
Abstract
We investigated the floristic composition of the Hyrcanian forests and the related forest-steppe ecotone in Northern Iran by using two long ecological transects, from lowland to upper mountain areas. The study was conducted during 2008 and 2009 and yielded the identification of 395 plant taxa belonging to 233 genera and 78 families. Dicots with 300 taxa were the richest groups of the flora, followed by monocots with 75 taxa, pteridophytes with 18 species, and gymnosperms with two species. The largest families were Asteraceae (33 taxa); Rosaceae (32 taxa); and Poaceae (30 taxa), and the most diverse genera included Carex (15 taxa); Alchemilla (7 taxa); and Poa, Geranium and Acer (6 taxa each). Hemicryptophytes were the most dominant life forms in the area (40 %); followed by geophytes (31.4 %); phanerophytes (15.4 %); therophytes (11.4 %); and chamaephytes (1.8 %). Phytogeographically, Euro-Siberian/Irano-Turanian elements (86 taxa, 21.8 %) and Euro-Siberian elements (85 taxa, 21.5 %) were the most common chorotypes in the area. Out of 395 taxa, 66 taxa (16.7 %) were endemics and subendemics in Iran, of which 26 taxa were exclusively endemics of Iran. According to the IUCN Red List Categories, 48 threatened plant taxa were found in the study area. Plant diversity, life form, and chorotypes in the current study were compared with similar transect studies in other areas of the Hyrcanian forests and in different altitudinal belts, using Sørenson similarity indices. Floristic composition of the surveyed transects demonstrated almost 50 % similarity between them.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
Studies on ecology and the distribution of organisms within a long elevational gradient have been considered in many ongoing projects in mountainous ecosystems across the world (McCain and Grytnes 2010), particularly with an emphasis on flora and species attributes (Rahbek 1995; Montana and Valientebanuet 1998; Kessler 2000; Pavón et al. 2000; Grytnes and Vetaas 2002; Wang et al. 2002; Grytnes 2003; Klimeš 2003). The study of floristic composition is considered to be more suited to direct study (Nicholes 1930). Furthermore, knowledge of plant species and their diversity within the long gradient is a prerequisite for any ecological and vegetation investigations, particularly for poorly known areas. With these tools, it is possible to expand conservation and management activities in each area (Jafari and Akhani 2008).
Only the northern section of the Alborz mountain ranges in Iran, that is the Hyrcanian area, is located in the Euro-Siberian phytogeographical region (sensu Zohary 1973; Léonard 1989; Akhani 1998) and includes deciduous forests and forest-steppe ecotones, unlike the arid and semiarid landscapes throughout most of Central and Southern Iran (Naqinezhad et al. 2008). The forests are one of the few remnants of natural closed-canopy deciduous forests in the world (Bobek 1951) and are located in three Northern provinces of Iran, namely, Gilan, Mazandaran and Golestan with approximately 800 km in length, 110 km width and a total surface area of 1.84 million hectares.
In spite of rather long history of studies on the flora and vegetation ecology of the Hyrcanian area (Hamzeh’ee et al. 2008; Siadati et al. 2010; Naqinezhad et al. 2013), floristic studies, particularly within the long lowland-upper mountain forest transects are scarce. A rough, general description of the flora and vegetation of the Hyrcanian area within such transects was already provided by (Mobayen and Tregubov 1970), Zohary (1973) and Frey and Probst (1986). Detailed description of flora, however, is unknown for many parts of this area. Siadati et al. (2010) firstly reported all plants within their ecological transects in the Kheyrud forest, a protected forest in the center of Hyrcanian area.
The purpose of the current investigation is to provide the first floristic inventory of the vascular plants of the two forest and forest-steppe ecotone transects in Gilan and Mazandaran provinces and to determine the life forms, chorotypes, IUCN Red List categories of the plants, and to estimate similarity between all the studied transects. The findings should provide insights into the potential of biodiversity within the Hyrcanian forests and their conservation values, and the changing of floristic composition of the forests in different surveyed transects.
Materials and methods
Study areas
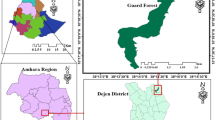
Two well-protected forest lines along an altitudinal gradient from the Caspian Sea to timberline were selected in the Hyrcanian area (Fig. 1). The Rudsar transect close to Rudsar city (Gilan province) ranging from 300 to 2,600 m a.s.l. (50°20′–50°30′E, 37°05′–36°50′N) and, the Savadkuh transect adjacent to Savadkuh city (Mazandaran province), ranging from 300 to 2,300 m a.s.l. (52°45′–53°10′E, 35°55′–36°20′N). The majority of both transects are listed as protected areas by the Iranian Department of the Environment.

Locations of the four ecological transects in the Hyrcanian forest accompanied with climatic curves of two stations near to the studied areas (1 Rudsar transect, 2 Ramsar transect, 3 Kheyrud transect and 4 Savadkuh transect)
In addition to a comparison of floristic composition between the two studied transects here, data from two other earlier transects (i.e., Ramsar transect and Kheyrud transect) were also included in the analyses (Naqinezhad et al. 2010; Siadati et al. 2010). The Ramsar transect (located close to Ramsar city in Mazandaran province) has 63 studied plots, ranging from 300 to 2,300 m a.s.l. (50°31′–50°42′E, 36°48′–36°54′N). The Kheyrud transect, with 117 plots, is located close to Nowshahr city (Mazandaran province), between Savadkuh and Ramsar ranging from 100 to 2,100 m a.s.l. (51°33′–51°39′E, 36°33′–36°45′N) (Fig. 1). The size of plots, elevation interval between the plots, data collection, nomenclature, and other species attributes in these transects were similar to that of recent transects.
Middle Jurassic to Upper Cretaceous limestone formations form very high rock cliffs along the East–West directed thrust fault zones in northern section of the Alborz (Stöcklin 1974). The northern foothills of Alborz are mainly composed of Neogene continental clastic and marine sediments, with some high peaks of Mesosoic limestones along the main faults (Axen et al. 2001; Davidson et al. 2004). The majority of area under 500 m a.s.l. particularly in Rudsar transect, belongs to Quaternary formation which includes old marine deposits, new and old costal deposits and deltaic alluvials. The age of this area is determined to be related to the Pleistocene and Holocene. In upper altitudes, volcanic units of Cretaceous formation are dominant features.
Two climate stations, Ramsar (time period: 1955–2005) and Gharakhil in Ghaemshahr (time period: 1984–2005), are the closest to the studied transects. The mean monthly temperature is the lowest in January and February and the highest in July and August in both stations. Mean total annual precipitation in Ramsar and Gharakhil is 1217.8 and 738.7 mm with the highest precipitation in October (269.6 and 100.6 mm), respectively (Fig. 1). In a recent bioclimatic classification of Iran, the climate of the area varies from Temperate oceanic in the Ramsar site to Mediterranean pluviseasonal-oceanic in the Gharakhil site (Djamali et al. 2011).
The vegetation structure and relevant floristic composition of ecotone places in the two studied transects (upper than 2,000 m a.s.l.) are clearly different. The Rudsar transect harbors species, such as Acer hyrcanum, Quercus macranthera and Viburnum lantana as dominant species accompanied with Geranium purpureum, Lapsana communis, Phuopsis stylosa, Stachys byzantine and Veronica rechingeri in greater than 2,000 m a.s.l. Acer campestre, Berberis integerrima, Pyrus boissieriana are the dominant species in the ecotone area of Savadkuh transect. The latter species are accompanied with Centaurea hyrcanica, Crepis sancta subsp. nemausensis, Saponaria cerastioides, Stellaria holostea, and Veronica mazanderanae in this area.
In contrast to the ecotone area, forest sectors of both transects show relatively similar floristic composition in the same altitudinal bands. In the lowlands (up to 800 m a.s.l.), Parrotia persica, Diospyrus lotus, Alnus subcordata var. villosa, Acer velutinum and Ruscus hyrcanus are more prominent. Furthermore, in the higher altitudes up to 1,900 m a.s.l. Acer cappadocicum, Carpinus betulus, Quercus castaneifolia subsp. castaneifolia, Tilia platyphyllos subsp. caucasica become dominant accompanied by some species like Epipactis persica, Euphorbia amygdaloides, Festuca drymeia, Galium odoratum in the herbal layer. However, the bulk of forests, ranging from 1,000 to 1,900 m a.s.l., are dominated by Fagus orientalis accompanied by Acer velutinum, Laurocerasus officinalis and Vaccinum arctostaphylos, Aruncus vulgaris, Brachypodium sylvaticum, Cardamine impatiens var. pectinata, Carex pendula.
Data collection and analyses
Data collections were performed during 2008 and 2009. Three 20 × 20 m plots in the forest area and three 10 × 10 m plots in the ecotone and upper mountain areas were allocated in each 100 m elevation interval. Selection of plot size was determined by the main dominant physiognomy of vegetation. A total of 122 plots were allocated, of which 68 belonged to Rudsar transect and 54 belonged to Savadkuh transect. The cover of all vascular plant species was estimated using the old Braun-Blanquet scale (r, +, 1–5). The nomenclatures are mainly based on Rechinger (1963–2010), Assadi et al. (1988–2011), Frey et al. (2006) and Khoshravesh et al. (2009).
The distribution data of species were extracted from Flora Iranica (Rechinger 1963–2010), Flora of Turkey (Davis 1965–1988) and mainly from the above-mentioned nomenclature sources. The main concepts of phytochoria are based on Zohary (1973), Takhtajan (1986), and Léonard (1988). We used the following abbreviations in the current paper: ES (plants distributed in the Euro-Siberian region), IT (plants distributed in the Irano-Turanian region), M (plants distributed in the Mediterranian region), PL (pluriregional elements, referring to plants ranging over three phytogeographical regions); and COS (cosmopolitan, referring to plants that have a broad worldwide distribution).
For statistical purposes, four altitudinal belts within the studied transects were defined: lowland belt between 300 and 500 m; submountain belt between 501 and 1000 m; mountain belt between 1001 and 1900 m and upper mountain/ecotone belt, between 1901 and 2600 m. This delimitation is according to the floristic information provided by Zohary (1973), Frey and Probst (1986).
Threatened categories were proposed for the endemic and rare taxa in the study area according to the IUCN risk categories (Jalili and Jamzad 1999; IUCN 2001). The following abbreviations are used: EN, endangered; VU, vulnerable; LR, lower risk; and DD, data deficient.
Sørenson’s similarity indices between the studied transects were determined by using the following formula (Kent and Coker 2002): Ss = 2a/(2a + b + c), where a, number of species common to both stands (A, B); b, number of species only in stand A; and c, number of species only in stand B.
Results
Floristic richness in the two transects
In total, 395 plant taxa belonging to 233 genera and 78 families were determined in the two studied transects. Of the total families, 56 families were dicotyledons, 12 families were monocotyledons, two families were gymnosperms, and eight families were pteridophytes (Appendix 1). A total of 89 plant taxa (22.5 %) were restricted exclusively in the forest areas (located in 300–1,900 m a.s.l.); 231 plant taxa (58.5 %) were exclusively found in the ecotone and upper mountain areas (located between 2,000–2,600 m a.s.l.); and 75 plant taxa (19 %) were shared between two habitats. In total, 306 plant taxa were observed in the ecotone and upper mountain areas (Fig. 2). The most species-rich families in the current study were Asteraceae (33 taxa, 8.4 %), Rosaceae (32 taxa, 8.1 %), Poaceae (30 taxa, 7.6 %), Fabaceae (24 taxa, 6.1 %), and Lamiaceae (21 taxa, 5.3 %), respectively. The genera with the larger number of species were Carex (15 taxa), Alchemilla (7 taxa) and Geranium, Poa and Acer (6 taxa each), respectively (Table 1).

The number of plant taxa within four transects in the Hyrcanian area and in different altitudinal belts
The dominant life forms were hemicryptophytes (158 taxa, 40 %), followed by geophytes (124 taxa, 31.4 %), phanerophytes (61 taxa, 15.4 %), therophytes (45 taxa, 11.4 %), and chamaephytes (7 taxa, 1.7 %) (Fig. 3). Phytogeographically, Euro-Siberian/Irano-Turanian elements (86 taxa, 21.8 %) and Euro-Siberian elements (85 taxa, 21.5 %) were the most common chorotypes in the area. Out of 395 taxa, 66 taxa (16.7 %) were endemic and/or subendemic in Iran, of which 26 taxa were exclusively endemic of Iran. Furthermore, 20 taxa were Iranian Hyrcanian endemic. Eleven taxa (2.8 %) were not identified down to species level in our checklist.

The proportion of different life forms across the four transects
Comparison of the species attributes among the four ecological transects
The number of plant taxa and life form spectrum in the four ecological transects and within four altitudinal bands (50–500, 501–1,000, 1001–1900, 1901–2600 m a.s.l.) are shown in Figs. 2, 3, and 4. In the Rudsar transect, 17 taxa were exclusively found in the level higher than 2,300 m a.s.l. and in the Kheyrud transect, 28 taxa were exclusively collected in the level lower than 300 m. In all transects, hemicryptophytes and geophytes possessed the highest proportions. Moreover, the rate of geophytes were relatively constant in the different altitudinal belts in all transects (Fig. 4).

The proportion of various life forms (%) in different transects and within different altitudinal belts
Calculating the Sørenson similarity indices indicated that the floristic composition of the surveyed transects had almost 50 % similarity between them (Table 2). For a more comparable survey, only the forest parts of all transects were separately evaluated, which showed that Rudsar and Savadkuh had only 64 % similarity in their forestcomponents. A total of 125 taxa in Rudsar and 118 taxa in Savadkuh were collected, of which 78 taxa were common in these two transects. Comparison of the forest parts of the two remaining transects did not show any remarkable departure from the latter overall comparison (Table 2).
IUCN categories and threatened plants
A total of 47 endemic and rare taxa were included in the risk categories defined by IUCN (Table 3). Distribution of the threat categories of these taxa is as follows: three endangered (EN) taxa, six vulnerable taxa (VU), 33 lower risk (LR) taxa and five data deficient (DD) taxa. Out of the total, 16 endemic taxa are in northern Iran (Hyrcanian endemics), five taxa are endemic to Iran (endemics of Irano-Turanian parts of Iran), and five taxa are subendemics. It should be noted that four endemic taxa of northern Iran (Arenaria serpyllifolia var. macrosepala, Cousinia pterocaulos, Ilex spinigera, Scabiosa columbaria subsp. caespitosa), and one endemic taxon of Iran (Crucianella gilanica subsp. demavendensis) have not yet been included in the IUCN categories.
Discussion
The Hyrcanian area represents a region with high species richness (ca. 3200 taxa) (Akhani 1998; Akhani et al. 2010) that also has numerous endemic species, particularly in the upper mountain and alpine areas (Noroozi et al. 2008). Flora of ecotone and upper mountain areas of the studied transects were more heterogeneous (with 306 plant taxa) compared to the forest sectors. Ecotones are frequently encountered in the transitional zone between the Hyrcanian forests in the South Caspian region and their adjacent semi-arid steppes (Djamali et al. 2009; Naqinezhad et al. 2010). These ecosystems are among the most sensitive sections of the biosphere to environmental change and can shift considerably in response to climate change (Neilson 1993).
The occurrence of a high number of Euro-Siberian elements in the surveyed transects indicated the floristic linkage of the study area to the Euro-Siberian phytogeographical region (sensu Zohary 1973; Léonard 1989; Akhani 1998; Jafari and Akhani 2008). The number of Irano-Turanian elements increased with increasing altitude. This is due to the occurrence of a high number of Irano-Turanian elements in the steppe parts of upper mountain areas of Alborz (Klein 1991, 2001; Noroozi et al. 2008). Consequently, this caused a concentration of biregional Euro-Siberian/Irano-Turanian elements in some transects.
The study of plant life forms is important because it provides the basic structural components of vegetation stands (Box 1981). It is also indicative of habitat conditions (Archibold 1995). Hemicryptophytes increased with increasing altitude, particularly above 2,000 m. This was compatible with the fact that hemicryptophytes were related to humid and cold climates in upper altitudes (Raunkiaer 1934).
The high occurrence of geophytes in the forest parts has been previously reported (Akbarinia et al. 2004; Ghahreman et al. 2006; Naqinezhad et al. 2010; Siadati et al. 2010). Likewise, increasing geophytes was considered to be related to increasing of altitudinal bands (Fig. 4). The lower proportion of phanerophytes in the higher altitudes indicated they were susceptible to frost and extreme cold. The lower frequency of this life form across different altitudinal belts and among surveyed transects proved this finding.
The surveyed transects exhibited almost 50 % similarities among them. Contrary to the homogenous flora of the four transects, the similarity of floristic composition was not very prominent. The following reasons provide some explanation for this moderate similarity:
-
1.
Species difference might be related to the different number of plots and relevant altitudinal bands surveyed.
-
2.
Different management regimes in the transects could contribute to discriminatory species adaptation.
-
3.
Differences in climatic features (temperatures and precipitations) between the stations demonstrate, to a degree that there is a clear trend of decreasing precipitation from the west to east of the Hyrcanian area (e.g. Khalili 1973; Akhani et al. 2010).
-
4.
The authors provided supplementary plant collections of the two surveyed transects (Ramsar and Kheyrud) that showed how the forest margins and all surrounding habitats had shifted beyond their original plots (Naqinezhad et al. 2010; Siadati et al. 2010).
The high occurrence of therophytes in the two latter transects (particularly in Kheyrud) makes this claim reasonable. Therophytes are considered indicators for highly stressful habitat factors such as forest roads and margins (Archibold 1995; Naqinezhad et al. 2008).
In a separate account, only forest sectors of the four transects (300–1,900 m) were specifically analyzed. However, similarity indices did not obviously change except for the two transects of Rudsar and Savadkuhthat showed a similarity of 64 %. This might be due to different climatic regimes in the areas.
The low percentage of similarity across the surveyed transects could be due to different factors, including climate, slope inclination and exposition, other topographic and soil features, and controlling floristic composition in the area (Abella and Convington 2006; Sang 2009; Wang et al. 2002; Naqinezhad et al. 2008).
Habitat loss, clearing of natural vegetation for cultivation, overgrazing, and overharvesting are the main threats in the study areas. Space for threatened plants will become even more restricted because of human impacts on the natural process and the limited geographical distribution of the species (Jalili and Jamzad 1999). Among all plants listed as threatened species in the paper (Table 4), some have been under massive economic uses and thus considered within Red data list of Iran. Of which, three endangered species are harvested for economic gain (e.g. Buxus hyrcana and Taxus baccata) and ornamental purposes (Danae racemosa). Buxus hyrcana is a shrub distributed once in majority parts of the lowland Hyrcanian area. There are only some small fragmented patches of this endemic shrub in the area. Furthermore, Taxus baccata is becoming rare and mainly constricted in very steepy and non-reachable places in the forest. The importance of these species as sources of information is crucial to guide conservation efforts (Rodrigues et al. 2006). Conservation policies and ecological management schemes should be applied in order to protect the remaining important, vital but ever so fragile ecosystems of the Hyrcanian area.
References
Abella SR, Convington WW (2006) Vegetation–environment relationships and ecological species groups of an Arizona Pinus ponderosa landscape, USA. Plant Ecol 185:255–268
Akbarinia M, Zare H, Hoseini SM, Ejtehadi H (2004) Study on vegetation structure, floristic composition and chorology of silver birch communities at Sangdeh, forest of Hyrcanian region. Pajouhesh Sazandegi 64:84–96
Akhani H (1998) Plant biodiversity of Golestan National Park, Iran. Stapfia 53:1–411
Akhani H, Djamali M, Ghorbanalizadeh A, Ramezani E (2010) Plant biodiversity of Hyrcanian relict forests, N Iran: an overview of the flora, vegetation, palaeoecology and conservation. Pak J Bot 42:231–258
Archibold OW (1995) Ecology of world vegetation. Chapman & Hall. Press, London, p 510
Assadi M, Massoumi AA, Khatamsaz M, Mozaffrian V (eds) (1988–2011) Flora of Iran. Research Institute of Forests and Rangelands Publication, Tehran, vol 1–73 (in Persian)
Axen GJ, Lam PS, Grove M, Stockli DF, Hassanzadeh J (2001) Exhumation of the west-central Alborz Mountains, Iran, Caspian subsidence, and collision-related tectonics. Geology 29:559–562
Bobek H (1951) Die naturlichen Walder und Geholzfluren Iran. Bonner Geogr Abh 8:1–62
Box EU (1981) Macroclimate and plant forms: an introduction to predictive modeling in phytogeography. Dr.W. Junk Press, The Hague, p 258
Davidson J, Hassanzadeh J, Berzins R, Stockli DF, Bashukooh B, Turrin B, Pandamouz A (2004) The geology of Damavand volcano, Alborz Mountains, northern Iran. GSA Bull 116:16–29
Davis PH (1965–1988) Flora of Turkey and East Aegean Islands. Edinburgh University Press, Edinburgh, p 1–10
Djamali M, de Beaulieu JL, Campagne P, Andrieu-Ponel V, Ponel P, Leroy SAG, Akhani H (2009) Modern pollen rain–vegetation relationships along a forest–steppe transect in the Golestan National Park, NE Iran. Rev Palaeobot Palynol 153:272–281
Djamali M, Akhani H, Khoshravesh R, Andrieu-Ponel V, Ponel P, Brewer S (2011) Application of the global bioclimatic classification to Iran: implications for understanding the modern vegetation and biogeography. Ecologia Mediterranea 37:91–114
Frey W, Probst W (1986) A synopsis of the vegetation of Iran. In: Kürschner H (ed) Contributions to the vegetation of Southwest Asia. Dr. Ludwig Reichert, Wiesbaden, pp 1–43
Frey W, Frahm JP, Fischer E (2006) The liverworts, mosses and ferns of Europe. T. L. Blockeel Press, WolframLobin, p 512
Ghahreman A, Naqinezhad AR, Hamzeh’ee B, Attar F, Assadi M (2006) The flora of threatened black alder forests in the Caspian lowlands, northern Iran. Rostaniha 7(1):5–29
Grytnes JA (2003) Species-richnes spatterns of vascular plants along seven altitudinal transects in Norway. Ecography 26:291–300
Grytnes JA, Vetaas OR (2002) Species richness and altitude: a comparison between simulation models and interpolated plant species richness along the Himalayan altitude gradient, Nepal. Am Nat 159:294–304
Hamzeh’ee B, Naqinezhad A, Attar F, Ghahreman A, Assadi M, Prieditis N (2008) Phytosociological survey of remnant Alnus glutinosa ssp. barbata communities in the lowland Caspian forests of northern Iran. Pytocoenologia 38:117–132
IUCN. 2001. IUCN Red List Categories and Criteria: Version 3.1. Prepared by the IUCN Species Survival Commission. IUCN, Gland, Switzerland, pp ii+ 30
Jafari SM, Akhani H (2008) Plants of Jahan Nama protected area, Golestan province. N Iran Pak J Bot 40:1533–1554
Jalili A, Jamzad Z (1999) Red data book of Iran: a preliminary survey of endemic, rare & endangerd plant species in Iran. Research Institute of Forests & Rangelands Publication, Tehran, p 758
Kent M, Coker P (2002) Vegetation description and analysis: a practical approach. Wiley, New York, p 363
Kessler M (2000) Elevational gradients in species richness and endemism of selected plant groups in the central Bolivian Andes. Plant Ecol 149:181–193
Khalili A (1973) Precipitation patterns of Central Elburz. Theor Appl Climatol 21:215–232
Khoshravesh R, Akhani H, Eskandari M, Greuter W (2009) Ferns and fern allies of Iran. Rostaniha 10(1):1–132
Klein JC (1991) Endemisme á l’ étage alpin de l’ Alborz (Iran). Flora Vegetat Mundi 9:247–261
Klein JC (2001) La végétation altitudinale de L’Alborz Central (Iran): entre les régions Irano-Touranienne et Euro-sibérienne. Institut Français de Recherche, Téhéran
Klimeš L (2003) Life-forms and clonality of vascular plants along an altitudinal gradient in E Ladakh, NW Himalayas. Basic Appl Ecol 4:317–328
Léonard J (1988) Contribution à I’etude la flore et de la végétation des desert d’Iran, Fascicule 8: Etude des aries de distribution, Les phytochories, Les chorotypes. Bulletin of the Jardin Botanique Nacional de Belgique, Meise, p 190
Léonard J (1989) Contribution a l’etude de la flore et de la vegetation des deserts d’Iran, vol 9. Jardin Botanique National de Belgique, Meise
McCain CM, Grytnes JA (2010) Elevational gradients in species richness Encyclopedia of Life Sciences (ELS). John Wiley & Sons Press, Chichester, pp 1–10
Mobayen S, Tregubov V. 1970. Carte de la vegetation naturelle de ľIran, 1:2,500,000. University of Tehran , UNDP/FAO no. IRA 7
Montana C, Valientebanuet A (1998) Floristic and life-form diversity along an altitudinal gradient in an intertropical semiarid Mexicanregion. Southwest Nat 43:25–39
Naqinezhad A, Hamzeh’ee B, Attar F (2008) Vegetation–environment relationships in the alderwood communities of Caspian lowlands, N Iran (toward an ecological classification). Flora 203:567–577
Naqinezhad A, Hosseini S, Rajamand MA, Saeidi Mehrvarz S (2010) A floristic study on Mazibon and Sibon protected forests, Ramsar, across the altitudinal gradient (300–2300 m). Taxon Biosyst 5:93–114
Naqinezhad A, Zare-Maivan H, Gholizadeh H, Hodgson JG (2013) Understory vegetation as an indicator of soil characteristics in the Hyrcanian area, N Iran. Flora 208:3–12
Neilson RP (1993) Transient ecotone response to climatic change; some conceptual and modelling approaches. Ecol Appl 3:385–395
Nicholes GE (1930) Methods in floristic study of vegetation. Ecology 11:127–135
Noroozi J, Akhani H, Breckle SW (2008) Biodiversity and phytogeography of the alpine flora of Iran. Biodivers Coserv 17:493–521
Pavón NP, Hernandez-Trejo H, Rico-Gray V (2000) Distribution of plant life forms along an altitudinal gradient in the semi-arid valley of Zapotitlan, Mexico. J Veg Sci 11:39–42
Rahbek C (1995) The elevational gradient of species richness: a uniform pattern? Ecography 18:200–205
Raunkiaer C (1934) The Life Form of Plants and Statistical Plant Geography. Clarendon Press, Oxford, p 632
Rechinger KH (eds) (1963–2010) Flora Iranica. Akademische Druck-U. Verlagsanstalt, Graz vol 1–178
Rodrigues ASL, Pilgrim JD, Lamoreux JF, Hoffmann M, Brooks TM (2006) The value of the IUCN Red List for conservation. Trends Ecol Evol 21(2):71–76
Sang W (2009) Plant diversity patterns and their relationships with soil and climatic factors along an altitudinal gradient in the middle Tianshan Mountain area, Xinjiang. China. Ecological Research 24(2):303–314
Siadati S, Moradi H, Attar F, Etemad V, Hamzeh’ee B, Naqinezhad A (2010) Botanical diversity of Hyrcanian forests; a case study of a transect in the Kheyrud protected lowland mountain forests in northern Iran. Phytotaxa 7:1–18
Stöcklin J (1974) Northern Iran: Alborz Mountains. Geol Soc London Spec Pub 4:213–234
Takhtajan A (1986) Floristic regions of the World. University of California Press, Berkeley, p 522
Wang G, Zhou G, Yang L, Li Z (2002) Distribution, species diversity and life-form spectra of plant communities along an altitudinal gradient in the northern slopes of Qilianshan Mountains, Gansu, China. Plant Ecol 165:169–181
Zohary M (1973) Geobotanical foundations of the Middle East, vol 2. Fischer, Stuttgart, p 739
Acknowledgments
The authors would like to thank the Department of the Environment of Iran (DOE), the Mazandaran Provincial DOE and Rudsar Department of Natural Resources for their logistics support and assistance during field studies. The authors are also indebted to the University of Mazandaran, for financial support of the project.
Author information
Authors and Affiliations
Corresponding author
Additional information
Corresponding editor: Hu Yanbo
Appendix 1
Appendix 1
See Appendix Table 4
Rights and permissions
About this article

Cite this article
Naqinezhad, A., Zare-Maivan, H. & Gholizadeh, H. A floristic survey of the Hyrcanian forests in Northern Iran, using two lowland-mountain transects. J. For. Res. 26, 187–199 (2015). https://doi.org/10.1007/s11676-015-0019-y
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s11676-015-0019-y