vítková et al. 2017). It uses nutrients from senesced leaves to grow in nutrient-poor soils. Although R. pseudoacacia has been studied by numerous researchers (Cierjacks et al. 2013; Kowarik and Schepker 1998; Rice et al. 2004), most studies have focused on its economic value rather than its ecological function (Vítková et al. 2017). Relatively little attention has been given to strategies for enhancing interactions among soil nutrients, and green and senesced leaves to improve soil moisture.
Foliar resorption is a common process to avoid loss of nutrients, an important mechanism for nutrient conservation and a strategy for adapting to low nutrient environments (Aerts 1996). Nutrient resorption plays a critical role in the development of different soil environments, making vegetation less dependent on soil nutrients (Tang et al. 2013). This resorption mechanism also affects plant growth and the limitation of nutrients. Previous studies have shown that plant species, climate and soil conditions were the main factors affecting nutrient levels of green and senesced leaves (Chen et al. 2013; Reich and Oleksyn 2004; Tang et al. 2013). At a regional level, green leaf nutrients show a biogeographical pattern which also influence the distribution of nutrient resorption efficiency (Yuan and Chen 2009). N and P (phosphorous) resorption efficiencies were significantly related with latitude and climate factors (Yuan and Chen 2009). A global study reported that green leaf nutrients and their resorption efficiency significantly increased with latitude (Vergutz et al. 2012).
Regarding the effect of latitude on soil and plant nutrients (Reich and Oleksyn 2004), the resorption efficiency of nutrients might be used to evaluate the potential for sustainable regional Robinia pseudoacacia plantations based on changes in the contents in green and senesced leaves. A specific objective was to examine the distribution of C, N and P through the soil–green leaf–senesced leaf system and evaluate the following hypotheses: (1) C, N and P contents in the soil would show more variability compared to green and senesced leaf samples along a latitudinal gradient; and, (2) the resorption efficiency of Robinia pseudoacacia would increase along with an increase in latitude across the northern Loess Plateau.
Material and methods
Study area
The Loess Plateau (ca. 640,000 km2), one of the world’s most seriously eroded area, is located along the upper and middle catchments of the Yellow River (Fu et al. 2000). From south to north, mean annual precipitation (MAP) varies from around 800 mm to approximately 100 mm. Along a latitude gradient, mean annual temperature (MAT) decreases from 15 to–7 °C (Zhao et al. 2013). In response to the implementation of China′s “Green for Grain” project, numerous plantations have been established, including ones using R. pseudoacacia. Therefore, we selected 20-year-old R. pseudoacacia plantations from north to south in 14 counties on the Loess Plateau, northwest Shaanxi province (Fig. 1). The study area is within a typical temperate zone (34°42′39.79″–38°48′35.17″N, 108°40′31.6″–110°21′54.57″E). Three sampling sites were selected in each county as replicates.

Geographic information of sampling sites on the Loess Plateau
Soil sampling
All the sampling sites were > 10 km from settlements to eliminate anthropogenic disturbances. In each site, one 20 m × 20 m plot was used for sampling. Ten soil cores (10 cm) were taken in each plot to form a comprehensive soil sample. Stones, roots, decayed materials were removed and the soil samples sieved through 0.15-mm sieve for the analysis of soil C, N, and P.
Green leaf and senesced leaf sampling
Five healthy R. pseudoacacia trees of similar height and age were randomly chosen in each plot. Green leaves were picked from the east, north, west and south sides of the crowns, with an evenly distributed amount from each direction. Green leaves from five trees on each site were mixed into one paper bag. Green leaves were collected mid-August when biomass peaked with the highest levels of nutrients (Zeng et al. 2016b). At the end of the growing season, senesced leaves were collected from the same tree and site (Delgado-Baquerizo et al. 2016).
Measurement of total C, N, and P
Green and senesced leaves were oven-dried at 65 °C to a consistent weight, and ground with a ball mill. Total N in plants and soils were determined by the Kjeldahl acid-digestion method (Sparks et al. 1996). Plant and soil samples were digested with H2SO4 and H2O2 and the P content in plant material determined by a spectrophotometer at 700 nm (Thomas et al. 1967) and in soil samples by a spectrophotometer (Parkinson and Allen 1975). A modified Mebius method was used to measure the organic carbon contents (Nelson and Sommers 1982).
Statistical analysis
The resorption efficiency (RE) was calculated using Eq. (1) (Zeng et al. 2016b):
The statistical package for the social sciences (SPSS 20.0) was used for statistical analyses. The ratio of the standard deviation to the mean of each variable expressed variations of C, N and P. General linear models were used to study the relationships between levels of C, N and P, the resorption efficiency, and latitude. Pearson relation analysis was determined to explore the relations between C, N and P contents in soil, and in green and senesced leaves.
Results
C, N and P contents in soil, and in green and senesced leaves
Latitude had significant effects on soil C and N (p < 0.05) but not on P (Fig. 1). Soil C, N and P levels ranged from 2.4 to 24.6 g kg−1, 0.2 to 2.1 g kg−1, and 0.5 to 1.8 g kg−1, respectively. Average C, N and P levels were 10.8 g·kg−1, 1.1 g·kg−1 and 1.2 g kg−1, respectively (Table 1). Higher coefficients of variation were observed in soil C (69%) and soil N (63%). The distribution of soil C and total N were spatially consistent, declining linearly with an increase in latitude (p < 0.05). The association between latitude and soil P content was significant (p < 0.05).
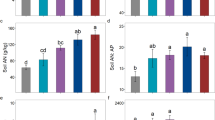
Green leaf C, N, and P levels (g kg−1) ranged from 429 to 485 g kg−1, 18 to 27 g kg−1, and 1.9 to 3.0 g kg−1, respectively. The coefficients of variation for C, N and P were 4%, 12% and 15%, respectively. The CV for green leaf P (15%) was higher than that of green leaf N (12%). Senesced leaf C, N, and P contents were 149–216 g kg−1, 15–21 g kg−1, and 1.4–2.1 g kg−1, respectively, and the coefficients of variation were 11%, 11% and 10%, respectively. Overall, the coefficients of variation for soil nutrients were greater than for plant nutrients. There were no significant relationship of green leaf C, N, P and senesced leaf C to latitude. Senesced leaf N and P levels significantly decreased linearly with an increase in latitude (p < 0.05) (Fig. 2).

Linear regressions of C, N and P contents in soil and green and senesced leaves to latitude
Association between soil and plant C, N and P levels
Pearson correlation analysis revealed that soil C was significantly related with soil N levels (Table 2). Correlation between carbon content and green leaf N content was negative (p < 0.05). Phosphorous levels in senesced leaves were significantly related with soil C and N. Soil N was negatively correlated with green leaf N and senesced leaf P levels (p < 0.05). Correlations between senesced leaf and soil C, N and P levels were insignificant.
Resorption efficiency for N and P by latitude
N resorption efficiency ranged from 7 to 34% and was positively related to latitude (Fig. 3), and P resorption efficiency ranged from 20 to 45%, but the relationship with latitude was insignificant. Average resorption efficiency for N and P were 20% and 23%, respectively. Soil N was significantly related with N resorption efficiency and green leaf N content, however, the correlation between soil N, P resorption efficiency, and green leaf P content were insignificant.

Associations between resorption efficiency, latitude and N content in soil, and between N content in green and senesced leaves and soil nitrogen; RN, N resorption efficiency; RP, P resorption efficiency
Discussion
Variations of C, N and P levels with latitude
The data are partially in accord with the first hypothesis that soil C and N levels (except P) are significantly decreased with increasing latitude, which has also been observed on the Loess Plateau by Deng et al. (2014). Sources of soil nitrogen are mainly derived from organic matter through litter decomposition (Dang et al. 2007). Decomposition and plant productivity processes are closely related with climate and thereby influence the accumulation and distribution of soil organic matter (SOM) and N levels (Yang et al. 2013). On the Loess Plateau, average annual rainfalls and temperatures decrease from south to north (Zhao et al. 2013), which results in slower litter decomposition rates in northern regions. This results in decreasing SOM and plant productivity from south to north. The coefficient of variation for phosphorous was lowest in terms of spatial variation compared to soil C and total N. Being relatively immobile, phosphorous is not easily leached out from soils and the spatial distribution of total P in soil is homogeneous. There were a large number of factors (e.g., parent material, organisms and geochemical processes) affecting soil P content. Climate factors such as temperature and rainfall may play an important role in promoting P leaching and its loss in soil (Sharpley et al. 1997; Simard et al. 2000). However, this study did not find that latitude affected soil phosphorous levels (Fig. 2). On the Loess Plateau, soil parent material may be the main sources of phosphorous (Mage and Porder 2013). Weathering and subsequent leaching of parent minerals resulted in the sources of soil P (Walker and Syers 1976).
Soil C and N are generally high in hot and humid regions because of high plant productivity and low P levels due to leaching (Veldkamp et al. 2008). Reich and Oleksyn (2004) indicated that phosphorous was the main limiting element in soils of the tropics; however, in the present study, soil P levels were relatively stable.
Latitude influences climate, depending on the distance from the equator and this is because the poles (90° N and 90° S) receive less sunlight than the equator. The higher the latitude, the lower the temperature. In the sites in this study, changes of soil total C and N may be attributed to hydrothermal conditions. Temperature and rainfall decrease with increase in latitude. However, the soil undergoes a transformation from “loess” to “sandy loess” beyond a specific latitude (Zeng et al. 2016a). Increasing proportions of sand in the soil with latitude decreases the soil’s nutrient contents.
Variations of C, N and P in green and senesced leaves along a latitude gradient
The data show that green leaf C (454 mg g−1) was slightly lower than the global level (464 mg g−1) (Elser et al. 2000), but N (21.36 mg g−1) and P (2.08 mg g−1) levels were slightly higher than the global level (20.6 mg g−1 and 2.0 mg g−1). This may be attributed to climatic and hydrothermal conditions on the Loess Plateau.
Within ecosystems, the relative inputs of above- and below-ground biomass strongly control the quantity and quality of dead leaves being decomposed (He et al. 2016). This determines the dynamics of labile organic matter and associated nutrient release in the ecosystem which contributes to plant nutrition (Freschet et al. 2013). Therefore, senesced leaves play important roles in the soil–plant interface. N and P are important elements that can limit plant growth as they are involved in basic physiological and biochemical processes. With increasing latitude, R. pseudoacacia senesced leaf N and P significantly decreased but no variation was observed in C levels. Precipitation, temperature and soil type may cause these variations. The soil on the Loess Plateau in Shaanxi Province is sandy loess and nutrient levels gradually decrease from south to north. Sandy loess is prone to wind and water erosion and does not adequately preserve nutrients, leading to a poor growth environment for R. pseudoacacia in the northern areas on the plateau. With decreasing soil nutrients, the senesced leaf N and P contents decline with increasing latitude.
Green leaf nitrogen levels were relatively stable as a result of higher resorption efficiency in higher latitudes. These levels were negatively correlated with soil N, explained by the higher resorption efficiency and the function of N fixation for R. pseudoacacia (Boring and Swank 1984). Nutrient resorption efficiency is a conservation mechanism, especially in lower nutrient environments (Aerts 1996). Increasing N resorption efficiency resulted in the stability of green nitrogen contents which is in agreement with the results of Yuan and Chun (2009). Generally, plants in lower nutrient environments have higher resorption efficiencies (Vitousek 1982) and lower nutrient concentrations in senesced leaves (Killingbeck 1996). In the present study, green leaf nitrogen levels were negatively related to N resorption efficiency, suggesting that the green leaves had a higher nutrient status associated with a lower resorption efficiency (Kobe et al. 2005). In this study, N resorption efficiency (20%) was lower than reported by Han et al. (2013). They reported that nitrogen fixing species such as R. pseudoacacia had lower nitrogen resorption efficiency than other species. Soil nutrient levels also influence plant nutrient resorption efficiency (Tang et al. 2013; Yuan et al. 2005). The significant correlation between soil N and N resorption efficiency indicates that soil nitrogen levels affect green leaf N content.
Soil N levels in this study were lower than the average for China, which suggests that the growth of R. pseudoacacia on the Loess Plateau was limited by nitrogen. More precisely, in higher latitude areas with lower soil N, nitrogen resorption efficiency and the ability of N fixation to meet nutrient demands was enhanced. This adaptive strategy resulted in a linear decrease of nitrogen levels in senesced leaves. Senesced leaf N contents declined with an increase in latitude. Thus, this result would provide useful insights into nutrient limitation strategy for reforestation under different latitudes.
Conclusions
This study indicates that changes in soil–plant associations are influenced by climate and the soil environment. Spatial variations in soil C and N, senesced leaf N and P contents declined significantly with an increase in latitude. Latitude had no effect on green leaf C, N and P levels. However, latitude affected green leaf nitrogen resorption efficiency with lower soil nitrogen, showing that it significantly affected green leaf nitrogen levels. These results highlight the importance of resorption efficiency in regulating R. pseudoacacia plantations over different latitudes on the Loess Plateau.
References
Aerts R (1996) Nutrient resorption from senescing leaves of perennials: are there general patterns? J Ecol 84:597–608
Boring L, Swank W (1984) The role of black locust (Robinia pseudo-acacia) in forest succession. J Ecol 72:749–766
Chen YH, Han WX, Tang LY, Tang ZY, Fang JY (2013) Leaf nitrogen and phosphorus concentrations of woody plants differ in responses to climate, soil and plant growth form. Ecography 36:178–184
Cierjacks A, Kowarik I, Joshi J, Hempel S, Ristow M, Lippe M, Weber E (2013) Biological flora of the British Isles: Robinia pseudoacacia. J Ecol 101:1623–1640
Dang YA, Li SH, Wang GL, Shao MA (2007) Distribution characteristics of soil total nitrogen and soil microbial biomass nitrogen for the typical types of soils on the Loess Plateau. Plant Nutr Fertil Sci 13:1020–1027
Deng L, Shangguan ZP, Sweeney S (2014) “Grain for Green” driven land use change and carbon sequestration on the Loess Plateau, China. Sci Rep 4:1–8
Delgado-Baquerizo M, Reich PB, García-Palacios P, Milla R (2016) Biogeographic bases for a shift in crop C: N: P stoichiometries during domestication. Ecol Lett 19:564–575
Elser J, Sterner RW, Gorokhova EA, Fagan W, Markow T, Cotner JB, Harrison J, Hobbie SE, Odell G, Weider L (2000) Biological stoichiometry from genes to ecosystems. Ecol Lett 3:540–550
Freschet GT, Cornwell WK, Wardle DA, Elumeeva TG, Liu WD, Jackson BG, Onipchenko VG, Soudzilovskaia NA, Tao JP, Cornelissen JH (2013) Linking litter decomposition of above- and below-ground organs to plant–soil feedbacks worldwide. J Ecol 101:943–952
Fu BJ, Chen LD, Ma KP, Zhou HF, Wang J (2000) The relationships between land use and soil conditions in the hilly area of the loess plateau in northern Shaanxi, China. CATENA 39:69–78
Han WX, Tang LY, Chen YH, Fang JY (2013) Relationship between the relative limitation and resorption efficiency of nitrogen vs phosphorus in woody plants. PLoS One 8(12):e83366
He ZM, Yu ZP, Huang ZQ, Davis M, Yang YS (2016) Litter decomposition, residue chemistry and microbial community structure under two subtropical forest plantations: a reciprocal litter transplant study. Appl Soil Ecol 101:84–92
Ji JN, Kokutse N, Genet M, Fourcaud T, Zhang ZQ (2012) Effect of spatial variation of tree root characteristics on slope stability. A case study on Black Locust (Robinia pseudoacacia) and Arborvitae (Platycladus orientalis) stands on the Loess Plateau. China Catena 92:139–154
Killingbeck KT (1996) Nutrients in senesced leaves: keys to the search for potential resorption and resorption proficiency. Ecology 77:1716–1727
Kobe RK, Lepczyk CA, Iyer M (2005) Resorption efficiency decreases with increasing green leaf nutrients in a global data set. Ecology 86:2780–2792
Kowarik I, Schepker H (1998) Plant invasions in northern Germany: human perception and response. In: Starfinger U, Edwards K, Kowarik I, Williamson M (eds) Plant invasions: ecological mechanism and human response. Backhuys Publishers, Leiden, pp 109–120
Li JJ, Peng SZ, Li Z (2017) Detecting and attributing vegetation changes on China’s Loess Plateau. Agric For Meteorol 247:260–270
Li SS, Yang SN, Liu XF, Liu YX, Shi MM (2015) NDVI-based analysis on the influence of climate change and human activities on vegetation restoration in the Shaanxi-Gansu-Ningxia Region, Central China. Remote Sens 7:11163–11182
Mage SM, Porder S (2013) Parent material and topography determine soil phosphorus status in the Luquillo Mountains of Puerto Rico. Ecosystems 16:284–294
Nelson DW, Sommers LE (1982) Total carbon, organic carbon, and organic matter. Methods of soil analysis. Part 2. Chemical and microbiological properties, pp 539–579
Parkinson J, Allen S (1975) A wet oxidation procedure suitable for the determination of nitrogen and mineral nutrients in biological material. Commun Soil Sci Plant Anal 6:1–11
Qiu LP, Zhang XC, Cheng JM, Yin XQ (2010) Effects of black locust (Robinia pseudoacacia) on soil properties in the loessial gully region of the Loess Plateau, China. Plant Soil 332:207–217
Reich PB, Oleksyn J (2004) Global patterns of plant leaf N and P in relation to temperature and latitude. Proc Natl Acad Sci USA 101:11001–11006
Rice SK, Westerman B, Federici R (2004) Impacts of the exotic, nitrogen-fixing black locust (Robinia pseudoacacia) on nitrogen-cycling in a pine–oak ecosystem. Plant Ecol 174:97–107
Sharpley A, Rekolainen S (1997) Phosphorus in agriculture and its environmental implications, Phosphorus loss from soil to water. Proceedings of a workshop, Wexford, Irish Republic, 29–31 September 1995. CAB International, pp. 1–53
Simard R, Beauchemin S, Haygarth P (2000) Potential for preferential pathways of phosphorus transport. J Environ Qual 29:97–105
Sparks D, Page A, Helmke P, Loeppert RS, Tabatabaei P, Johnson M, Sumner C (1996) Methods of soil analysis. Part 3-Chemical methods. Soil Science Society of America Inc, Madison
Sun WY, Song XY, Mu XM, Gao P, Wang F, Zhao GJ (2015) Spatiotemporal vegetation cover variations associated with climate change and ecological restoration in the Loess Plateau. Agric For Meteorol 209–210:87–99
Tang LY, Han WX, Chen YH, Fang JY (2013) Resorption proficiency and efficiency of leaf nutrients in woody plants in eastern China. J Plant Ecol 6:408–417
Thomas R, Sheard R, Moyer J (1967) Comparison of conventional and automated procedures for nitrogen, phosphorus, and potassium analysis of plant material using a single digestion. Agron J 59:240–243
Veldkamp E, Purbopuspito J, Corre MD, Brumme R, Murdiyarso D (2008) Land use change effects on trace gas fluxes in the forest margins of Central Sulawesi Indonesia. J Geophys Res Biogeosci. https://doi.org/10.1029/2007JG000522
Vergutz L, Manzoni S, Porporato A, Novais RF, Jackson RB (2012) Global resorption efficiencies and concentrations of carbon and nutrients in leaves of terrestrial plants. Ecol Monogr 82:205–220
Vítková M, Müllerová J, Sádlo J, Pergl J, Pyšek P (2017) Black locust (Robinia pseudoacacia) beloved and despised: A story of an invasive tree in Central Europe. For Ecol Manage 384:287–302
Vitousek P (1982) Nutrient cycling and nutrient use efficiency. Am Nat 119:553–572
Walker T, Syers JK (1976) The fate of phosphorus during pedogenesis. Geoderma 15:1–19
Yang QL, Wang XJ, Shen YY, Philp J (2013) Functional diversity of soil microbial communities in response to tillage and crop residue retention in an eroded loess soil. Soil Sci Plant Nutr 59:311–321
Yuan ZY, Li LH, Han XG, Huang JH, Wan SQ (2005) Foliar nitrogen dynamics and nitrogen resorption of a sandy shrub Salix gordejevii in northern China. Plant Soil 278:183–193
Yuan ZY, Chen HY (2009) Global scale patterns of nutrient resorption associated with latitude, temperature and precipitation. Glob Ecol Biogeogr 18:11–18
Zeng QC, Dong YH, An SS (2016a) Bacterial community responses to soils along a latitudinal and vegetation gradient on the Loess Plateau. China PloS one 11:e0152894
Zeng QC, Li X, Dong YH, An SS, Darboux F (2016b) Soil and plant components ecological stoichiometry in four steppe communities in the Loess Plateau of China. CATENA 147:481–488
Zethof JHT, Cammeraat ELH, Nadal-Romero E (2019) The enhancing effect of afforestation over secondary succession on soil quality under semiarid climate conditions. Sci Total Environ 652:1090–1101
Zhang HH, Tang M, Chen H, Du XG (2008) Diversity of soil microbial communities in the mycorrhizosphere of five afforestation tree species in the Loess Plateau. J Beijing For Univ 30:85–90
Zhang SQ, Wang GD, Liu JJ, Guo MC (2003) Soil hydro-physical properties of Robinia pseudoacacia plantation forestland in Loess Plateau. J Northwest For Univ 19:11–14
Zhao GJ, Mu XM, Wen ZM, Wang F, Gao P (2013) Soil erosion, conservation, and eco-environment changes in the loess plateau of China. Land Degrad Dev 24:499–510
Zheng XZ, Lin C, Guo BL, Yu JH, Ding H, Peng SY, Sveen TR, Zhang YS (2020) Effects of re-vegetation restoration on soil bacterial community structure in degraded land in subtropical China. Euro J Soil Biol 98:103184
Zhong Z, Chen XR, Xue WP, Wang DH, Yuan ZF (2006) Difference of fine root vertical distribution of Robinia pseudoacacia under the different climate regions in the Loess Plateau. Scientia Silvae Sinicae 42:1–7
Acknowledgements
We thank Zhijie Wang for creating the site map.
Author information
Authors and Affiliations
Corresponding author
Additional information
Corresponding editor: Yanbo Hu.
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Project funding: This study was supported by the National Natural Science Foundation of China (41907051 and 41671280), International Partnership Program of Chinese Academy of Sciences (161461KYSB20170013) and China Postdoctoral Science Foundation (219M662678).
The online version is available at http://www.springerlink.com.
Rights and permissions
About this article

Cite this article
Zeng, Q., Liu, Y., Yang, J. et al. Nutrients of green and senesced leaves of a Robinia pseudoacacia plantation along a latitudinal gradient on the Loess Plateau, China. J. For. Res. 32, 2499–2506 (2021). https://doi.org/10.1007/s11676-020-01273-w
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s11676-020-01273-w