
Abstract
Cytoskeletal proteins determine cell shape and integrity and membrane-bound structures connected to extracellular components allow tissue integrity. These structural elements have an active role in the interaction of blood vessels with their environment. Shear stress due to blood flow is the most important force stimulating the endothelium. The role of cytoskeletal proteins in endothelial responses to flow has been studied in resistance arteries using pharmacological tools and transgenic models. Shear stress activates extracellular “flow sensing” elements associated with a thick glycocalyx communicating the signal to membrane-bound complexes (integrins and/or dystrophin-dystroglycans) and to eNOS through a pathway involving the intermediate filament vimentin, the microtubule network and actin. When blood flow increases chronically the endothelium triggers diameter enlargement and medial hypertrophy. This is facilitated by the genetic absence of the intermediate filaments, vimentin and desmin suggesting that these elements oppose the process.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
1 Introduction
Cell shape and integrity relies in great part on the structure of the cytoskeleton and tissue integrity relies on cell cytoskeleton through membrane-bound structures and extracellular components. These structural elements are no longer regarded as passive architectural components but an increasing number of experiments evidence their active role in most biologically active processes. In blood vessels cytoskeletal components have been widely shown to be involved in smooth muscle cells contractility. Smooth muscle cells actin fibers are the key contractile proteins and their connection to the plasma membrane allows coordination of the force between cells for an optimal contractility. Cytoskeletal proteins have also a key role in cell motility in angiogenesis associated with growth, wound healing and post-ischemic revascularization [1, 2]. In these situations, endothelial cells respond first and their cytoskeleton undergoes dramatic and rapid changes. Shear stress due to blood flow is probably the most important driving force pushing endothelial cells in the direction on new vessels [3]. More globally, the endothelium plays a central role in vascular homeostasis and allows the vasculature to adapt immediately to the local need of each tissue. The role of cytoskeletal proteins in this acute endothelial responsiveness is now better understood, especially when the stimulus is a mechanical factor such as shear stress. Drugs disturbing the cytoskeleton, although not always very selectively, affect preferentially flow (shear stress)-mediated dilation [4]. Similarly, gene knockout of vimentin, an intermediate filament shaping the endothelial cell architecture, is associated with strong reduction in flow-mediated dilation (FMD) [5]. Genetic deficiency in integrins or dystrophin, both anchoring cytoskeletal elements to the plasma membrane, also reduce FMD [6]. When blood flow increases chronically, the endothelium triggers a remodeling of the arterial wall with diameter enlargement and hypertrophy. This phenomenon is facilitated by the genetic absence of the intermediate filaments, vimentin or desmin [7, 8] suggesting that intermediate filaments stabilize arterial cell shape and oppose the remodeling process. Nevertheless, other proteins such as dystrophin are important in the flow sensing process after either short- or long-term simulation [6, 9].
This article will detail the role of structure proteins in flow-mediated dilation (FMD, acute response) and flow-induced remodeling (chronic response) in resistance arteries.
1.1 Mechanotransduction in the microcirculation: acute responses
1.1.1 Control of microvascular tone and role of shear stress
The vasculature consists of large arteries branching out into smaller and smaller vessels terminated by capillaries. Capillaries irrigate tissues and are connected to venules that associate together to form larger veins. Structural and functional differences between large arteries (representing most of the available literature) and arterioles (representing most of the arterial vasculature) are obvious when comparing the aorta to pre-capillary arterioles. Indeed, differences in blood pressure, pulsatility, and organ or tissue specificity induce a specialization of vascular cells. Changes in haemodynamic environment induce short-term responses and structural adaptation of the vascular tree, thus allowing an optimal tissue perfusion [10, 11]. Small arteries having the most important influence on local blood flow are called resistance arteries. They are submitted to chemical and neuro-hormonal influences in addition to the continuous effect of the mechanical factors generated by the blood stream. This latter affects arterial tone through the effect of pressure and flow. Pressure induces a myogenic response, which is a rapid and sustained vasoconstriction [12]. Myogenic tone represents a tonic arteriolar contraction or basal tone facilitating the action of the other vasoactive systems through a synergistic mechanism, or potentiation, controlling the sensitivity to calcium of the contractile apparatus [13–16].
On the other hand, blood flow produces shear stress, which stimulates the endoluminal surface of the endothelium. The discovery that the endothelium is a strong modulator of vascular tone followed by the discovery of nitric oxide (NO) as a vasodilator produced by endothelial cells is one of the greatest advances in modern vascular biology [17, 18]. For the endothelium shear stress due to blood flow represents a basal stimulus activating continuously the endothelium and together with myogenic tone FMD determines a basal tone over which other vasoactive systems are more efficient. The large number of studies, which followed the discovery of NO, was focused on large arteries. Nevertheless when investigating endothelium-dependent dilation and more specifically FMD in smaller arteries it became evident that besides NO a large number of other vasoactive substances are produced by the endothelial cells thus providing the arterioles located in each tissue a very complete arsenal for the control of blood flow. A reduction in FMD is the hallmark of endothelial dysfunction and it is becoming more and more important in the detection, the follow-up and even the prevention of vascular disorders in a growing number of diseases described in recent review articles [19–21].
1.1.2 Mechanotransduction of shear stress: role of the structure proteins
Sensing and transducing shear stress into a biological signal is a process remaining mainly unknown in the peripheral circulation. In addition, shear stress is extremely small, a few dyn per centimeter square (compared to thousands for blood pressure). Shear stress stimulates very gently the intraluminal side of the endothelial cells. Thus the first challenge is to understand the sensing process of this force by the glycocalix and/or by or structures in the extracellular space before its transmission to the intracellular machinery through the plasma membrane. A flow sensor is probably made of the interaction between several elements of the extracellular matrix as described in cultured endothelial cells [22] but with a time scale not necessarily compatible with FMD, which occur in the seconds following a change in blood flow. In FMD a possible “flow sensor” might be made of proteoglycans carrying negative charges and binding extracellular sodium cations as described by Bevan [11]. Indeed, the capacity of proteoglycans to bind sodium is sensitive to shear stress [23, 24] and the response of blood vessels to flow involves preferentially sodium influx [25]. In addition, shear-induced dilation of small arteries is abolished by removal of sialic acids by neuraminidase [26] and in cultured bovine aortic endothelial cells the partial removal of heparn sulfate proteoglycan with heparinase prevents shear-induced NO release [27]. The importance of the glycocalyx is also evidenced in diabetic patients, who have a reduced FMD in association with a lower glycocalyx volume and an enhanced level of circulating glycocalyx elements [28]. As discussed in a recent review article describing the importance of a thick glycocalyx for endothelium integrity [29], numerous factors modulate the integrity of the endothelial glycocalyx. Furthermore, endothelial carbohydrate damage during ischemia reperfusion, hypoxia or in an oxidative environment with high oxidized low-density lipoprotein level, contribute to tissue edema and alteration in shear stress sensing [29].
The ultrastructural analyses of the glycocalyx shows that it forms a network, which parallels a cortical web located beneath the membrane and made of filamentous actin [30]. Nevertheless, this observation does not show how shear stress is transmitted to this actin network, as shear stress is normally equivalent to zero at the level of the membrane. It is likely that equilibrium between sodium influx as described by Bevan [11] and a direct transmission of a deformation to the membrane-anchored complexes and to the cytoskeleton might occur in FMD.
The role of the actin network and of the microtubules in mechanotransduction is now well accepted although the exact pathway involved remains unclear. The use of cytochalasin B to depolymerize and phalloidin to stabilize filamentous actin microfilament and microtubule networks reduced FMD in the rabbit abdominal aorta without affecting acetylcholine-induced dilation [4]. In addition, the transformation of filamentous into glomerulous (inactive) actin might also play a role in FMD as shear stress activates rapidly this transformation [31]. Interestingly, in the response of resistance arteries to pressure (myogenic tone), the opposite effect was observed with a rapid formation of filamentous actin after a rise in pressure [32]. Microtubule disassembly with nocodazole or colchicine also suppressed FMD in isolated rat gracilis arterioles without affecting the dilation due to acetylcholine or sodium nitroprusside [33]. Thus a selective pathway leading to the production of NO is activated by shear stress in endothelial cells, not by acetylcholine or other endothelium-dependent vasodilator agents. This pathway requires a transmission of the signal from the extracellular matrix to the enzyme eNOS. Several protein complexes are bound to the plasma membrane and usually concentrated in caveolin-rich area, which also link eNOS [34].
Two main complexes have been studied, the dystrophin-dystroglycan complex and the integrins complexes. They have been mainly studied in isolated muscular and endothelial cells respectively. The role of integrins in flow-dependent mechanotransduction has been described in great details in endothelial cells cultured in flow chambers as described in very well documented review articles [22, 35–37]. Integrins connects the extracellular matrix to the intracellular machinery. Integrins forming focal adhesion sites activate the focal adhesion kinase FAK and then a cascade of kinases including the MAP kinase pathway leading to cellular remodeling, cytoskeletal rearrangement and elongation of the cell in the direction of flow [35]. Although this phenomenon takes hours and involves cells initially cultured without flow these studies provide possible pathways to explore in the understanding of FMD. In cultured isolated blood vessels (aorta and carotid arteries) these pathways have been confirmed and more precisely described in “real” blood vessels [36, 38].
In resistance arteries less information is available. A role for integrins in FMD has been shown in rat coronary arterioles. Anti-β3-integrin antibodies or an antagonist to the matrix–integrin-binding site attenuates FMD [39]. This was associated with the inhibition of tyrosine phosphorylation of endothelial cells and with a reduced NO production [39]. The same group has also shown that FMD is reduced by the inhibition of the FAK-dependent signaling pathway. This inhibition of FAK prevents flow-induced activation of Akt and eNOS [40].
In addition to the β3 integrin involved in FMD in the rat coronary artery, blockade of the alpha-1 integrin with blocking antibodies severely reduced FMD in mesenteric arteries. Similarly, in mice lacking the alpha-1 integrin, FMD is strongly reduced in resistance arteries, although acetylcholine-induced dilation or contractility to pressure (myogenic tone) or adrenergic stimulation is not affected [41]. Interestingly, activation of the alpha-1 integrin by flow in the mesenteric artery is also associated with Akt activation upstream eNOS phosphorylation [41]. Nevertheless, the role of integrins in FMD remains to be better defined as only two vascular beds have been investigated and so far one cannot tell whether β3 integrins are involved in rat coronary arteries and alpha-1 integrins in mesenteric arteries only or in other resistance arteries. Whether tissue specificity exists and/or whether there is a cross talk between integrins in the different arterial beds is not known.
Besides integrins the dystrophin-dystroglycan complex may also play a role in FMD. Indeed, the lack of a functional dystrophin in mdx mice causes a specific defect in the transduction of shear stress into dilation through the NO-cGMP pathway in endothelial cells being able to dilate normally to other relaxing stimuli [6]. Dystrophin is a main cytoskeletal structure protein [42] involved in skeletal and cardiac muscle cells mechanotransduction. Indeed, dystrophin has a key position between membrane structure proteins and the actin cytoskeleton [43]. Dystrophin is not only expressed in muscular cells but also in both human and mouse vascular endothelial cells [6]. In mdx mice an incomplete and non-functional dystrophin is produced. In these animals both small and large arteries respond weakly to flow (shear stress) whereas dilation to acetylcholine, which also stimulates NO production by endothelial cells or dilation induced by the NO donor sodium nitroprusside, are normal [6]. The presence of dystrophin in endothelial cells was initially controversial. Nevertheless, its presence has been demonstrated in endothelial cells in mice, rats and humans [6]. In addition, it is located at the level of the plasma membrane in both endothelial and smooth muscle cells [6]. Surprisingly, pressure-induced (myogenic) tone as well as agonist-induced contractions were unaffected although dystrophin is a main structure protein in muscular cells, including smooth muscle cells. The role of dystrophin in FMD was confirmed using mdx mice treated with gentamycin. Gentamycin has the property to induce the recovery of the expression of a full and functional dystrophin in tissues as visualized using antibodies targeting different parts of the protein. In mdx mice treated with gentamycin for 2 weeks FMD was equivalent to control [44]. Related compounds with a higher efficiency and less toxicity are actually studied in order to find a treatment of some forms of myopathy [45]. This defect in FMD was also associated with an absence of adaptation of resistance arteries to a chronic rise in blood flow [9] (see below, paragraph B-2). Utrophin is structurally close to dystrophin and in many situations can compensate, at least in part for the absence of dystrophin. In human umbilical veins endothelial cells, utrophin forms a complex with beta- and epsilon-sarcoglycan, which associates with caveolin-1 and eNOS [46]. Nevertheless, the precise role of dystrophin and utrophin in endothelial cells in vivo remains little understood. In endothelial cells FMD has been suggested to involve a pathway different from that of agonist such as acetylcholine or bradykinin [47]. Thus, dystrophin, which may interact with caveolin-1 and eNOS, might be the first step in a pathway specific for FMD. Nevertheless, no direct link between these proteins has yet been shown in vascular endothelial cells, although in muscular cells the link between caveolin-3 and dystrophin is clear. In dystrophin-related myopathies vascular disorders are not determinant in the evolution of the diseases but they might at least contribute to extent of tissue damage [48].
Caveolin-1 is a structure protein with a key position in caveolae and it is associated to many proteins including eNOS. In endothelial cells chronically exposed to shear stress both caveolin and eNOS expression increase and cells become more sensitive to subsequent stimulation by flow [49]. In addition, FMD is improved in mice lacking caveolin-1, which link eNOS in an inactive state [34]. Thus, an attractive hypothesis is that the actors of the response to flow would be concentrated in caveolae, from integrins, dystroglycans and focal adhesion sites on one side to dystrophin, eNOS, HSPs, Akt on the other side. Among the proteins potentially associated to caveolae, PECAM-1 might also play a role in mechanotransduction. In gracilis muscle arterioles isolated from PECAM-1-deficient mice, FMD is strongly reduced [50]. PECAM-1 is a scaffold protein able to modulate eNOS activity [51]. Indeed, earlier studies of shear stress distribution in endothelial cell have demonstrated that tension is higher at the level of cell–cell junctions [52]. In addition, in endothelial cells exposed to increasing shear stress, proteins associated with cell–cell junction undergo a rapid reorganization [53]. Interestingly, polymorphisms in the gene encoding for PECAM-1 are associated to a higher risk of coronary artery disease [54].
Vimentin is probably another key element in the chain linking shear stress to vasodilatation although its position in the sequence is not yet known. Indeed, in mice lacking vimentin FMD is selectively reduced in resistance arteries [5]. In cultured cells, vimentin has been shown to be sensitive to mechanical factor [55] including shear stress [56]. The vimentin network is important in the stabilization of the cell. Indeed, in vimentin knockout mice, which have no obvious phenotype [57], regional defects at the cellular level may require a very precise mechanical organization as FMD [5, 58] and cell migration [59] are reduced in mice lacking vimentin. Thus, interactions between vimentin and other cytoskeletal elements may be critical in the vascular response to shear stress. Nevertheless, a direct observation and measurement of intermediate filaments position during flow has shown that short-term changes in flow do not induce assembly or disassembly of vimentin network segments [60]. On the other hand, vimentin rearrangement depends on PI3kinase stimulation, which has a key role in FMD as well as in the actin network reorganization. Vimentin has also been shown to regulate focal contact size when stimulated by shear stress [61]. This helps stabilizing cell-matrix adhesions in endothelial cells [61]. More generally, intermediate filaments are involved in the targeting of proteins to specific cellular locations, in the transport and distribution of organelles, and in the regulation of cell–cell and cell–matrix adhesion as recently described in a review article [62]. We have investigated the role of another main intermediate filament, desmin, in vascular tone. Nevertheless, desmin has no selective effect on FMD as its absence lead to a non-selective decrease in contractility and dilatory capacity of arteries [8]. Indeed, desmin is most likely involved in the contractility of smooth and skeletal muscle. Defects in the gene encoding for desmin induce rare but severe cardiomyopathies [44].
Thus, in the response of arteries to changes in blood flow the role of cytoskeletal or structure proteins is well accepted although it remains to be better defined and it remains many empty boxes. Nevertheless, changes in shear stress are likely activating extracellular elements within the glycocalyx transmitting the signal to membrane-anchored complexes. These latter might be different depending on cell type, species or environment. So far PECAM, integrins and the dystrophin-dystroglycan complex have been identified as the best candidates in this flow-dependent mechanotransduction. Nevertheless, their precise role and their interaction are not yet understood. The signal is then transmitted to eNOS through activation of FAK and PI3kinase-Akt. The location of vimentin in this pathway, although essential, remains to be determined.
1.2 Mechanotransduction in the microcirculation: chronic responses
1.2.1 Microcirculatory adaptation to chronic changes in blood flow
Whereas stimulation of the endothelium by shear stress induces a rapid vasodilatation, when the stimulus remains for a long period of time the arterial wall has to remodel in order to adapt to the new haemodynamic environment [63]. Remodeling due to a chronic rise in blood pressure is well known in both large and small arteries. This phenomenon has been widely studied in an effort to discover the vascular consequences of high blood pressure (see for review: [64, 65]). In resistance arteries high blood pressure triggers a structural and functional adaptation with changes in receptor density, type and distribution. On the other hand, remodeling due to chronic changes in blood flow has been mainly studied in large arteries allowing a better understanding of diseases affecting large arteries such as atherosclerosis or aneurysms [36, 66]. In the microcirculation the main difficulty is the size of the vessels. Nevertheless, most ischemic and metabolic diseases require angio- and arteriogenesis and a remodeling of the arterial bed. After chronic exercise training, which increases chronically blood flow, resistance artery diameter increases in association with an enhanced endothelial capacity to produce NO and prostanoids [67]. Similarly, chronic treatment of rats or mice with a non-selective vasodilator induces an increase in blood flow with consequences on the arterial wall structure and vasodilator capacity similar to those observed after exercise training [68]. Nevertheless in these models chronic changes in blood flow occur in parallel with changes in metabolism and hormonal environment. A more comprehensive model has been developed in the mesenteric circulation: arteries are alternatively ligated so that they are submitted to low, high or normal blood flow without changes in blood pressure, hormonal environment or body mass [69, 70]. In this model, chronic increases and decreases in blood flow induce outward and inward arterial remodeling, respectively (Fig. 1). Arterial wall remodeling is accompanied by a compensatory change in medial mass, which restores circumferential wall stress [70, 71]. A chronic increase in blood flow also increases the dilatory capacity of the endothelium, especially in response to shear stress [9, 72, 73]. Endothelial cells from high-flow arteries express more eNOS than control arteries [73–75] and the production of NO is critical for the diameter enlargement [74]. In addition, NO probably acts in association with reactive oxygen species (ROS) to form peroxynitrites (ONOO−), which in turn activate MMPs; although this has been shown in the carotid artery submitted to a much higher rise in shear stress due to a fistulae [76]. This has not been shown in resistance arteries yet, although preliminary data suggest a similar pathway in mesenteric resistance arteries (see next paragraph). Arterial wall hypertrophy associated with the increase in diameter might involve the local production of angiotensin II as either angiotensin I converting enzyme inhibition or angiotensin II type 1 receptor blockade prevented the hypertrophy without affecting the increase in diameter (Cousin and Henrion, unpublished data). This is in agreement with our previous works showing the involvement of the local renin-angiotensin system in the vascular response to shear stress [14, 77, 78] (Fig. 2).

Schematic representation of the model used to change chronically blood flow in mesenteric arteries in vivo. Arrows indicate location of the ligations of second-order mesenteric arterial branches. The artery located between two ligated vessels is designated as a high flow (HF) artery. Equivalent arteries located at distance of the ligatures are used as control or normal flow arteries (NF)

Schematic representation of the possible pathways involved in the acute response to flow (flow-mediated dilation, left panel) and in response to a chronic increase in blood flow (remodeling, right panel). Flow-mediated dilation involves the activation of extracellular structures, which stimulate the F-actin network and membrane-anchored complexes. Transmission of the signal to eNOS through activation of Akt and HSPs requires the presence of dystrophin and vimentin. Nevertheless their precise role remains to be elucidated. High flow-induced remodeling is made of a diameter increase involving the NO and MMPs pathways and of an arterial hypertrophy involving ROS and RhoA. The increase in diameter requires the production of both NO and ROS followed by the activation of MMPs. Arterial wall hypertrophy results of the activation of RhoA and MAP kinases. CAV1: caveolin-1, eNOS: endothelial NO-synthase, MMPs: metalloproteases, FAK: focal adhesion kinase; HSPs: heat shock proteins; ECM: extracellular matrix; AT1/2R: angiotensin II type 1 or 2 receptor
1.2.2 Role of structure proteins in the adaptation of the microcirculation to chronic changes in blood flow
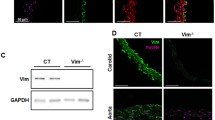
Considering our initial study showing that FMD was selectively impaired in vimentin-null mice [5], we submitted these animals to carotid artery ligation [7]. Whereas vimentin is essential in FMD (its absence selectively reduced FMD, see above), in response to a chronic increase in blood flow the absence of vimentin might oppose remodeling [7]. Indeed, intermediate filaments determine, together with actin and microtubules, cells architecture and shape. In addition, Intermediate filaments are generally considered the most rigid elements involved in the overall cell shape [79]. Thus intermediate filaments, which play a selective role in FMD (vimentin) or in vascular contractility (desmin), are probably regulating the remodeling process following a chronic change in blood flow.
First, using a model of carotid artery ligation we found that the contralateral artery submitted to high flow remodeled excessively in vimentin-null mice [7]. Similarly, in mice lacking desmin, mesenteric artery remodeling is higher than in control mice with a larger diameter increase [8]. These findings are in agreement with the concept that the intermediate filaments desmin and vimentin stabilize the arterial wall structure and thus oppose diameter expansion when chronically stimulated by shear stress. This is in accordance with the observation that vimentin content increases after exposition to high flow or pressure, thus strengthening cellular architecture [80]. In addition, a proteomic analysis performed in mesenteric resistance submitted chronically to high blood flow for 1 week shows a significant increase in vimentin content [81]. Interestingly, in mature rats (8–10 month-old) vimentin content was lower than in young animals and no diameter expansion occurred after a chronicle increase in blood flow [81]. In high flow resistance arteries several heat shock proteins (HSP) expression level increased as well, which is a characteristic of a response to a stress [81]. HSPs may be involved in the stabilization of cytoskeletal element after diameter expansion [82]. High flow-induced remodeling is also associated with eNOS overexpression and depends on NO production in resistance arteries [74, 81]. In addition, in large blood vessels [76] and most probably in resistance arteries (preliminary result: [83]), high flow-induced remodeling requires a rise in reactive oxygen species (ROS) production. Indeed, in these studies ROS scavenging [83] or NADPH oxidase p47 subunit knockout [76] prevents remodeling due to high flow. NO and ROS might form peroxynitrites, which would then activate MMPs [76]. In cultured endothelial cells shear stress activates ROS production [84], which stimulates tyrosine kinases [85]. Furthermore, in the same bovine aortic endothelial cells, pulsatile shear stress increases superoxides formation and upregulates NADPH oxidase (p22phox) gene expression [86]. Superoxides and NO are also able to activate HSPs production [87] and both are produced by endothelial cells in response to shear stress [47]. Enolase, which has properties similar to those of HSPs, is also increased in mesenteric resistance arteries submitted to high flow in young and mature rats [81]. In young rats diameter and medial mass increased whereas in mature rats medial hypertrophy occurred without increase in diameter [81].
Thus, flow-activated NO and ROS production would preferentially stimulate MMPs and diameter enlargement whereas HSPs would take part in wall hypertrophy. Nevertheless, basal ROS level might determine the type of response obtained after stimulation of endothelial cells by a chronic rise in blood flow. Indeed, in hypertensive rats (SHRs), mesenteric arteries submitted to high flow do not remodel (no increase in diameter) but the treatment of SHRs with the antioxidant tempol restores a significant remodeling [88]. Thus basal ROS level might determine the type of response obtained after a chronic increase in blood flow in rat mesenteric arteries. Nevertheless, this issue requires further investigations, especially in other diseases such as diabetes or obesity also characterized by high oxidative stress and altered endothelial function. It should be noted that in obese Zucker rats, basal ROS production is higher than in control lean rats and that increasing chronically blood flow in mesenteric arteries remain able to increase arterial diameter. This latter was accompanied by a rise in ROS production in both lean and obese rats [73].
By contrast with mice lacking vimentin or desmin, mice lacking a functional dystrophin (mdx mice) do not remodel in response to a chronic increase in flow [9] although FMD is reduced in mdx mice, as it is in mice lacking vimentin. Thus, both vimentin and dystrophin are activated by an acute rise in shear stress in a sequence starting outside the cells and being transmitted across the plasma membrane through a pathway that might involve dystrophin first and then intermediate filaments. The activation of this pathway leads to a rapid vasodilatation FMD) not requiring a change in cell architecture. In the vascular response to a chronic rise in blood flow vimentin and desmin oppose the structural changes required for the diameter expansion. This is probably due to their role in the maintenance of cell architecture. On the other hand, dystrophin, located exclusively beneath the plasma membrane, has a role in the transmission of the signal (rise in shear stress) to the cellular machinery (eNOS, NADPH-oxidase) needed for NO and ROS production, essential for high flow-induced diameter increase in resistance arteries.
That no remodeling was seen in mdx mice might be of importance in dystrophin-related myopathies such as Duchenne’s muscular dystrophy. Indeed, in situations such as ischemia, physical effort or wound healing, an inappropriate vascular wall adaptation to an increasing need in blood flow might be deleterious and increase tissue damages. So far it is considered that no important vascular disorder occurs in dystrophin-associated myopathies [48]. Nevertheless remodeling of the vasculature might be necessary in the process of skeletal and/or cardiac muscle regeneration using different approaches (review [89]). In a selective muscular therapy aiming to restore a functional dystrophin in patients the microcirculation would need a proper adaptation with diameter expansion of preexisting vessels in addition of angiogenesis. In this situation arteries lacking this capacity to adapt would reduce the efficacy of such a therapy.
Chronic changes in blood flow may also involve the stimulation of the glycocalyx and the transmission of the signal from the glycocalyx to the F-actin network located beneath the plasma membrane as described above for acute responses (FMD). Stimulation by shear stress is followed by a change in caveolar density, thus rendering the signal more selective for the NO-pathway [90]. This adaptation takes place between 1 and 3 days after a chronic change in flow. Nevertheless, no study has yet investigated the role of the glycocalyx specifically in high flow-induced remodeling. Similarly no study has yet investigated the role of the transmembrane complexes, especially as regard to the potential role of the integrins-FAK and Src pathway, which is likely to play a role in the process.
Activation of the structure proteins in response to a chronic rise in blood flow might also involve the RhoA/Rho kinase system. A first study has been performed in the model described in Fig. 1 using a cDNA microarray analysis (14,000 genes were analyzed). This analysis, confirmed by an Rt-PCR analysis showed an overexpression of RhoA and Rho kinase at day 4 after ligature followed a down-regulation at day 16 and 32 [91]. This is consistent with a role of the RhoA–Rho kinase system in high flow-induced remodeling. This is in agreement with previous studies showing that the RhoA/Rho kinase pathway is involved in many cellular activities besides the cytoskeleton including vesicle trafficking, endocytosis, cellular differentiation and gene transcription (review [92]). Although the RhoA–Rho kinase system has a central role in cell function including pathways activated by shear stress, a treatment with the Rho-kinase inhibitor Fasudil failed to prevent or to reduce high flow-remodeling [91], although the relatively low dose of fasudil used might not be high enough to block sufficiently the activity of the enzyme. In a model of chronic increase in flow in the pulmonary artery, a ten fold higher dose of fasudil prevented arterial wall hypertrophy [93]. Nevertheless, in this study the rise in flow was much higher than in the ligated mesenteric arteries (four fold increase versus 60%) and blood pressure probably increased substantially in the pulmonary circulation as the model is based on a carotid shunt [93]. Thus, the hypertrophy due to a rise in blood flow and pressure most probably involves the RhoA pathway. This is in accordance with previous studies demonstrating that fasudil is efficient against hypertension and the associated hypertrophy. In addition, RhoA is involved in the signaling pathway activated by trophic hormones involved in hypertrophy [94].
Thus, A chronic increase in blood flow in resistance arteries induces a rise in diameter associated with a medial hypertrophy. So far the pathway involved have been mainly investigated in first order mesenteric arteries. This is a limitation, although the mesenteric circulation has a key role in vascular homeostasis. Nevertheless, in the mesenteric arterial bed the existence of arcading arteries allows ligating alternatively second order arteries and thus to create low and high blood flow areas. In summary, the increase in blood flow induces overexpression of eNOS and NADPH-oxidase, which produce NO and ROS, both important for the activation of MMPs and diameter enlargement. Growth and migration of vascular cells probably results of the stimulation by shear stress of the local rennin–angiotensin system and RhoA inducing activation of MAP kinases needed for the process.
Abbreviations
- INSERM:
-
National Institute for Health and Medical Research
- CNRS:
-
National Center for Scientific Research
- FMD:
-
Flow-mediated dilation
- NO:
-
Nitric oxide
- ROS:
-
Reactive oxygen species
- ONOO−:
-
Peroxynitrites
- eNOS:
-
Endothelial NO-synthase
- FMD:
-
Flow-mediated dilation
- CAV1:
-
Caveolin-1
- MMPs:
-
Metalloproteases
- FAK:
-
Focal adhesion kinase
- HSPs:
-
Heat shock proteins
- ECM:
-
Extracellular matrix
- AT1/2R:
-
Angiotensin II type 1 or 2 receptor
References
Ingber DE (2002) Mechanical signaling and the cellular response to extracellular matrix in angiogenesis and cardiovascular physiology. Circ Res. 91(10):877–887
Kodama A, Lechler T, Fuchs E (2004) Coordinating cytoskeletal tracks to polarize cellular movements. J Cell Biol.167(2):203–207
Li S, Huang NF, Hsu S (2005) Mechanotransduction in endothelial cell migration. J Cell Biochem 96(6):1110–1126
Hutcheson IR, Griffith TM (1996) Mechanotransduction through the endothelial cytoskeleton: mediation of flow- but not agonist-induced EDRF release. Br J Pharmacol 118(3):720–726
Henrion D, Terzi F, Matrougui K, Duriez M, Boulanger CM, Colucci-Guyon E, Babinet C, Briand P, Friedlander G, Poitevin P, Levy BI (1997) Impaired flow-induced dilation in mesenteric resistance arteries from mice lacking vimentin. J Clin Invest 100(11):2909–2914
Loufrani L, Matrougui K, Gorny D, Duriez M, Blanc I, Levy BI, Henrion D (2001) Flow (shear stress)-induced endothelium-dependent dilation is altered in mice lacking the gene encoding for dystrophin. Circulation 103(6):864–870
Schiffers PM, Henrion D, Boulanger CM, Colucci-Guyon E, Langa-Vuves F, van Essen H, Fazzi GE, Levy BI, De Mey JG (2000) Altered flow-induced arterial remodeling in vimentin-deficient mice. Arterioscler Thromb Vasc Biol 20(3):611–616
Loufrani L, Li Z, Levy BI, Paulin D, Henrion D (2002) Excessive microvascular adaptation to changes in blood flow in mice lacking gene encoding for desmin. Arterioscler Thromb Vasc Biol 22(10):1579–1584
Loufrani L, Levy BI, Henrion D (2002) Defect in microvascular adaptation to chronic changes in blood flow in mice lacking the gene encoding for dystrophin. Circ Res 91(12):1183–1189
Bevan JA, Laher I (1991) Pressure and flow-dependent vascular tone. Faseb J 5(9):2267–2273
Bevan JA, Henrion D (1994) Pharmacological implications of the flow-dependence of vascular smooth muscle tone. Annu Rev Pharmacol Toxicol 34:173–190
Henrion D (2005) Pressure and flow-dependent tone in resistance arteries. Role of myogenic tone. Arch Mal Coeur Vaiss 98(9):913–921
Henrion D, Laher I, Laporte R, Bevan JA (1992) Angiotensin II amplifies arterial contractile response to norepinephrine without increasing Ca++ influx: role of protein kinase C. J Pharmacol Exp Ther 261(3):835–840
Henrion D, Benessiano J, Levy BI (1997) In vitro modulation of a resistance artery diameter by the tissue renin-angiotensin system of a large donor artery. Circ Res 80(2):189–195
Iglarz M, Levy BI, Henrion D (1998) Chronic endothelin-1-induced changes in vascular reactivity in rat resistance arteries and aorta. Eur J Pharmacol 359(1):69–75
Yeon DS, Kim JS, Ahn DS, Kwon SC, Kang BS, Morgan KG, Lee YH (2002) Role of protein kinase C- or RhoA-induced Ca(2+) sensitization in stretch-induced myogenic tone. Cardiovasc Res 53(2):431–438
Furchgott RF, Zawadzki JV (1980) The obligatory role of endothelial cells in the relaxation of arterial smooth muscle by acetylcholine. Nature 288(5789):373–376
Palmer RM, Ferrige AG, Moncada S (1987) Nitric oxide release accounts for the biological activity of endothelium-derived relaxing factor. Nature 327(6122):524–526
Feletou M, Vanhoutte PM (2006) Endothelial dysfunction: a multifaceted disorder (The Wiggers Award Lecture). Am J Physiol Heart Circ Physiol 291(3):H985–H1002
Aird WC (2007) Phenotypic heterogeneity of the endothelium: II. Representative vascular beds. Circ Res 100(2):174–190
Aird WC (2007) Phenotypic heterogeneity of the endothelium: I. Structure, function, and mechanisms. Circ Res 100(2):158–173
Davies PF, Spaan JA, Krams R (2005) Shear stress biology of the endothelium. Ann Biomed Eng 33(12):1714–1718
Bevan JA, Siegel G (1991) Blood vessel wall matrix flow sensor: evidence and speculation. Blood Vessels 28(6):552–556
Siegel G, Walter A, Kauschmann A, Malmsten M, Buddecke E (1996) Anionic biopolymers as blood flow sensors. Biosens Bioelectron 11(3):281–294
Henrion D, Bevan JA (1995) Magnitude of flow-induced contraction and associated calcium influx in the rabbit facial vein is dependent upon the level of extracellular sodium. J Vasc Res 32(1):41–48
Pohl U, Herlan K, Huang A, Bassenge E (1991) EDRF-mediated shear-induced dilation opposes myogenic vasoconstriction in small rabbit arteries. Am J Physiol 261(6 Pt 2):H2016–2023
Florian JA, Kosky JR, Ainslie K, Pang Z, Dull RO, Tarbell JM (2003) Heparan sulfate proteoglycan is a mechanosensor on endothelial cells. Circ Res 93(10):e136–e142
Nieuwdorp M, Meuwese MC, Vink H, Hoekstra JB, Kastelein JJ, Stroes ES (2005) The endothelial glycocalyx: a potential barrier between health and vascular disease. Curr Opin Lipidol 16(5):507–511
VanTeeffelen JW, Brands J, Jansen C, Spaan JA, Vink H (2007) Heparin impairs glycocalyx barrier properties and attenuates shear dependent vasodilation in mice. Hypertension 50(1):261–267
Weinbaum S, Zhang X, Han Y, Vink H, Cowin SC (2003) Mechanotransduction and flow across the endothelial glycocalyx. Proc Natl Acad Sci U S A 100(13):7988–7995
Morita T, Kurihara H, Maemura K, Yoshizumi M, Nagai R, Yazaki Y (1994) Role of Ca2+ and protein kinase C in shear stress-induced actin depolymerization and endothelin 1 gene expression. Circ Res 75(4):630–636
Cipolla MJ, Gokina NI, Osol G (2002) Pressure-induced actin polymerization in vascular smooth muscle as a mechanism underlying myogenic behavior. Faseb J 16(1):72–76
Sun D, Huang A, Sharma S, Koller A, Kaley G (2001) Endothelial microtubule disruption blocks flow-dependent dilation of arterioles. Am J Physiol Heart Circ Physiol 280(5):H2087–2093
Yu J, Bergaya S, Murata T, Alp IF, Bauer MP, Lin MI, Drab M, Kurzchalia TV, Stan RV, Sessa WC (2006) Direct evidence for the role of caveolin-1 and caveolae in mechanotransduction and remodeling of blood vessels. J Clin Invest 116(5):1284–1291
Davies PF (1995) Flow-mediated endothelial mechanotransduction. Physiol Rev 75(3):519–560
Lehoux S, Castier Y, Tedgui A (2006) Molecular mechanisms of the vascular responses to haemodynamic forces. J Intern Med 259(4):381–392
Chien S (2007) Mechanotransduction and endothelial cell homeostasis: the wisdom of the cell. Am J Physiol Heart Circ Physiol 292(3):H1209–1224
Lehoux S, Tedgui A (2005) Making up and breaking up: the tortuous ways of the vascular wall. Arterioscler Thromb Vasc Biol 25(5):892–894
Muller JM, Chilian WM, Davis MJ (1997) Integrin signaling transduces shear stress–dependent vasodilation of coronary arterioles. Circ Res 80(3):320–326
Koshida R, Rocic P, Saito S, Kiyooka T, Zhang C, Chilian WM (2005) Role of focal adhesion kinase in flow-induced dilation of coronary arterioles. Arterioscler Thromb Vasc Biol 25(12):2548–2553
Dumont O, Loufrani L, You D, Henrion D (2004) Involvement of alpha-1 integrins in the activation of akt and no-synthesis after flow (shear stress) activation of vasodilation in resistance arteries. J Vasc Res 41(suppl 1):58
Brown SC, Lucy JA (1993) Dystrophin as a mechanochemical transducer in skeletal muscle. Bioessays 15(6):413–419
Lapidos KA, Kakkar R, McNally EM (2004) The dystrophin glycoprotein complex: signaling strength and integrity for the sarcolemma. Circ Res 94(8):1023–1031
Loufrani L, Dubroca C, You D, Li Z, Levy B, Paulin D, Henrion D (2004) Absence of dystrophin in mice reduces NO-dependent vascular function and vascular density: total recovery after a treatment with the aminoglycoside gentamicin. Arterioscler Thromb Vasc Biol 24(4):671–676
Aurino S, Nigro V (2006) Readthrough strategies for stop codons in Duchenne muscular dystrophy. Acta Myol 25(1):5–12
Ramirez-Sanchez I, Ceballos-Reyes G, Rosas-Vargas H, Cerecedo-Mercado D, Zentella-Dehesa A, Salamanca F, Coral-Vazquez RM (2007) Expression and function of utrophin associated protein complex in stretched endothelial cells: dissociation and activation of eNOS. Front Biosci 12:1956–1962
Fleming I, Busse R (2003) Molecular mechanisms involved in the regulation of the endothelial nitric oxide synthase. Am J Physiol Regul Integr Comp Physiol 284(1):R1–12
Deconinck N, Scaillon M, Segers V, Groswasser JJ, Dan B (2006) Opsoclonus-myoclonus associated with celiac disease. Pediatr Neurol 34(4):312–314
Rizzo V, Morton C, DePaola N, Schnitzer JE, Davies PF (2003) Recruitment of endothelial caveolae into mechanotransduction pathways by flow conditioning in vitro. Am J Physiol Heart Circ Physiol 285(4):H1720–H1729
Bagi Z, Frangos JA, Yeh JC, White CR, Kaley G, Koller A (2005) PECAM-1 mediates NO-dependent dilation of arterioles to high temporal gradients of shear stress. Arterioscler Thromb Vasc Biol 25(8):1590–1595
Dusserre N, L’Heureux N, Bell KS, Stevens HY, Yeh J, Otte LA, Loufrani L, Frangos JA (2004) PECAM-1 interacts with nitric oxide synthase in human endothelial cells: implication for flow-induced nitric oxide synthase activation. Arterioscler Thromb Vasc Biol 24(10):1796–1802
Liu SQ, Yen M, Fung YC (1994) On measuring the third dimension of cultured endothelial cells in shear flow. Proc Natl Acad Sci U S A 91(19):8782–8786
Seebach J, Dieterich P, Luo F, Schillers H, Vestweber D, Oberleithner H, Galla HJ, Schnittler HJ (2000) Endothelial barrier function under laminar fluid shear stress. Lab Invest 80(12):1819–1831
Listi F, Candore G, Lio D, Cavallone L, Colonna-Romano G, Caruso M, Hoffmann E, Caruso C (2004) Association between platelet endothelial cellular adhesion molecule 1 (PECAM-1/CD31) polymorphisms and acute myocardial infarction: a study in patients from Sicily. Eur J Immunogenet 31(4):175–178
Thoumine O, Ziegler T, Girard PR, Nerem RM (1995) Elongation of confluent endothelial cells in culture: the importance of fields of force in the associated alterations of their cytoskeletal structure. Exp Cell Res 219(2):427–441
Cucina A, Sterpetti AV, Pupelis G, Fragale A, Lepidi S, Cavallaro A, Giustiniani Q, Santoro D’Angelo L (1995) Shear stress induces changes in the morphology and cytoskeleton organisation of arterial endothelial cells. Eur J Vasc Endovasc Surg 9(1):86–92
Colucci-Guyon E, Portier MM, Dunia I, Paulin D, Pournin S, Babinet C (1994) Mice lacking vimentin develop and reproduce without an obvious phenotype. Cell 79(4):679–694
Terzi F, Henrion D, Colucci-Guyon E, Federici P, Babinet C, Levy BI, Briand P, Friedlander G (1997) Reduction of renal mass is lethal in mice lacking vimentin. Role of endothelin-nitric oxide imbalance. J Clin Invest 100(6):1520–1528
Eckes B, Dogic D, Colucci-Guyon E, Wang N, Maniotis A, Ingber D, Merckling A, Langa F, Aumailley M, Delouvee A, Koteliansky V, Babinet C, Krieg T (1998) Impaired mechanical stability, migration and contractile capacity in vimentin-deficient fibroblasts. J Cell Sci 111(Pt 13):1897–1907
Helmke BP, Goldman RD, Davies PF (2000) Rapid displacement of vimentin intermediate filaments in living endothelial cells exposed to flow. Circ Res 86(7):745–752
Tsuruta D, Jones JC (2003) The vimentin cytoskeleton regulates focal contact size and adhesion of endothelial cells subjected to shear stress. J Cell Sci 116(Pt 24):4977–4984
Kim S, Coulombe PA (2007) Intermediate filament scaffolds fulfill mechanical, organizational, and signaling functions in the cytoplasm. Genes Dev 21(13):1581–1597
Mulvany MJ (1999) Vascular remodelling of resistance vessels: can we define this? Cardiovasc Res 41(1):9–13
Prewitt RL, Rice DC, Dobrian AD (2002) Adaptation of resistance arteries to increases in pressure. Microcirculation 9(4):295–304
Mulvany MJ (2002) Small artery remodeling and significance in the development of hypertension. News Physiol Sci 17:105–109
Langille BL (1996) Arterial remodeling: relation to hemodynamics. Can J Physiol Pharmacol 74(7):834–841
Koller A, Huang A, Sun D, Kaley G (1995) Exercise training augments flow-dependent dilation in rat skeletal muscle arterioles. Role of endothelial nitric oxide and prostaglandins. Circ Res 76(4):544–550
Gorny D, Loufrani L, Kubis N, Levy BI, Henrion D (2002) Chronic hydralazine improves flow (shear stress)-induced endothelium-dependent dilation in mouse mesenteric resistance arteries in vitro. Microvasc Res 64(1):127–134
Unthank JL, Fath SW, Burkhart HM, Miller SC, Dalsing MC (1996) Wall remodeling during luminal expansion of mesenteric arterial collaterals in the rat. Circ Res 79(5):1015–1023
Pourageaud F, De Mey JG (1997) Structural properties of rat mesenteric small arteries after 4-wk exposure to elevated or reduced blood flow. Am J Physiol 273(4 Pt 2):H1699–H1706
Buus CL, Pourageaud F, Fazzi GE, Janssen G, Mulvany MJ, De Mey JG (2001) Smooth muscle cell changes during flow-related remodeling of rat mesenteric resistance arteries. Circ Res 89(2):180–186
Pourageaud F, De Mey JG (1998) Vasomotor responses in chronically hyperperfused and hypoperfused rat mesenteric arteries. Am J Physiol 274(4 Pt 2):H1301–H1307
Bouvet C, de Chantemele EB, Guihot AL, Vessieres E, Bocquet A, Dumont O, Jardel A, Loufrani L, Moreau P, Henrion D (2007) Flow-induced remodeling in resistance arteries from obese Zucker rats is associated with endothelial dysfunction. Hypertension 50(1):248–254
Dumont O, Loufrani L, Henrion D (2007) Key role of the NO-pathway and matrix metalloprotease-9 in high blood flow-induced remodeling of rat resistance arteries. Arterioscler Thromb Vasc Biol 27(2):317–324
Tulis DA, Unthank JL, Prewitt RL (1998) Flow-induced arterial remodeling in rat mesenteric vasculature. Am J Physiol 274(3 Pt 2):H874–H882
Castier Y, Brandes RP, Leseche G, Tedgui A, Lehoux S (2005) p47phox-dependent NADPH oxidase regulates flow-induced vascular remodeling. Circ Res 97(6):533–540
Matrougui K, Loufrani L, Heymes C, Levy BI, Henrion D (1999) Activation of AT(2) receptors by endogenous angiotensin II is involved in flow-induced dilation in rat resistance arteries. Hypertension 34(4 Pt 1):659–665
Matrougui K, Levy BI, Henrion D (2000) Tissue angiotensin II and endothelin-1 modulate differently the response to flow in mesenteric resistance arteries of normotensive and spontaneously hypertensive rats. Br J Pharmacol 130(3):521–526
Etienne-Manneville S (2004) Actin and microtubules in cell motility: which one is in control? Traffic 5(7):470–477
Schnittler HJ, Schmandra T, Drenckhahn D (1998) Correlation of endothelial vimentin content with hemodynamic parameters. Histochem Cell Biol 110(2):161–167
Tuttle JL, Hahn TL, Sanders BM, Witzmann FA, Miller SJ, Dalsing MC, Unthank JL (2002) Impaired collateral development in mature rats. Am J Physiol Heart Circ Physiol 283(1):H146–H155
Li S, Piotrowicz RS, Levin EG, Shyy YJ, Chien S (1996) Fluid shear stress induces the phosphorylation of small heat shock proteins in vascular endothelial cells. Am J Physiol 271(3 Pt 1):C994–C1000
de Chantemele EB, Vessieres E, Guihot AL, Dumont O, Loufrani L, Henrion D (2007) Reactive oxygen species have a central role in flow (shear stress)-induced remodeling in rat mesenteric resistance arteries. Hypertension (in press)
Laurindo FR, Pedro Mde A, Barbeiro HV, Pileggi F, Carvalho MH, Augusto O, da Luz PL (1994) Vascular free radical release. Ex vivo and in vivo evidence for a flow-dependent endothelial mechanism. Circ Res 74(4):700–709
Tai LK, Okuda M, Abe J, Yan C, Berk BC (2002) Fluid shear stress activates proline-rich tyrosine kinase via reactive oxygen species-dependent pathway. Arterioscler Thromb Vasc Biol 22(11):1790–1796
Silacci P, Desgeorges A, Mazzolai L, Chambaz C, Hayoz D (2001) Flow pulsatility is a critical determinant of oxidative stress in endothelial cells. Hypertension 38(5):1162–1166
Xu Q, Hu Y, Kleindienst R, Wick G (1997) Nitric oxide induces heat-shock protein 70 expression in vascular smooth muscle cells via activation of heat shock factor 1. J Clin Invest 100(5):1089–1097
Miller SJ, Norton LE, Murphy MP, Dalsing MC, Unthank JL (2007) The role of the renin-angiotensin system and oxidative stress in spontaneously hypertensive rat mesenteric collateral growth impairment. Am J Physiol Heart Circ Physiol 292(5):H2523–H2531
McNally EM (2007) New approaches in the therapy of cardiomyopathy in muscular dystrophy. Annu Rev Med 58:75–88
Thi MM, Tarbell JM, Weinbaum S, Spray DC (2004) The role of the glycocalyx in reorganization of the actin cytoskeleton under fluid shear stress: a “bumper-car” model. Proc Natl Acad Sci U S A 101(47):16483–16488
Wesselman JP, Kuijs R, Hermans JJ, Janssen GM, Fazzi GE, van Essen H, Evelo CT, Struijker-Boudier HA, De Mey JG (2004) Role of the Rhoa/Rho kinase system in flow-related remodeling of rat mesenteric small arteries in vivo. J Vasc Res 41(3):277–290
Tzima E (2006) Role of small GTPases in endothelial cytoskeletal dynamics and the shear stress response. Circ Res 98(2):176–185
Li F, Xia W, Li A, Zhao C, Sun R (2007) Long-term inhibition of Rho kinase with fasudil attenuates high flow induced pulmonary artery remodeling in rats. Pharmacol Res 55(1):64–71
Loirand G, Guerin P, Pacaud P (2006) Rho kinases in cardiovascular physiology and pathophysiology. Circ Res 98(3):322–334
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article
Cite this article
Loufrani, L., Henrion, D. Role of the cytoskeleton in flow (shear stress)-induced dilation and remodeling in resistance arteries. Med Biol Eng Comput 46, 451–460 (2008). https://doi.org/10.1007/s11517-008-0306-2
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s11517-008-0306-2