
Abstract
Physical exercise and the training effects of repeated practice of skills over an extended period of time may have additive effects on brain networks and functions. Various motor skills and attentional styles can be developed by athletes engaged in different sports. In this study, the effects of fast ball sports and dance training on attention were investigated by event related potentials (ERP). ERP were recorded in auditory and visual tasks in professional dancer, professional fast ball sports athlete (FBSA) and healthy control volunteer groups consisting of twelve subjects each. In the auditory task both dancer and FBSA groups have faster N200 (N2) and P300 (P3) latencies than the controls. In the visual task FBSA have faster latencies of P3 than the dancers and controls. They also have higher P100 (P1) amplitudes to non-target stimuli than the dancers and controls. On the other hand, dancers have faster latencies of P1 and higher N100 (N1) amplitude to non-target stimuli and they also have higher P3 amplitudes than the FBSA and controls. Overall exercise has positive effects on cognitive processing speed as reflected on the faster auditory N2 and P3 latencies. However, FBSA and dancers differed on attentional styles in the visual task. Dancers displayed predominantly endogenous/top down features reflected by increased N1 and P3 amplitudes, decreased P1 amplitude and shorter P1 latency. On the other hand, FBSA showed predominantly exogenous/bottom up processes revealed by increased P1 amplitude. The controls were in between the two groups.
Similar content being viewed by others


Explore related subjects
Discover the latest articles, news and stories from top researchers in related subjects.Avoid common mistakes on your manuscript.
Introduction
Performance on a variety of sports, for instance, skiing, dancing, and fast ball sports like basketball, improves through training. Long term physical exercise and repeated practice of skills also can induce positive changes on brain functions through plasticity (Nakata et al. 2010). Visual attention is an important component of performance in motor skills in sports and attentional styles are developed depending on the type exercise being executed by athletes (Fontani et al. 1999). During a competition or performance of dance, overwhelming amount of information is being encountered at least by visual system. One has to select the information that is relevant to the task and to process them. As processing of information has a limited capacity, in order to select the appropriate information for the relevant event, suitable mechanisms are needed to guide attention. One mechanism that involves attentional selection is called endogenous, goal directed or top/down processes and the other is exogenous, involuntary, stimulus-driven or bottom/up attention (Reynolds and Heeger 2009; Stoppel et al. 2013). For the sake of simplicity, endogenous attention occurs when we have prior representation for the relevant location and in the case of exogenous attention salient stimuli capture attention (Corbetta and Shulman 2002; Fox et al. 2006; Yantis and Jonides 1984). In this respect dancers and fastball sports athletes (FBSA) might have opposite attentional demands in visual tasks. In fast ball sports, the athlete encounters unexpected information under time pressure and she/he has to cope with those changes predominantly by involuntary, stimulus driven attentional shifts (Runigo et al. 2010). FBSA encounter continuously new information, they might rely heavily on working memory based processing. FBSA also have to extract auditory information among the noise of cheers, acclamations and shouts of team mates and opponents. On the other hand, in dancing, representations of figures are practiced repeatedly and accord with the involved music. Perfection, harmony, alignment and consistency are aimed however, unexpected events are avoided (Minvielle-Moncla et al. 2008). Dancers seem to rely mostly on endogenously oriented visual attention.
However, it is challenging to examine the cognitive functions of athletes during competition or performance due to difficulties regarding movements. One way to examine the brain of athletes is to record EEG before and after performance and assess short term cognitive functions and comparisons are made between professionals and novices for the long-term effects (Basar 2013; Thompson et al. 2008).
P3 component of ERP is derived from EEG epochs time locked to visual or auditory stimuli or a cognitive event related to the stimuli (Picton and Hillyard 1988). P3 is usually elicited in the oddball paradigm where subjects give motor response to or count target stimuli that occur irregularly and infrequently among frequent “standard” or non-target stimuli. It is a positive going wave elicited around 300 ms after target stimulus presentation (Ritter and Vaughan 1969; Linden et al. 1999). P3 was thought to result from cognitive activities such as context updating, allocation of attention and working memory. The amplitude of P3 is supposed to reflect allocation of attention to a given task (Schubert et al. 1998; Wickens et al. 1983), whereas the latency is considered to be a measure of stimulus evaluation time (Kutas et al. 1977). In ERP studies examining visual spatial attention, the effects of endogenous attention were found about 300 ms after stimuli coinciding with P3 latency. On the other hand, the involuntary deployment of attention is faster and peaks at about 100–120 ms around the time window of P100 (P1) component (Liu et al. 2007; Remington et al. 1992).
P3 latencies were found to be related to superior cognitive performance in normal subjects (Emmerson et al. 1989; Polich and Martin 1992) and P3 latencies increased progressively as a result of dementing illnesses and aging (Brown et al. 1982; Polich et al. 1986). Lavoie et al. (2004) also showed P3 amplitude differences among college athletes who had concussion with symptoms, without symptoms and athletes that never had concussion which highlights the sensitivity of P3 amplitude over symptom presentation and neuropsychological tests. Therefore measuring latency and amplitude of ERP components could be useful not only to compare exercise groups and controls but also to make comparisons among different exercise groups.
Aside from repeatedly reported differences in information processing between exercise and non-exercise groups (Rossi et al. 1992; Iwadate et al. 2005), Brümmer et al. (2011) reported also that “cortical activation patterns depend on exercise mode and intensity and individual exercise preferences”. Recently Ermutlu et al. (2015) reported significant quantitative EEG (qEEG) differences between dancers and fast ball players. Bianco et al. (2017) compared fencers and football players with controls. Overall athletes were faster than controls and they had better motor preparation revealed by Bereitshaftspotential. Fencers also were more accurate in their responses and better top down control reflected by increased prefrontal negativity.
We chose to use oddball paradigm with visual spatial stimuli so that to be attended stimuli around 300 ms would reflect endogenous attention and not to be attended stimuli around 100–200 ms time window would give information about exogenous attention. In addition, ERP to auditory stimuli were also recorded in order to obtain electrophysiological evidence of cognitive functions like processing speed, allocation of attention, context updating and working memory and replicate or compare them with studies that used oddball paradigm.
In this study, we hypothesize that overall as a general effect of training FBSA and dancers may show faster latencies and or higher P3 amplitudes than the controls. However, in the visual task FBSA and dancers may be differentiated in respect to P1 to unattended stimuli and P3 amplitudes to attended stimuli where dancers may have increased P3 amplitudes reflecting predominance of endogenous orientation of attention and FBSA may have increased P1 amplitudes related to exogenous orientation of attention.
Materials and methods
Participants
Professional dancers, professional FBSA, and healthy controls twelve in each group participated to the study. Female/male ratio was equal in each group. All participants were right handed. Average duration of physical activity was in the range of 7–13 years. Dancers were practicing/exercising 5–6 days a week for more than 2–3 h per day, and FBSA (6 basketball and 6 volleyball players) were practicing/exercising 5–6 days a week for 2–4 h a day. Controls were healthy sedentary age- and sex-matched volunteers (see Table 1). All participants were highly motivated. The participants had no neurological, psychiatric disorders or sensory deficiencies. All participants provided written informed consent before participation to the study. The study was performed in accordance with the Declaration of Helsinki. Participants were free to withdraw from the study any time. The study was approved by the ethical committee of Istanbul University, Istanbul Faculty of Medicine, approval no: 2010/450-123.
Experimental procedure
The EEGs were recorded in a sound- and echo-free, dimly illuminated room. All participants were informed about the recording session and the duration of the session. Participants were all drug-free and had abstained from caffeine on the day of measurement. For the EEG recordings, all participants were asked to have slept well and be well rested the night before. They were tested after abstaining from food for 2–3 h. All participants were explained about target and non-target (standard) stimuli during the ERP recording session. They were instructed to maintain fixation, and to minimize blinking and eye movements. The computer screen was set 100 cm far from the subjects. The EEG were recorded unipolarly from 19 scalp electrode sites of the international 10–20 system (Fp1, Fp2, F7, F3, Fz, F4, F8, T3, C3, Cz, C4, T4, T5, P3, Pz, P4, T6, O1, and O2) with an electro-cap system referenced to the earlobes. Grounding electrodes were also located at both earlobes. EEG was recorded using a 19-channel, digital EEG amplifier (Mitsar-EEG-201: Mitsar Co. Ltd., St. Petersburg, Russia Federation; bandwidth 0.3–30 Hz, sampling frequency 500 Hz) and the WinEEG v.2.4 Software recording and analysis package (Mitsar Co. Ltd., St. Petersburg, Russia Federation). Eye blinking and eye movements were monitored by bipolar electrooculographic (EOG). Electrode impedance was kept under 5 kΩ using Quick Gel®. The raw data were digitized at a sampling rate of 500 Hz with an online bandpass filter of 0.3–30 Hz in continuous recording mode. Electrical activity of the brain on the scalp recorded in two different sessions: visual P3 and auditory P3. After automatic artifact rejection, the artifact free epochs were again visually inspected for eye movement or blink artifact. Stimuli were created and presented using the Psytask v.1.52 Software (Mitsar Co. Ltd., St. Petersburg, Russia Federation).
Visual oddball paradigm
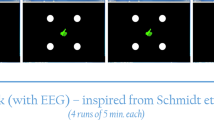
Psytask programme was used on a screen for Visual Oddball presentation. The visual stimuli were discriminated according to their spatial location. Target stimuli were a blue square in the bottom whereas standard stimuli were a blue square at the top of the background (Fig. 1).

Visual oddball Paradigm. The stimulus was presented binocularly on a computer screen. The stimuli consisted of target and standard stimuli presented with probabilities of 0.20 and 0.80, respectively. The volunteers were instructed to discriminate the target stimuli from the standard stimuli and to press a button of the computer mouse immediately following target stimulus. Target stimuli was a blue square in the bottom of the background and standard stimuli was the same square presented at the top of the background
Auditory oddball paradigm
Auditory Oddball was presented binaurally with a headphone by Psytask stimulus program. Target stimuli were the high frequency tone (1300 Hz) and standard stimuli was the low frequency tone (1000 Hz) and participants focused a fixation point on the screen.
In both visual spatial and auditory oddball tasks 330 stimuli were presented randomly. The inter-stimulus interval (ISI) was 2 s. The probabilities of the targets, and standard stimuli were 0.20 and 0.80, respectively. The volunteers were asked to discriminate targets from the standard stimuli and to press the button of the mouse after the target stimuli.
Analysis of the ERPs responses
EEG epochs were consisted of 1200, 200 ms pre-stimulus and 1000 ms after-stimulus. Artifact free EEG epochs were analyzed. The amplitudes and latencies of the averaged non-target visual P1 and N1, auditory N2 and visual and auditory target P3 components were measured. The visual P1 was defined as the largest positive wave between 70 and 130 ms after stimulus onset, visual N1 was defined as the largest negative wave between 120 and 190 ms after stimulus onset, the auditory N2 was defined the largest negative wave between 200 and 350 ms after stimulus onset and the visual P300 as the largest positive wave between 250 and 700 ms after stimulus onset. The peak amplitude was defined relative to baseline and the peak latency was defined as the time from stimulus onset to the maximum peak.
Statistical analysis
The latency and amplitude of the ERP components (non-target P1 and N1 and target P3) of the exercise groups and the volunteers were compared by ANOVA. For post hoc analysis Bonferroni tests were used.
Statistical analyses were carried out with IBM-SPSS version 20.0, with alpha set at 0.05.
Results
Demographic data
The ages of dancers were significantly higher than the other groups [F(2,33) = 21.33; p = 0.001] (see Table 1).
Electrophysiological data
Comparison of P3 responses of visual oddball paradigm
The P3 amplitudes of visual target stimuli showed statistical differences between the groups [F(2,627) = 29.46; p = 0.0001] (Fig. 2). In the post hoc Bonferroni test, amplitude of target P3 of the dancers (9.47 µV) were significantly higher than amplitudes of FBSA (6.67 µV) and amplitudes of controls (6.96 µV) p = 0.0001, p = 0.0001; (respectively). The P3 latencies of visual target stimuli were statistically different between the groups [F(2,627) = 4.674; p = 0.01]. The latency of target P3 of the FBSA (373.2 ms) were significantly earlier than controls (383.2 ms) with post hoc Bonferroni testing (respectively; p = 0.008).

Grand averages ERPs to visual spatial target stimuli of dancers (dotted line), FBSA (broken line) and controls (solid line)
Comparison of P3 responses of auditory oddball paradigm
The P3 amplitudes of auditory target stimuli were not statistically significant between the groups [F(2,627) = 1.216; p = 0.297] (Fig. 3). However the P3 latencies of auditory target stimuli showed statistical differences between the groups [F(2,627) = 5.642; p = 0.004]. The latency of target P3 of the FBSA (377.2 ms) and dancers (378.8 ms) were significantly earlier than latency of the controls (385.5 ms) with post hoc Bonferroni testing (respectively; p = 0.012; p = 0.002).

Grand averages ERPs to auditory target stimuli of dancers (dotted line), FBSA (broken line) and controls (solid line)
Comparison of N2 responses of auditory oddball paradigm
The N2 amplitudes of auditory target stimuli were not statistically significant between the groups [F(2,627) = 2.243; p = 0.107] (Fig. 3). However the N2 latencies of auditory target stimuli showed statistical differences between the groups [F(2,627) = 71.262; p = 0.000]. The latency of target N2 of the dancers (240.1 ms) and the FBSA (249 ms) were significantly earlier than controls (260 ms).
Comparison of non-target P1 responses of visual oddball paradigm
The P1 amplitudes of visual non-target stimuli were statistically different between the groups [F(2,627) = 37.243; p = 0.0001] (Fig. 4). The amplitude of non-target P1 of both the FBSA (2.71 µV) and the controls (2.35 µV) were significantly higher than amplitudes of the dancers in the post hoc Bonferroni test (1.25 µV) (respectively; p = 0.0001, p = 0.0001). The latencies of P1 of visual non-target stimuli were statistically significant between the groups [F(2,627) = 1867.177; p = 0.0001]. In the post hoc Bonferroni test the latency of non-target P1 of the dancers (70.8 ms) were significantly earlier than FBSA (123.9 ms) and latency of the controls (129 ms) (respectively; p = 0.0001, p = 0.0001). Also the latency of non-target P1 of the FBSA (123.9 ms) were significantly earlier than latency of the controls in the post hoc Bonferroni test (129 ms) (p = 0.0001).

Grand averages ERPs to visual spatial non-target stimuli of dancers (dotted line), FBSA (broken line) and controls (solid line)
Comparison of non-target N1 responses of visual oddball paradigm
The N1 amplitudes of non-target stimuli showed statistical differences between the groups [F(2,627) = 30.722; p = 0.0001] (Fig. 4). The amplitude of non-target N1 of the dancers (− 3.71 µV) were significantly higher than the amplitudes of both FBSA (− 2.50 µV) and controls (− 2.21 µV) with post hoc Bonferroni testing (respectively; p = 0.0001, p = 0.0001). The grand average N1 latencies of visual non-target stimuli showed statistical differences between the groups [F(2,627) = 765.513; p = 0.0001]. The latency of non-target N1 of the dancers (126.1 ms) were significantly earlier than latencies of both FBSA (179.6) and controls (182.8 ms) with post hoc Bonferroni testing (respectively; p = 0.0001, p = 0.0001).
Discussion
In this study, we found significant amplitude and latency differences of ERP between the controls and exercise groups and among the exercise groups. Overall, exercise groups have shorter N2 and P3 latencies than controls in the auditory task however the exercise groups were differentiated in the visual task in respect to P3 amplitude to targets and P1, N1 amplitudes and latencies to non-target stimuli.
In the visual task the while dancers had shorter latencies to early components of non-target stimuli, namely P1 and N1, FBSA had shorter P3 latency. In the case of ERP amplitudes, FBSA had larger P1 amplitudes to non-target stimuli whereas dancers had larger amplitude to non-target N1 and target P3 than FBSA and controls.
In an electrophysiological study Bashore (1989) reported that low-fit subjects have smaller P3 amplitude and longer P3 than the high-fit groups. Dustman et al. (1990) also compared low-fit to high-fit subjects using a visual oddball task and confirmed that high fit subjects had shorter latencies as well. Furthermore, the age effect of significant longer P3 latency was attributable to the low-fit older men. In volleyball players. Fontani et al. (1999) reported larger P3 amplitude and shorter latency compared with controls. Iwadate et al. (2005) found lower limb somatosensory P3 amplitude was larger and latency shorter in football players. Taddei et al. (2013) compared young and middle aged fencers and non-athletes. In fencers, the latencies P3 component were found to be longer in older subjects, however fit older participants displayed less delay. The fencers had shorter N2 latencies and higher amplitudes of the same component than non-athletes. In no-go trials, the amplitude of P3 component was larger in all fencer groups irrespective of age.
Aforementioned studies displayed increased P3 amplitude and/or shortened latency in exercise groups compared with controls. Overall results confirmed that exercise have positive influences on cognitive functions reflected in P3 ERP component (Hillman et al. 2008).
Auditory processing is a very important component of daily life however it is especially vital in competitive fast ball sports and in performing arts like dance. During competitive sports the athlete has to extract auditory signal such as command of the coach and communication of team friends from the noise of the spectators and the opponent team and organize timing of his movements. In contrast to FBSA, dancers do not perform in a noisy setting. Instead, the accompanying music helps dancer to synchronize her/his movements according to another dancer while acting the choreography. Music is an important part of timing in the practice of dance as musical cues help to act the harmonization of movements at specific time points (Bläsing et al. 2009).
Even though nature of auditory environments are completely different, intensive auditory processing and timing according to auditory stimuli are part of the two exercise groups. Intense exercise may activate the networks responsible from auditory related cognition and might have had plastic changes that resulted in faster processing.
Unlike the auditory domain, visual attention involves scanning of the visual field. Often, we direct our visual attention with eye movements, but it also is possible to direct one’s attention covertly around the visual field while keeping the eyes fixated on a particular point. The covert attention helps to monitor the environment and can facilitate to simultaneously deploy eye movements and attention more than one location (Carrasco 2011).
It was proposed that the covert attention system has two components that facilitate processing and selecting information. Information can be selected endogenously (top down) or exogenously (bottom-up), depending on the context and the task at hand (Carrasco 2011).
The processing according to internal goals and tasks were carried out by endogenous orienting in dorsal fronto-parietal network (Chica et al. 2013). However, ventral fronto-parietal network was driven by unexpected but behaviorally relevant events through bottom up processes via exogenous mechanisms.
The dorsal fronto-parietal network comprises the Intraparietal Sulcus (IPS) and Superior Parietal Lobule (SPL) of dorsal parietal cortex, and the dorsal frontal cortex along the precentral sulcus, near or at the frontal eye fields (FEF) (Bundesen 1990; Desimone and Duncan 1995).
When a subject was instructed to see an object at a particular location or with certain features, and asked to respond to a specific stimulus, the dorsal network is activated (Corbetta et al. 2008). The preparatory activation of the dorsal frontoparietal network could affect the visual cortex with top-down modulation either by inhibiting or facilitating the sensory traces (Coull and Nobre 1998).
While endogenous attention is deployed about 300 ms the exogenous deployment of attention is at about 100–120 ms (Hein et al. 2006; Ling and Carrasco 2006; Liu et al. 2007; Nakayama and Mackeben 1989; Remington et al. 1992). In an ERP study, it was found that occurrence of exogenous attention coincides with the time window of P1 component however, the effect of endogenous attention was demonstrated at the time window corresponding to N1 and P3 (Busse et al. 2008).
The ERP data seems to fit the peculiarities of the situation in the FBS game. Increased P1 amplitude to non-targets might reflect predominant orientation of exogenous attention and bottom up selection processing whereas shorter P3 latencies might indicate faster stimulus categorization speed in response to unpredictable and competitive settings. However, in the case of dancers, they perform and practice according to internal goals and expectations based on choreography. They expect to see a partner’s movement at a particular location and prepare a specific response accordingly which seems similar to tasks that elicit endogenous attention. Increased P3 amplitude in the visual task suggests dancers have stronger control of endogenous orientation of attention than FBSA and controls. On the other hand, decreased P1 amplitude and shorter P1 and N1 latencies to non-target stimuli might suggest gating out the bottom up sensory stimuli which may interfere with endogenous task.
In this study, we showed that dancers and FBSA have different visual attentional styles which might be innate and/or be induced by plastic changes that could have occurred as a consequence of years of training. High brain-derived neurotrophic factor (BDNF) induced by intermittent high intensity training might have brought about plastic changes on brain networks that were active during the performance of aforementioned exercises (Marquez et al. 2015).
Cross sectional nature of the study and recording of EEG outside performance due to technical difficulties limits our conclusions. Longitudinal studies should be carried out in order to reveal whether this finding is the effect of long term, intense training or call of nature in accord with talent for a particular exercise Also if technical possibilities permit measurement should be repeated during real conditions.
Exercise and dance is also used in the therapy of some medical conditions. Exercise training seems to have broad effects on cognition as revealed by task related to speed, spatial processing, controlled processing and executive control tasks (Belardinelli et al. 2008; Kramer and Erickson 2007). However as shown in our study, different kinds of exercises not only have general exercise dependent effects but also have specific impacts on cognitive components. Those specific futures might be used to choose the right athlete to appropriate exercise type. Also future studies are needed to apply the differential effects of exercise on different kinds of cognitive disorders.
References
Basar E (2013) Brain oscillations and neuropsychiatry. Dialogues Clin Neurosci 15:291–300
Bashore TR (1989) Age, physical fitness, and mental processing speed. Ann Rev Gerontol Geriatr 9:120–144
Belardinelli R, Lacalaprice F, Ventrella C, Volpe L, Faccenda E (2008) Waltz dancing in patients with chronic heart failure new form of exercise training. Circ Heart Fail 1:107–114
Bianco V, Di Russo F, Perri RL, Berchicci M (2017) Different proactive and reactive action control in fencers’ and boxers’ brain. Neuroscience 343:260–268
Bläsing B, Tenenbaum G, Schack T (2009) The cognitive structure of movements in classical dance. Psychol Sport Exerc 10:350–360
Brown WS, Marsh JT, LaRue A (1982) Event-related potentials in psychiatry: differentiating depression and dementia in the elderly. Bull Los Angel Neurol Soc 47:91–107
Brümmer V, Schneider S, Abel T, Vogt T, Strüder HK (2011) Brain cortical activity is influenced by exercise mode and intensity. Med Sci Sports Exerc 43(10):1863–1872
Bundesen C (1990) A theory of visual attention. Psychol Rev 97(4):523–547
Busse L, Katzner S, Tillmann C, Treue S (2008) Effects of attention on perceptual direction tuning curves in the human visual system. J Vis 8(9):1–13
Carrasco M (2011) Visual attention: the past 25 years. Vis Res 51(13):1484–1525
Chica AB, Bartolomeo P, Lupiánez J (2013) Two cognitive and neural systems for endogenous and exogenous spatial attention. Behav Brain Res 237:107–123
Corbetta M, Shulman GL (2002) Control of goal-directed and stimulus driven attention in the brain. Nat Rev Neurosci 3:201–215
Corbetta M, Patel G, Shulman GL (2008) The reorienting system of the human brain: from environment to theory of mind. Neuron 58(3):306–324
Coull JT, Nobre AC (1998) Where and when to pay attention: the neural systems for directing attention to spatial locations and to time intervals as revealed by both PET and fMRI. J Neurosci 18(18):7426–7435
Desimone R, Duncan J (1995) Neural mechanisms of selective visual attention. Ann Rev Neurosci 18:193–222
Dustman RE, Emmerson RY, Ruhling RO, Shearer DE, Steinhaus LA, Johnson SC, Bonekat HW, Shigeoka JW (1990) Age and fitness effects on EEG, ERPs, visual sensitivity, and cognition. Neurobiol Aging 11:193–200
Emmerson RY, Dustman RE, Shearer DE, Turner C (1989) P3 latency and symbol digit performance correlations in aging. Exp Aging Res 15:15l–159
Ermutlu N, Yucesir İ, Eskikurt G, Temel T, İşoğlu-Alkaç U (2015) Brain electrical activities of dancers and fast ball sports athletes are different. Cogn Neurodyn 9(2):257–263
Fontani G, Maffei D, Cameli S, Polidori F (1999) Reactivity and event-related potentials during attentional tests in athletes. Eur J Appl Physiol 80:308–317
Fox MD, Corbetta M, Snyder AZ, Vincent JL, Raichle ME (2006) Spontaneous neuronal activity distinguishes human dorsal and ventral attention systems. Proc Natl Acad Sci USA 103:1046–1051
Hein E, Rolke B, Ulrich R (2006) Visual attention and temporal discrimination: differential effects of automatic and voluntary cueing. Vis Cogn 13(1):29–50
Hillman CH, Erickson K, Kramer AF (2008) Be smart, exercise your heart: exercise effects on brain and cognition. Nat Rev Neurosci 9:58–65
Iwadate M, Mori A, Ashizuka T, Takayose M, Ozawa T (2005) Long-term physical exercise and somatosensory event-related potentials. Exp Brain Res 160:528–532
Kramer AF, Erickson KI (2007) Effects of physical activity on cognition, well-being, and brain: human interventions. Alzheimer Dement 3:45–50
Kutas M, McCarthy G, Donchin E (1977) Augmenting mental chronometry: the P300 as a measure of stimulus evaluation time. Science 197:792–795
Lavoie ME, Dupuis F, Johnston KM, Leclerc S, Lassonde M (2004) Visual P300 effects beyond symptoms in concussed college athletes. J Clin Exp Neuropsychol 26(1):55–73
Linden DEJ, Prvulovic D, Formisano E, Vollinger M, Zanella FE, Goebel R, Dierks T (1999) The functional neuroanatomy of target detection: an fRMI study of visual and auditory oddball tasks. CerebralCortex 9:815–823
Ling S, Carrasco M (2006) Sustained and transient covert attention enhance the signal via different contrast response functions. Vis Res 46(8–9):1210–1220
Liu T, Stevens ST, Carrasco M (2007) Comparing the time course and efficacy of spatial and feature-based attention. Vis Res 47(1):108–113
Marquez CMS, Vanaudenaerde B, Troosters BT, Wenderoth N (2015) High-intensity interval training evokes larger serum BDNF levels compared with intense continuous exercise. J Appl Physiol 119:1363–1373
Minvielle-Moncla J, Audiffren M, Macar F, Vallet C (2008) Overproduction timing errors in expert dancers. J Mot Behav 40:291–300
Nakata H, Yoshie M, Miura A, Kudo K (2010) Characteristics of the athletes’ brain: evidence from neurophysiology and neuroimaging. Brain Res Rev 62:197–211
Nakayama K, Mackeben M (1989) Sustained and transient components of focal visual attention. Vis Res 29(11):1631–1647
Picton TW, Hillyard SA (1988) Endogenous event-related potentials. In: Picton TW (ed) Human event-related potentials. Elsevier, New York, pp 361–426
Polich J, Martin S (1992) P300, cognitive capability and personality: a correlational study of university undergraduates. Personal Individ Differ 13:533–543
Polich J, Ehlers CL, Otis S, Mandell A, Bloom FE (1986) P300 latency reflects the degree of cognitive decline in dementing illness. Electroencephalogr Clin Neurophysiol 63:138–144
Remington RW, Johnston JC, Yantis S (1992) Involuntary attentional capture by abrupt onsets. Percept Psychophys 51(3):279–290
Reynolds JH, Heeger DJ (2009) The normalization model of attention. Neuron 61:168–185
Ritter W, Vaughan HG (1969) Averaged evoked responses in vigilance and discrimination: a reassessment. Science 164:326–328
Rossi B, Zani A, Taddei F, Pesce C (1992) Chronometric aspects of information processing in high level fencers as compared to non-athletes: an ERPs and RT study. J Hum Mov Stud 23:17–28
Runigo CL, Benguigui N, Bardy BG (2010) Visuo-motor delay, information movement coupling, and expertise in ball sports. J Sports Sci 28:327–337
Schubert M, Johannes S, Koch M (1998) Differential effects of two motor tasks on ERPs in an auditory classification task: evidence of shared cognitive resources. Neurosci Res 30:125–134
Stoppel CM, Boehler CN, Strumpf H, Krebs RM, Heinze HJ, Hopf JM, Schoenfeld MA (2013) Distinct representations of attentional control during voluntary and stimulus-driven shifts across objects and locations. Cereb Cortex 23:1351–1361
Taddei F, Bultrini A, Spinelli D, Di Russo F (2013) Neural correlates of attentional and executive processing in middle-age fencers. Med Sci Sports Exerc 44(6):1057–1066
Thompson T, Steffert T, Ros T, Leach J, Gruzelier J (2008) EEG applications for sport and performance. Methods 45:279–288
Wickens C, Kramer A, Vanasse L, Donchin E (1983) The performance of concurrent tasks: a psychophysiological analysis of the reciprocity of information processing resources. Science 221:1080–1082
Yantis S, Jonides J (1984) Abrupt visual onsets and selective attention: evidence from visual search. J Exp Psychol Hum Percept Perform 10:601–621
Acknowledgements
The authors would like to thank the athletes, the dancers and their coaches for their cooperation and participation in this study. The results of the present study do not constitute endorsement by the American College of Sports Medicine.
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Conflict of interest
The authors declare that they have no conflict of interest.
Rights and permissions
About this article

Cite this article
Isoglu-Alkac, U., Ermutlu, M.N., Eskikurt, G. et al. Dancers and fastball sports athletes have different spatial visual attention styles. Cogn Neurodyn 12, 201–209 (2018). https://doi.org/10.1007/s11571-017-9469-6
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s11571-017-9469-6