
Abstract
Leaf functional traits are adaptations that enable plants to live under various environmental conditions. This study aims to determine the differences in leaf functional traits among plants grouped by growth habit, leaf life span, leaf lifestyle, leaf form, and origin. Specific leaf area (SLA) of perennial or evergreen species was lower than that of annual or deciduous species because longer-lived leaves of perennial or evergreen species require more investment in structural integrity and/or defense against disturbances, especially with any resource constraint. SLA of large individuals was lower than that of small individuals. The low SLA in large individuals can improve their response to changing light and water conditions because increasing plant height is advantageous for light competition, but it can also impose a cost in terms of structural support and water transport. Petioles of plants with compound leaves were significantly longer than those of simple leaves because branching is expensive in terms of gaining height. SLA of plants increased with increasing invasiveness accordingly, and SLA of invasive plants was higher than that of their native congeners because invasive plants should invest more biomass on leaf growth rather than leaf structures per unit area to achieve a higher growth rate. Overall, variation in leaf functional traits among different groups may play an adaptive role in the successful survival of plants under diverse environments because leaf functional traits can lead to pronounced effects on leaf function, especially the acquisition and use of light. Plant species with different growth and leaf traits balance resource acquisition and leaf construction to minimize trade-offs and achieve fitness advantages in their natural habitat.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
Since leaves are exposed and sensitive to environmental changes, the response of leaf functional traits to such changes could enable plants to occupy a wide variety of environmental conditions (Poorter et al. 2009; Campitelli and Stinchcombe 2013). Thus, leaf functional traits can offer a link between various environmental factors and leaf function (Legner et al. 2014; Xiao et al. 2015; Wang et al. 2016).
Normally, leaf functional traits may vary by plant species and groups because species and groups vary in their tolerance to various environmental selection pressures; thus, leaf functional trait differences among plants with different growth and leaf types can reflect their successfully adaptive strategy (Poorter et al. 2009; Scheepens et al. 2010; Campitelli and Stinchcombe 2013). Hence, determination of the differences in leaf functional trait among these different plant types is important in illuminating the mechanism underlying the ecological strategy of plant species to occupy different habitats successfully.
Generally, specific leaf area (SLA, defined as investment per unit of light capture surface deployed) is one of the most important indices of leaves because it can be used as an indicator of the resource-use strategy of plants (Poorter et al. 2009; Scheepens et al. 2010; Pietsch et al. 2014). In particular, high SLA indicates high resource acquisition and use efficiency with low investment in leaf construction and protective tissues (Poorter et al. 2009; Scheepens et al. 2010; Pietsch et al. 2014). Petiole length, leaf size (indicated by leaf length and leaf width), and leaf shape index (calculated as the ratio of leaf length to leaf width) are also important indices of leaf functional traits because these indices can also be used as indicators of resource-use strategy of plants (Poorter et al. 2009; Wang and Zhang 2012; Soudzilovskaia et al. 2013).
This study aims to determine the differences in leaf functional traits among different groups of plant species in Zhenjiang, China and then assess the role that the main factors played in successful ecological strategies. In particular, leaf functional traits, including petiole length, leaf length, leaf width, leaf shape index, and SLA of 50 plant species grouped according to different traits (growth habit, leaf life span, leaf lifestyle, leaf form, and origin) were assessed to provide insights into the resource-use strategy of these plant species.
Materials and methods
Experimental design
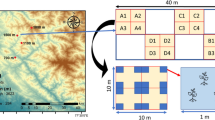
The leaf samples of the 50 plant species (the names and characteristics of the 50 plant species are shown in Table 1) were collected from Zhenjiang having subtropical, humid climate. The annual mean temperature of the area is approximately 15.6 °C, and its monthly mean temperature reaches a maximum of 25 °C in July and decreases to a minimum of −1 °C in January. The annual precipitation is approximately 1088 mm. The 50 species were the common plants in the surveyed sample sites. The 50 species were divided into different groups based on their life form (four categories: tree, shrub, liana, and herb), leaf life span (two categories: annual and perennial), leaf lifestyle (two categories: evergreen and deciduous), leaf form (two categories: simple and compound), and origin (three categories: native, alien, and invasive) (Table 1). Sixteen adult plant samples for each species were collected randomly. At least five fully expanded and intact leaves of one plant sample were selected randomly to determine the functional traits.
Determination of leaf functional traits
Four indices of leaf functional traits (petiole length, leaf shape index, leaf length and width, and SLA) of the fifty plant species were determined. Petiole length was determined by ruler measurement (Xiao et al. 2015; Wang et al. 2016). Leaf shape index was calculated as the ratio of leaf length to the corresponding leaf width (Wang and Zhang 2012; Xiao et al. 2015; Wang et al. 2016). Leaf length, the maximum value along the midrib, and leaf width, the maximum value perpendicular to the midrib (Wang and Zhang 2012) were measured using a ruler (Xiao et al. 2015; Wang et al. 2016). SLA was computed using the ratio of the leaf area to the corresponding leaf dry mass (cm2 g−1) according to previous studies (Scheepens et al. 2010; Xiao et al. 2015; Wang et al. 2016).
Statistical analyses
Data were evaluated to determine the deviations from normality and homogeneity of variance before analysis. Differences among various dependent variables were assessed using an analysis of variance between groups followed by multiple comparisons using the S–N–K test when appropriate. Variance analysis for the differences in the leaf functional traits among different plant groups were assessed based on the classification of the 50 species surveyed in this study. Statistically significant differences were set at P ≤ 0.05. Correlation patterns among various dependent variables were determined by correlation analysis using SPSS Statistics (version 22.0; IBM, Armonk, NY, USA).
Results
Leaf functional traits of different plant groups
Differences were observed among leaf functional traits among the 50 species (Table 2). In particular, mean petiole length of trees, lianas, and herbs was 48.04 % (P < 0.05), 55.09 % (P < 0.05), and 60.04 % (P < 0.05), respectively, higher than those of shrubs (Table 3). Mean leaf length of trees was 19.64 % (P < 0.05) and 32.44 % (P < 0.05) higher than those of shrubs and lianas, respectively; leaf length of herbs was 20.09 % (P < 0.05) and 37.87 % (P < 0.05) higher than those of shrubs and lianas, respectively (Table 3). Mean leaf width of shrubs, lianas, and herbs was 40.79 % (P < 0.05), 28.21 % (P < 0.05), and 37.74 % lower (P < 0.05) than those of trees (Table 3). Mean leaf shape index of plants with different life forms significantly decreased in the following order: herbs, shrubs, trees, and lianas (Table 3). The mean SLA of lianas was 23.23 % (P < 0.05) and 21.66 % (P < 0.05) higher than those of trees and shrubs, respectively; the mean SLA of herb was 25.64 % (P < 0.05) and 24.02 % (P < 0.05) higher than those of trees and shrubs, respectively (Table 3).
Mean leaf length, leaf shape index, and SLA of annual species were 26.10 % (P < 0.05), 41.67 % (P < 0.05), and 17.97 % (P < 0.05) higher than those of perennial species, respectively (Table 3). No significant difference (P > 0.05) was found for mean petiole length and leaf width between the annual plants and perennial ones (Table 3).
Mean leaf width and SLA of deciduous plants were 23.76 % (P < 0.05) and 9.05 % (P < 0.05) higher than those of evergreen ones, respectively, whereas mean leaf shape index of deciduous plants was 23.61 % (P < 0.05) lower than that of evergreen ones (Table 3). Mean petiole length and leaf length were not significantly different (P > 0.05) between deciduous plants and evergreen ones (Table 3).
Mean leaf length and width of plants with simple leaves were 56.06 % (P < 0.05) and 60.58 % (P < 0.05) higher than those with compound, respectively, while mean petiole length and SLA of plants with simple leaves were 13.06 % (P < 0.05) and 22.10 % (P < 0.05) lower than those with compound, respectively (Table 3). The difference in mean leaf shape index between plants with simple and compound leaves was not significant (P > 0.05) (Table 3).
Mean petiole length of plants with different origins decreased significantly (P < 0.05) in the following order: invasive, native, and alien (Table 3) but the order was the reverse for mean leaf length (P < 0.05) (Table 3). Mean leaf width of invasive plants was 50.85 % (P < 0.05) and 57.89 % (P < 0.05) lower than those of alien and native, respectively (Table 3). Mean leaf shape index of invasive plants was 15.49 % (P > 0.05) and 23.91 % (P < 0.05) higher than those of alien and native, respectively (Table 3). The mean SLA of invasive plants was 19.37 and 25.42 % higher than those of alien and native, respectively (Table 3).
Relationships between the indices of leaf functional traits
Correlation patterns between leaf functional traits among different plant groups were assessed with a correlation analysis (Table 4). In particular, petiole length was positively (P < 0.0001) correlated with leaf length, leaf width, and SLA, but negatively (P < 0.05) correlated with leaf shape index (Table 4). Leaf length was positively (P < 0.0001) correlated with leaf width and leaf shape index (Table 4). Leaf width was negatively (P < 0.0001) correlated with leaf shape index (Table 4). Leaf shape index was negatively (P < 0.01) correlated with SLA (Table 4).
Discussion
Perennial or evergreen species always produce leaves that have a longer leaf life span than annual or deciduous species. Thus, the leaves of perennial or evergreen species had lower SLA and higher tensile strength than in those of annual or deciduous species, and the leaves of trees and shrubs also generally had a lower SLA and higher tensile strength than those of lianas and herbs (Xu et al. 2014; Silva et al. 2015). The results of this study are consistent with those previous studies, i.e., the SLA of perennial or evergreen species was lower than that of annual or deciduous species (Xu et al. 2014; Silva et al. 2015), and the SLA of large individuals was lower than that of small individuals (Liu et al. 2010). The longer life span of leaves for perennial or evergreen species requires more investment in structural integrity and/or defense against disturbances, especially with resource constraints (Liu et al. 2010; Cianciaruso et al. 2013; Silva et al. 2015). Hence, perennial or evergreen species allocate more biomass to structural rather than metabolic components. The reason that the SLA of trees and shrubs was lower than that of lianas and herbs may be due to the higher photosynthetic capacity of their leaves at a given investment (e.g., SLA, N and phosphorus contents) because their canopies can intercept more light than small individuals can (Liu et al. 2010). The low SLA in large individuals can improve their response to changing light and water conditions (Liu et al. 2010; Silva et al. 2015) because their greater height can serve as a strategy for light competition, although it can also impose a cost in terms of structural support and water transport (Ishii and Asano 2010). The SLA also often decreases with increasing light availability (Liu et al. 2010). Thus, the SLA-leaf life span spectrum is a trade-off between the potential rate of return per leaf area per unit time and the duration of return for plant species to achieve the maximum individual resources possible, ultimately leading to maximum fitness (Cianciaruso et al. 2013).
Large twigs may enhance the competitive potential of plants because they minimize branching and thereby facilitate sapling growth to the unshaded overstory (Xiang et al. 2009). The petiole mass disproportionally increased with increasing lamina mass and twig mass, and the allometric relationship between petiole mass and twig mass implied that the costs of supporting the lamina increased with increasing twig size (Xiang et al. 2009). Thus, the biomass allocation to petiole for plants with compound leaves may be significantly higher than those with simple leaves. The result of this study showed that petiole length of plants with compound leaves was significantly higher than those with simple leaves. A previous study also found that lamina support (including petioles, rachis, and petiolules) of compound-leaved species was significantly higher than that of simple-leaved species (Li et al. 2008). The higher biomass allocation of lamina support for compound-leaved species may also provide advantages other than lamina support (Li et al. 2008); for instance, branching is expensive in terms of gaining height because not only is C required for branch extension, but C is also required for additional stem diameter growth to support the wider crown. Although the function of petioles of compound-leaved species is mechanically analogous to that of branches, they are much less costly in terms of C and nutrients (Li et al. 2008). Meanwhile, compound leaves often arrange the small leaflets along the rachis to maximize light interception, which shifts the center of mass of the leaf, thereby increasing the mechanical bending stress (Li et al. 2008). Thus, compound leaves may be an important adaptive feature contributing to minimizing branching and to rapid height gain, particularly in light-demanding tree species (Poorter et al. 2006). Moreover, there was an isometric relationship between leaf mass and twig mass and between lamina mass and twig mass (Li et al. 2008; Xiang et al. 2009). Plants with compound leaves can thereby display high SLA characterized by high resource acquisition and use efficiency with low investment in leaf construction and protective tissues and leading to a higher relative growth rate.
Trait differences between invasive and native species are believed to be closely related to whether the invasives are successful because exotic and native plant species coexisting in the same ecosystem are exposed to similar environmental selection pressures (van Kleunen et al. 2010; Gao et al. 2013). Previous studies found that the SLA of invasive plants is higher than that of their native congeners because invasive plants invest more biomass in leaf growth than in leaf structures per unit area to achieve a higher growth rate (van Kleunen et al. 2010; Hou et al. 2014). Meanwhile, a higher SLA is often highly correlated with a growth advantage for invasive plants over native species (Huang et al. 2016). Thus, the SLA of plants is expected to increase with increasing invasiveness; that is, the SLA of plants with different origins increases in the following order: native, alien, and invasive (Smith and Knapp 2001). The result of the present study is consistent with this hypothesis, although there was no significant difference in SLA values between native and alien species. The higher SLA for invasive species can enable them to more efficiently use energy, facilitating plant growth (Hou et al. 2014; Huang et al. 2016).
Numerous studies have shown that leaf size was positively correlated with SLA because leaves with a high SLA have low structural investment, but leaves with low SLA likely invest more biomass into producing more leaf structures (Poorter et al. 2009; Soudzilovskaia et al. 2013; Pietsch et al. 2014). However, unlike these previous studies, SLA and leaf size were not significant correlated in the present study. Empirical studies have also provided conflicting results, with observed correlations between leaf size and SLA that are positive (Burns and Beaumont 2009), negative (Wang et al. 2016), or unrelated (Xiao et al. 2015). These findings suggest that there is species specificity for the relationship among leaf functional traits. Petiole length and leaf shape index, rather than leaf size, were the important characteristics that led to pronounced effects on SLA. The reason for the positive correlation between petiole length and SLA may be due to the isometric relationship between lamina mass and twig mass, i.e., the biomass allocation to either leaves or laminas was independent of twig mass (Xiang et al. 2009). The negative correlation between leaf shape index and SLA was consistent with a previous study (Wang et al. 2016), perhaps because leptosomic leaves may allocate more biomass to leaf construction, especially because the costs of lamina support increase with increasing leaf shape index.
Conclusion
The purpose of this study was to evaluate differences in leaf functional traits among different plant groups. Results showed that leaf functional traits of plants vary among the different groups. In particular, specific leaf area (SLA) of perennial or evergreen species was lower than that of annual or deciduous species because a longer life span of leaves for perennial or evergreen species requires more investment in structural integrity and/or defense against disturbances, especially with resource constraint. The SLA of large individuals was lower than that of small individuals. The low SLA in large individuals can improve their response to changing light and water conditions because that increase in height is a benefit for light competition; however, it can also be costly in terms of structural support and water transport. Petiole length of plants with compound leaves was significantly higher than those with simple leaves because branching is expensive in terms of gaining height; carbon is required for branch extension and the additional growth in stem diameter to support the wider crown. The SLA of plants increased with increasing invasiveness and was higher than in the invasives than in their native congeners because of their greater investment in leaf biomass rather than leaf structures per unit area to achieve a higher growth rate. The variations in leaf functional traits among different groups may play an adaptive role in the successful survival of these plants under diverse environments because leaf functional traits can have pronounced effects on leaf function, especially the acquisition and use of light. Successful plants in general obtain an optimal trade-off between resource capture and conservation to acquire more resources and achieve as great a fitness advantage as possible in variable environments (Xiao et al. 2015; Wang et al. 2016).
References
Burns KC, Beaumont SAM (2009) Scale-dependent trait correlations in a temperate tree community. Austral Ecol 34:670–677
Campitelli BE, Stinchcombe JR (2013) Natural selection maintains a single-locus leaf shape cline in Ivyleaf morning glory, Ipomoea hederacea. Mol Ecol 22:552–564
Cianciaruso MV, Silva IA, Manica LT, Souza JP (2013) Leaf habit does not predict leaf functional traits in cerrado woody species. Basic Appl Ecol 14:404–412
Gao XM, Zhao YJ, Yang XJ, Sun SC (2013) Linking trait differences to community dynamics: evidence from Eupatorium adenophorum and co-occurring native species during a three-year succession. PLoS One 8:e50247
Hou QQ, Chen BM, Peng SL, Chen LY (2014) Effects of extreme temperature on seedling establishment of nonnative invasive plants. Biol Invasions 16:2049–2061
Huang QQ, Shen YD, Li XX, Li SL, Fan ZW (2016) Invasive Eupatorium catarium and Ageratum conyzoides benefit more than does a common native plant from nutrient addition in both competitive and non-competitive environments. Ecol Res 31:145–152
Ishii H, Asano S (2010) The role of crown architecture, leaf phenology and photosynthetic activity in promoting complementary use of light among coexisting species in temperate forests. Ecol Res 25:715–722
Legner N, Fleck S, Leuschner C (2014) Within-canopy variation in photosynthetic capacity, SLA and foliar N in temperate broad-leaved trees with contrasting shade tolerance. Trees 28:263–280
Li GY, Yang DM, Sun SC (2008) Allometric relationships between lamina area, lamina mass and petiole mass of 93 temperate woody species vary with leaf habit, leaf form and altitude. Funct Ecol 22:557–564
Liu FD, Yang WJ, Wang ZS, Xu Z, Liu H, Zhang M, Liu YH, An SQ, Sun SC (2010) Plant size effects on the relationships among specific leaf area, leaf nutrient content, and photosynthetic capacity in tropical woody species. Acta Oecol 36:149–159
Pietsch KA, Ogle K, Cornelissen JHC, Cornwell WK, Bönisch G, Craine JM, Jackson BG, Kattge J, Peltzer DA, Penuelas J, Reich PB, Wardle DA, Weedon JT, Wright IJ, Zanne AE, Wirth C (2014) Global relationship of wood and leaf litter decomposability: the role of functional traits within and across plant organs. Glob Ecol Biogeogr 23:1046–1057
Poorter L, Bongers L, Bongers F (2006) Architecture of 54 moist-forest tree species: traits, trade-offs, and functional groups. Ecology 87:1289–1301
Poorter H, Niinemets U, Poorter L, Wright IJ, Villar R (2009) Causes and consequences of variation in leaf mass per area (LMA): a meta-analysis. New Phytol 182:565–588
Scheepens JF, Frei ES, Stöcklin J (2010) Genotypic and environmental variation in specific leaf area in a widespread Alpine plant after transplantation to different altitudes. Oecologia 164:141–150
Silva JO, Espírito-Santo MM, Morais HC (2015) Leaf traits and herbivory on deciduous and evergreen trees in a tropical dry forest. Basic Appl Ecol 16:210–219
Smith MD, Knapp AK (2001) Physiological and morphological traits of exotic, invasive exotic and native species in tallgrass prairie. Int J Plant Sci 162:785–792
Soudzilovskaia NA, Elumeeva TG, Onipchenko VG, Shidakov II, Salpagarova FS, Khubiev AB, Tekeev DK, Cornelissen JHC (2013) Functional traits predict relationship between plant abundance dynamic and long-term climate warming. Proc Natl Acad Sci USA 110:18180–18184
van Kleunen M, Weber E, Fischer M (2010) A meta-analysis of trait differences between invasive and non-invasive plant species. Ecol Lett 13:235–245
Wang Z, Zhang L (2012) Leaf shape alters the coefficients of leaf area estimation models for Saussurea stoliczkai in central Tibet. Photosynthetica 50:337–342
Wang CY, Liu J, Xiao HG, Du DL (2016) Response of leaf functional traits of Cerasus yedoensis (Mats.) Yü li to serious insect attack. Pol J Environ Stud 25:333–339
Xiang S, Wu N, Sun SC (2009) Within-twig biomass allocation in subtropical evergreen broad-leaved species along an altitudinal gradient: allometric scaling analysis. Trees 23:637–647
Xiao HG, Wang CY, Liu J, Wang L, Du DL (2015) Insights into the differences in leaf functional traits of heterophyllous Syringa oblata under different light intensities. J For Res 26:613–621
Xu SS, Li Y, Wang GX (2014) Scaling relationships between leaf mass and total plant mass across Chinese forests. PLoS One 9:e95938
Acknowledgments
We are grateful to the anonymous reviewers for insightful and constructive comments that greatly improved this manuscript.
Author information
Authors and Affiliations
Corresponding author
Additional information
Project funding
This work was supported by the National Natural Science Foundation of China (31300343), Natural Science Foundation of Jiangsu Province, China (BK20130500), Jiangsu Collaborative Innovation Center of Technology and Material of Water Treatment, and Research Foundation for Advanced Talents, Jiangsu University (12JDG086).
The online version is available at http://www.springerlink.com
Corresponding editor: Hu Yanbo
Rights and permissions
About this article

Cite this article
Wang, C., Zhou, J., Xiao, H. et al. Variations in leaf functional traits among plant species grouped by growth and leaf types in Zhenjiang, China. J. For. Res. 28, 241–248 (2017). https://doi.org/10.1007/s11676-016-0290-6
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s11676-016-0290-6