
Abstract
The splenic response to stroke is a proinflammatory reaction to ischemic injury resulting in expanded neurodegeneration. Splenectomy reduces neural injury in rodent models of hemorrhagic and ischemic stroke, however the exact nature of this response has yet to be fully understood. This study examines the migration of splenocytes after brain ischemia utilizing carboxyfluorescein diacetate succinimidyl ester (CFSE) to label them in vivo. The spleen was found to significantly decrease in size from 24 to 48 h following middle cerebral artery occlusion (MCAO) in rats compared to sham operated controls. By 96 h post-MCAO the spleen size returned to levels not different from sham operated rats. To track splenocyte migration following MCAO, spleens were injected with CFSE to label cells. CFSE positive cell numbers were significantly reduced in the 48 h MCAO group versus 48 h sham and CFSE labeled cells were equivalent in 96 h MCAO and sham groups. A significant increase of labeled lymphocyte, monocytes, and neutrophils was detected in the blood at 48 h post-MCAO when compared to the other groups. CFSE labeled cells migrated to the brain following MCAO but appear to remain within the vasculature. These cells were identified as natural killer cells (NK) and monocytes at 48 h and at 96 h post-MCAO NK cells, T cells and monocytes. After ischemic injury, splenocytes enter into systemic circulation and migrate to the brain exacerbating neurodegeneration.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
Stroke is a complex neural injury that progresses through several phases following the initial insult. The peripheral immune system plays a delayed role in the progression of neural injury following stroke. The presence of peripheral immune cells in the brain following ischemic stroke has been well documented in animal studies (Schroeter et al. 1994; Stevens et al. 2002). The role that these cells play in the progression of neural injury following stroke is still being investigated, as the origin of these cells is not clearly understood. Recent evidence indicates that the spleen is at least one of the sources of these immune cells.
The spleen is a large reservoir of immune and red blood cells. Its removal reduces neurodegeneration in a variety of brain injury models (Ajmo et al. 2008; Lee et al. 2008; Li et al. 2011; Das et al. 2011). Moreover, splenectomy decreases cellular death following ischemic-reperfusion injury in other organs, including the liver (Okuaki et al. 1996), intestines (Savas et al. 2003), kidney (Jiang et al. 2007), and heart (Leuschner et al. 2010). Splenectomy also decreases the number of immune cells in the brain (Ajmo et al. 2008; Seifert et al. 2012) following middle cerebral artery occlusion (MCAO). Additional studies have shown that preventing peripheral immune cell infiltration is neuroprotective following MCAO. Antibodies generated against cellular adhesion molecules prevent immune cell extravasation into the brain and decrease infarct volume (Chopp et al. 1996; Kanemoto et al. 2002; Liesz et al. 2011). Cytokine production within the brain following MCAO is also decreased with splenectomy (Seifert et al. 2012). These studies illustrate that the spleen is a focal point for the immune response to tissue injury.
Peripheral immune cells produce and secrete proinflammatory cytokines while residing in the ischemic brain. Several cytokines and chemokines have up regulated mRNA expression in the brain following transient MCAO in mice. (Chang et al. 2011; Hurn et al. 2007; Offner et al. 2006a) Elevated levels of interferon gamma (IFNγ) protein have been found in the brain following permanent MCAO in rats (Seifert et al. 2012). Additionally, intracerebroventricular injection of antibodies directed against either tumor necrosis factor alpha (TNFα) or IFNγ following MCAO decreased infarct volume (Liesz et al. 2009). These data demonstrate that the presence of peripheral immune cells is detrimental following stroke, whether these cells are acting through a cellular cytotoxic mechanism and/or through the secretion of proinflammatory cytokines and chemokines. Elucidating the source of these cells should provide some insight into the mechanisms by which these cells elicit a proinflammatory response following ischemic brain injury.
This study aims to determine temporal changes in splenic mass in rats following MCAO to determine if splenic contraction leads to an increase in circulating splenocytes. It has been well documented that there are peripheral immune cells present in the brain hours to days following MCAO (Stevens et al. 2002), however the origin of these cells remains to be elucidated. The spleen contributes to the proinflammatory response following MCAO and is a major reservoir of immune cells. Using carboxyfluorescein diacetate succinimidyl ester (CFSE), a fluorescent molecule that covalently binds to intracellular proteins, allowed splenocytes to be tracked in this study. CFSE is used in lymphocyte trafficking studies in vivo (Weston and Parish 1990) and can be detected for up to 8–10 cell divisions (Lyons 2000). It has also been shown to be detected out to 20 days in transplanted hepatocytes (Karrer et al. 1992). This suggests CFSE is a stable molecule that allows cells to be tracked using fluorescence. By labeling splenocytes in vivo with CFSE prior to MCAO, splenocyte migration was tracked following MCAO.
Materials and methods
Animal care
All animal procedures were conducted in accordance with the NIH Guide for the Care and Use of Laboratory Animals with a protocol approved by the Institutional Animal Care and Use Committee at the University of South Florida. Male Sprague-Dawley rats (300–350 g) were used for the in vivo experiments. All rats were purchased from Harlan Labs (Indianapolis, IN), maintained on a 12 h light/dark cycle (6 am–6 pm) and given access to food and water ad libitum.
Splenic CFSE injections
Splenic CFSE injections were performed five days prior to MCAO by making a midline skin incision at the caudal terminus of the 13th rib on the anatomical left. The abdominal wall was opened along midline and the spleen was externalized. The spleens were injected with 250 μl of a 4 mg/ml solution of CFSE (Molecular Probes, Eugene, OR) in dimethyl sulfoxide (DMSO). The injections were evenly spaced out in five sites of 50 μl per site. The spleen was then reinserted into the abdominal cavity. The abdominal cavity and then the skin incision were closed with sutures. To obtain baseline CFSE labeling, a group of CFSE only rats were euthanized five days following the splenic injections; at the same time point the other groups underwent MCAO or sham surgeries.
Laser doppler blood flow measurement
Laser Doppler was used to monitor blood perfusion (Moor Instruments Ltd, Devon, England) as previously described (Ajmo et al. 2006; Ajmo et al. 2008; Hall et al. 2009). Rats that did not show ≥60 % reduction in perfusion during MCAO were excluded from this study. Sham operated rats had the guide screw and laser doppler probe placed and blood flow was monitored to ensure that there was not a drop in cerebral blood flow during the sham procedure.
Permanent middle cerebral artery occlusion
MCAO surgery was performed using the intraluminal method originally described by Longa et al. (Longa et al. 1989) and as previously reported (Ajmo et al. 2006; Ajmo et al. 2008; Hall et al. 2009). Sham operations were performed for the CFSE rats at 48 and 96 h post-sham MCAO to ensure there no confounding factors from the CFSE injections.
Tissue extraction and sectioning
The animals were euthanatized with ketamine/xylazine mix, 75 mg/kg and 7.5 mg/kg respectively, intraperitoneal (i.p.) at 3, 24, 48, 51, 72 and 96 h post-MCAO, and perfused transcardially with 0.9 % saline followed by 4 % paraformaldehyde in phosphate buffer (PB). The spleens and thymi were removed prior to perfusion. Spleens were weighed immediately following removal and were subsequently placed in RPMI complete (RPMI-1640 without Phenol Red media (Cellgro, Manassas, VA), antibiotic/antimycotic (Cellgro), 29.2 mg/ml L-glutamine, low IgG FBS (Cellgro), and 14.2 M 2-mercaptoethanol) cell culture media. The thymi were placed in 4 % paraformaldehyde in PB. Thymi were subsequently sliced into 16 μm sections with a cryostat and thaw mounted on glass slides. The brains were harvested, post fixed in 4 % paraformaldehyde, and cryopreserved in sucrose solutions. Brains were sliced into 30 μm sections with a cryostat. Coronal sections were taken at six points from 1.7 to −3.3 mm from Bregma. Sections were either thaw mounted on glass slides and stored at −20 °C. All tissue processing was done under limited lighting to protect the CFSE dye.
Fluoro-Jade staining and infarct quantification
Brain sections mounted on glass slides were stained with Fluoro-Jade (Histochem, Jefferson, AR), which labels degenerating neurons. This method was adapted from that originally developed by Schmued et al. (Schmued et al. 1997) and as previously described (Duckworth et al. 2005; Ajmo et al. 2009; Seifert et al. 2012).
Fluoro-Jade stained tissue was digitally photographed with Zeiss Axioskop2 (Carl Zeiss Inc, Thornwood, NY) microscope controlled by Openlab software (Improvision, Waltham, MA) at a magnification of 1x. Area of neurodegeneration was measured using the NIH ImageJ software. To compensate for possible edema in the ipsilateral hemisphere, the area of the contralateral hemisphere was also measured. Infarct volumes were then calculated by the total area of ipsilateral staining divided by the total contralateral area for a given animal. Infarct quantification was done for all animals.
Immunohistochemistry and image capture
Immunohistochemistry was performed, as previously described, using fluorescence and peroxidase staining (Seifert et al. 2012). Double-labeled immunohistochemistry, for CFSE and immune cell surface markers was achieved by incubating the slides with primary antibodies, followed by incubation with secondary antibodies conjugated to 594 nm fluorophores as CFSE fluoresces at 488 nm.
The following primary antibodies were used: mouse anti-rat CD3 for T cells (1:2,000; BD Biosciences, San Jose, CA), mouse anti-rat CD161 for NK cells (1:1,000; Serotec, Raleigh, NC), and mouse anti-rat CD11b for microglia/macrophages (1:3,000; Serotec). Anti-fluorescein antibodies were used to detect the presence of any CFSE molecules: Alexa-Fluor® 488 goat-anti fluorescein (1:1,000; Invitrogen, Carlsbad, CA) and biotinylated rabbit anti-fluorescein (1:3,000; Invitrogen). Alexa-Fluor® 594 rabbit anti-mouse (1:300; Invitrogen) secondary was used in conjunction the immune cell surface markers.
Tissue sections that were double labeled were viewed on the Leica SP2 confocal microscope (Leica Microsystems, Buffalo Grove, IL). Images were taken at a magnification of 63x. Each fluorophore was scanned sequentially and then the two images were merged.
Splenic cell counts
Fresh spleens from the CFSE injected rats were placed in RPMI complete in stomacher bags and were disassociated then strained with a 70 μm filter. After straining through a 70 μm filter the volume was brought up to 40 ml. The cells were pelleted by centrifuging at 1500 rpm for 5 min. The supernatant was discarded and the cells were resuspended in 5 ml of ACK lysis buffer (0.15 M NH4Cl, 1 M KHCO3, 0.1 M Na2EDTA in water pH equaled 7.4) for 5 min. The volume was adjusted to 40 ml with RPMI complete and then centrifuged at 1500 rpm for 5 min. The supernatant was discarded and the cells were resuspended in 30 ml RPMI complete. Cells were counted using Trypan Blue exclusion of dead cells. Additionally, 40 μl of the solution was placed on a slide and cover slipped with a 22 mm circular cover slip (380 mm2), three slides were used per spleen to obtain CFSE cell counts for each spleen. Using a reticle with a 10×10 grid (1 mm2 area with 10x objective using Zeiss AxioSkop2 microscope, Carl Zeiss Inc) cells were counted in three different areas on each cover slip. A total number of CFSE positive cells and total number of cells for each area was recorded and then the nine total counts for each rat were averaged to give total number of CFSE positive cells and the percentage stained with CFSE for each time point.
Giemsa staining and analysis
Giemsa staining was used to identify the immune cell types within circulation following MCAO. Blood samples were obtained using cardiac puncture at the time of euthanasia. Blood smears were made on by placing a small drop of blood on a non-charged slide. The Giemsa (Sigma-Aldrich, St. Louis, MO) staining was performed as previously described (Ajmo et al. 2008). These slides were then used to count the total number of CFSE positive cells on the slide and identify these cells, as well as get a leukocyte count for each animal.
The total number of CFSE cells was determined by counting the number of cells on the total slide, two slides per animal. The cells were also indentified using the Giemsa staining. Additionally leukocytes were counted by indentifying 100 cells per slide, two slides per animal.
Statistical analysis
All data are expressed as group mean ± SEM. Significance of the data was determined by ANOVA with a Dunnet’s post hoc test for the spleen weights and the CFSE cells in the blood. A Tukey’s post hoc test was used for the CFSE cells in the spleen. A value of p < 0.05 was considered significant. All sections were blinded prior being analyzed by an investigator.
Results
The spleen transiently decreases in size following MCAO in rats
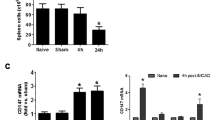
Spleens were weighed at different time points following MCAO. Spleen weights were measured at 3, 24, 48, 51, 72, and 96 h post-MCAO and sham procedure. Previously we have demonstrated 96 h post-MCAO as the time point at which the infarct is stable in this permanent model of MCAO (Newcomb et al. 2006). The spleen was found to significantly decrease in size at 24, 48, and 51 h following MCAO compared to sham operated animals (p < 0.05). However, spleen weights were not significantly different from sham controls at 3, 72 and 96 h post-MCAO (Fig. 1a).

Splenic response to MCAO. Mean spleen weights plotted over time following MCAO. Spleen weights were significantly decreased by the 24, 48, and 51 h time points compared to sham operated controls (*p < 0.05). Spleen weights were not significantly different from sham operated controls at the 3, 72, and 96 h time points (a). The total number of CFSE positive splenocytes was decreased in the 48 h MCAO group compared to the 48 h sham and the CFSE only groups. No differences were detected between the 96 h MCAO and sham groups, which were also significantly different from the 48 h sham and CFSE only groups (*p < 0.0001). The CFSE only group were rats that were euthanized 5 days post injection, the same time the other groups underwent sham or MCAO procedures (n ≥ 4) (b). Brain sections from CFSE treated rats at 48 and 96 h post-MCAO were immunostained with antibodies that recognize CD161 (NK cell), CD11b (monocytes), and CD3 (T cell). CD161 expression co-localized with CFSE labeled cells at 48 h (c) and 96 h (e). CD11b co-localized with CFSE labeled cells at 48 h (d) and 96 h (f). CD3 immunoreactivity was only detected with CFSE at 96 h (g). Arrows indicate double labeled cells. Scale bars = 50 μm
Changes in the number of CFSE positive cells within the spleen following MCAO
The total number of CFSE positive splenocytes was decreased in the 48 h MCAO group compared to the 48 h sham and the CFSE only groups. No differences were detected between the 96 h MCAO and sham groups, which were also significantly different from the 48 h sham and CFSE only groups (p < 0.0001).
Identification of CFSE positive in the brain following MCAO
To identify the types of CFSE labeled cells in the brain following MCAO, double labeling with staining for immune cell surface markers was employed. The double labeling demonstrated the presence of CFSE positive monocytes (CD11b) and NK cells (CD161) at 48 h post-MCAO (Fig. 1c and d). At 96 h following MCAO monocytes, NK cells and T cells (CD3) were present (Fig. 1e–g). There were not any CFSE positive T cells in the brains of animals 48 h post-MCAO (data not shown).
The percentage of CFSE cell labeling and MCAO induces changes in the number of CFSE positive cells within the circulation
CFSE labeled approximately 20 percent of splenocytes by five days. Circulating CFSE positive cells were identified by using blood smears and the cell types were determined using Giemsa staining. When blood smears from all the different groups were compared, an overall increase in CFSE positive cells was observed in circulation 48 h following MCAO (p < 0.0007). This increase resulted in an increase in circulating lymphocytes (p < 0.005), neutrophils (p < 0.0005), and monocytes (p < 0.02). Giemsa staining revealed a significant increase in lymphocytes at 48 h post-MCAO and in the 96 h sham and MCAO groups (p < 0.001). Neutrophils were significantly increased in the 48 h MCAO and 96 h sham groups (p < 0.001). In contrast, monocytes were significantly decreased in all groups compared to the CFSE only (p < 0.0001) (Table 1).
Discussion
The spleen is a highly proinflammatory organ following ischemic brain injuries (Ajmo et al. 2008; Lee et al. 2008; Li et al. 2011; Das et al. 2011) as well as ischemic liver (Okuaki et al. 1996), intestine (Savas et al. 2003), kidney (Jiang et al. 2007), and heart (Leuschner et al. 2010) injury. In animal studies splenectomy reduces injury in all of these organs. The spleen uses both a cellular and humoral response to tissue injuries. The humoral response has been characterized following stroke as there are many reports citing the splenic production of inflammatory cytokines and chemokines after this type of brain injury. These cytokines include increased IFNγ protein following MCAO in rats (Seifert et al. 2012). Increased cytokine mRNA for interleukin 1 beta (IL-1β) (Schwarting et al. 2008; Lee et al. 2008), TNFα (Lee et al. 2008; Schwarting et al. 2008; Hurn et al. 2007), IFNγ (Hurn et al. 2007), and IL-6 (Hurn et al. 2007; Lee et al. 2008) have been found in mice or rats following ischemic or hemorrhagic stroke. Chemokine mRNA for CXCL2 and CXCL10 have been found to be elevated following MCAO in mice (Offner et al. 2006a; Hurn et al. 2007). The cellular response has not been characterized as well as the humoral response following stroke. There are some data in a mouse model of transient MCAO (Offner et al. 2006b). However, cellular tracking has not been utilized following stroke.
Other investigators have found that the spleen decreases in size following transient MCAO in mice (Bao et al. 2010) and this decrease in spleen size is due to apoptosis of the cells and a loss of functional centers within the spleen (Offner et al. 2006b). In rats, the spleen is decreased in size 48 h following permanent MCAO and infarct volume was found to be inversely proportional to splenic weight (Vendrame et al. 2006). An extended time course demonstrated permanent MCAO results in a transient decrease in spleen size from 24–72 h post-MCAO. This effect is the result of a catecholamine surge following MCAO (Meyer et al. 2004) which activates α1-adrenergic receptors on the splenic capsule causing a contraction of the smooth muscles in the capsule. This leads to the transient decrease in splenic size observed in rats. Blocking the α1-adrenergic receptors with prazosin or carvedilol prevents the decrease in spleen size seen at 48 h following MCAO (Ajmo et al. 2009) while carvedilol is also neuroprotective (Savitz et al. 2000; Ajmo et al. 2009). Contraction of the splenic smooth muscle is associated with an increase in circulating white blood cells and erythrocytes (Bakovic et al. 2005). The splenocytes which are released into circulation following MCAO could account for the peripheral immune cells found in the brain after stroke. While there are differences between Offner et al. (2006b) and the current findings this could be due the different species used in the studies, mice versus rats, or the type of stroke surgery performed in each, transient MCAO versus permanent MCAO. Further investigations into how the spleen reacts to ischemic strokes in patients will help elucidate the role the spleen plays in stroke pathogenesis.
The spleen responds to MCAO induced injury differently than injury associated with a sham surgery. Sham operated rats at 48 h post-surgery have CFSE positive cell populations within the spleen not different than CFSE only rats. MCAO rats in both the 48 h and 96 h groups and those in the 96 h sham group all have significantly decreased CFSE positive cell populations within the spleen compared to CFSE only and 48 h sham groups. This indicates that the brain injury induces a different splenic response, as CFSE labeled cells are decreased in the spleen at 48 h post-MCAO compared to the 48 h sham operated group. The significant increase in circulating cells seen at 48 h post-MCAO demonstrates the injury to the brain induces splenocyte migration. Another study also found a decrease in splenocyte counts at 48 h post-MCAO compared to sham operated rats (Gendron et al. 2002). However, the sham operation induces a delayed splenic response as there is not a significant difference in the number of CFSE positive cells within the spleen at 96 h in the sham or MCAO treated groups. This indicates that the spleen does have a delayed response to a general insult like a sham operation. The splenic response to the sham operation at 96 h is also supported by the similar increases in circulating lymphocytes in both the 96 h MCAO and the 96 h sham groups.
Interestingly, while the spleen is decreased in size 48 h following MCAO, there is a significant increase in the number of circulating CFSE positive cells, regardless of immune cell type. The total number of circulating monocytes is decreased in all treatment groups compared to the CFSE only group, which was used as a CFSE injection control group. While the total number of circulating monocytes is decreased at 48 h there was a significant increase in CFSE positive monocytes demonstrating most of the monocytes in circulation following MCAO at 48 h are from the spleen. This is also consistent with evidence that there is a population of undifferentiated monocytes in the spleen that are released into circulation and travel to the heart following myocardial infarction in mice (Swirski et al. 2009). In a transient mouse MCAO model an increase in circulating monocytes was observed at 96 h following MCAO (Offner et al. 2006b), which is inconsistent with the data presented in this study. The differences could be due to the different models of MCAO and species used for each study. Additionally, few CFSE positive cells were found in the thymus (data not shown).
Circulating CFSE positive cells were found only in the brains of MCAO treated rats at 48 and 96 h following MCAO. There were no CFSE positive cells in the brains of sham-operated rats at any time point. These CFSE positive cells were identified as NK cells and monocytes at 48 h following MCAO, while at 96 h post-MCAO T cells were also present. The cells appear to be localized to the blood vessels. While there is clear documentation of the presence of immune cells within the infarct following MCAO (Stevens et al. 2002), it does not appear that the majority of these cells are of splenic origin. The types of immune cells found in the brain are consistent with the time course for different immune cell populations to appear within the brain following stroke (Stevens et al. 2002). Following the temporal pattern of immune cell infiltration 48 h post-MCAO, there was the presence of monocytes and a lack of CFSE positive T cells. Additionally, the presence of both T cells and monocytes 96 h post-MCAO is consistent with the infiltration of these cells, as T cells are found later as part of the delayed adaptive immune response (Stevens et al. 2002). The presence of these cells is known to exacerbate delayed neural injury following stroke and removal of the spleen decreases the number of peripheral immune cells that enter the brain (Ajmo et al. 2008; Seifert et al. 2012). Therefore, if splenocytes are not entering the brain parenchyma, but enhancing the pro-inflammatory response to the injured neural tissue, then the cells might be secreting inflammatory mediators. These inflammatory cytokines or chemokines could be the method by which splenocytes are detrimental following stroke versus being directly cytotoxic to neural cells. Secretion of proinflammatory cytokines which activate microglia/macrophages at the site of injury would be deleterious to the survival of neural cells following stroke.
This set of experiments has identified a safe way to track the cellular splenic response in vivo following permanent MCAO in rats by using CFSE, there was no difference in splenocyte numbers between all groups, no effect on infarct size, and no deaths or side effects associated with CFSE injection (data not shown). There is a transient decrease in splenic size following stroke. This decrease in spleen size is associated with an increase in circulating splenocytes which migrate to the brain. These data provide some insight into the mechanism by which the spleen is detrimental in ischemic brain injuries and why splenectomy is neuroprotective in these types of injuries. Further investigation into the splenic reaction in stroke patients would provide insight into how the peripheral immune system can be modulated following stroke to improve neurological outcomes.
References
Ajmo CT Jr, Vernon DO, Collier L, Pennypacker KR, Cuevas J (2006) Sigma receptor activation reduces infarct size at 24 hours after permanent middle cerebral artery occlusion in rats. Curr Neurovasc Res 3(2):89–98
Ajmo CT Jr, Vernon DO, Collier L, Hall AA, Garbuzova-Davis S, Willing A, Pennypacker KR (2008) The spleen contributes to stroke-induced neurodegeneration. J Neurosci Res 86:2227–2234
Ajmo CT Jr., Collier LA, Leonardo CC, Hall AA, Green SM, Womble TA, Cuevas J, Willing AE, Pennypacker KR (2009) Blockade of adrenoreceptors inhibits the splenic response to stroke. Exp Neurol
Bakovic D, Eterovic D, Saratlija-Novakovic Z, Palada I, Valic Z, Bilopavlovic N, Dujic Z (2005) Effect of human splenic contraction on variation in circulating blood cell counts. Clin Exp Pharmacol Physiol 32(11):944–951
Bao Y, Kim E, Bhosle S, Mehta H, Cho S (2010) A role for spleen monocytes in post-ischemic brain inflammation and injury. J Neuroinflammation 7:92
Chang L, Chen Y, Li J, Liu Z, Wang Z, Chen J, Cao W, Xu Y (2011) Cocaine-and amphetamine-regulated transcript modulates peripheral immunity and protects against brain injury in experimental stroke. Brain Behav Immun
Chopp M, Li Y, Jiang N, Zhang RL, Prostak J (1996) Antibodies against adhesion molecules reduce apoptosis after transient middle cerebral artery occlusion in rat brain. J Cereb Blood Flow Metab 16(4):578–584
Das M, Leonardo CC, Rangooni S, Mohapatra SS, Mohapatra S, Pennypacker KR (2011) Lateral fluid percussion injury of the brain induces CCL20 inflammatory chemokine expression in rats. J Neuroinflammation 8:148
Duckworth EA, Butler TL, De Mesquita D, Collier SN, Collier L, Pennypacker KR (2005) Temporary focal ischemia in the mouse: technical aspects and patterns of Fluoro-Jade evident neurodegeneration. Brain Res 1042(1):29–36
Gendron A, Teitelbaum J, Cossette C, Nuara S, Dumont M, Geadah D, du Souich P, Kouassi E (2002) Temporal effects of left versus right middle cerebral artery occlusion on spleen lymphocyte subsets and mitogenic response in Wistar rats. Brain Res 955(1–2):85–97
Hall AA, Guyer AG, Leonardo CC, Ajmo CT Jr, Collier LA, Willing AE, Pennypacker KR (2009) Human umbilical cord blood cells directly suppress ischemic oligodendrocyte cell death. J Neurosci Res 87(2):333–341
Hurn PD, Subramanian S, Parker SM, Afentoulis ME, Kaler LJ, Vandenbark AA, Offner H (2007) T- and B-cell-deficient mice with experimental stroke have reduced lesion size and inflammation. J Cereb Blood Flow Metab 27(11):1798–1805
Jiang H, Meng F, Li W, Tong L, Qiao H, Sun X (2007) Splenectomy ameliorates acute multiple organ damage induced by liver warm ischemia reperfusion in rats. Surgery 141(1):32–40
Kanemoto Y, Nakase H, Akita N, Sakaki T (2002) Effects of anti-intercellular adhesion molecule-1 antibody on reperfusion injury induced by late reperfusion in the rat middle cerebral artery occlusion model. Neurosurgery 51(4):1034–1041, discussion 1041-1032
Karrer FM, Reitz BL, Hao L, Lafferty KJ (1992) Fluorescein labeling of murine hepatocytes for identification after intrahepatic transplantation. Transplant Proc 24(6):2820–2821
Lee ST, Chu K, Jung KH, Kim SJ, Kim DH, Kang KM, Hong NH, Kim JH, Ban JJ, Park HK, Kim SU, Park CG, Lee SK, Kim M, Roh JK (2008) Anti-inflammatory mechanism of intravascular neural stem cell transplantation in haemorrhagic stroke. Brain 131(Pt 3):616–629
Leuschner F, Panizzi P, Chico-Calero I, Lee WW, Ueno T, Cortez-Retamozo V, Waterman P, Gorbatov R, Marinelli B, Iwamoto Y, Chudnovskiy A, Figueiredo JL, Sosnovik DE, Pittet MJ, Swirski FK, Weissleder R, Nahrendorf M (2010) Angiotensin-converting enzyme inhibition prevents the release of monocytes from their splenic reservoir in mice with myocardial infarction. Circ Res 107(11):1364–1373
Li M, Li F, Luo C, Shan Y, Zhang L, Qian Z, Zhu G, Lin J, Feng H (2011) Immediate splenectomy decreases mortality and improves cognitive function of rats after severe traumatic brain injury. J Trauma
Liesz A, Suri-Payer E, Veltkamp C, Doerr H, Sommer C, Rivest S, Giese T, Veltkamp R (2009) Regulatory T cells are key cerebroprotective immunomodulators in acute experimental stroke. Nat Med 15(2):192–199
Liesz A, Zhou W, Mracsko E, Karcher S, Bauer H, Schwarting S, Sun L, Bruder D, Stegemann S, Cerwenka A, Sommer C, Dalpke AH, Veltkamp R (2011) Inhibition of lymphocyte trafficking shields the brain against deleterious neuroinflammation after stroke. Brain 134(Pt 3):704–720
Longa E, Weinstein P, Carlson S, Cummins R (1989) Reversible middle cerebral artery occlusion without craniectomy in rats. Stroke 20:84–91
Lyons AB (2000) Analysing cell division in vivo and in vitro using flow cytometric measurement of CFSE dye dilution. J Immunol Methods 243(1–2):147–154
Meyer S, Strittmatter M, Fischer C, Georg T, Schmitz B (2004) Lateralization in autonomic dysfunction in ischemic stroke involving the insular cortex. Neuroreport 15(2):357–361
Newcomb JD, Ajmo CT, Sanberg CD, Sanberg PR, Pennypacker KR, Willing AE (2006) Timing of cord blood treatment after experimental stroke determine therapeutic efficacy. Cell Transplant 15(3):213–223
Offner H, Subramanian S, Parker SM, Afentoulis ME, Vandenbark AA, Hurn PD (2006a) Experimental stroke induces massive, rapid activation of the peripheral immune system. J Cereb Blood Flow Metab 26(5):654–665
Offner H, Subramanian S, Parker SM, Wang C, Afentoulis ME, Lewis A, Vandenbark AA, Hurn PD (2006b) Splenic atrophy in experimental stroke is accompanied by increased regulatory T cells and circulating macrophages. J Immunol 176(11):6523–6531
Okuaki Y, Miyazaki H, Zeniya M, Ishikawa T, Ohkawa Y, Tsuno S, Sakaguchi M, Hara M, Takahashi H, Toda G (1996) Splenectomy-reduced hepatic injury induced by ischemia/reperfusion in the rat. Liver 16(3):188–194
Savas MC, Ozguner M, Ozguner IF, Delibas N (2003) Splenectomy attenuates intestinal ischemia-reperfusion-induced acute lung injury. J Pediatr Surg 38(10):1465–1470
Savitz SI, Erhardt JA, Anthony JV, Gupta G, Li X, Barone FC, Rosenbaum DM (2000) The novel beta-blocker, carvedilol, provides neuroprotection in transient focal stroke. J Cereb Blood Flow Metab 20(8):1197–1204
Schmued LC, Albertson C, Slikker W Jr (1997) Fluoro-Jade: a novel fluorochrome for the sensitive and reliable histochemical localization of neuronal degeneration. Brain Res 751(1):37–46
Schroeter M, Jander S, Witte OW, Stoll G (1994) Local immune responses in the rat cerebral cortex after middle cerebral artery occlusion. J Neuroimmunol 55(2):195–203
Schwarting S, Litwak S, Hao W, Bahr M, Weise J, Neumann H (2008) Hematopoietic stem cells reduce postischemic inflammation and ameliorate ischemic brain injury. Stroke 39(10):2867–2875
Seifert HA, Leonardo CC, Hall AA, Rowe DD, Collier LA, Benkovic SA, Willing AE, Pennypacker KR (2012) The spleen contributes to stroke induced neurodegeneration through interferon gamma signaling. Metab Brain Dis 27(2):131–141
Stevens SL, Bao J, Hollis J, Lessov NS, Clark WM, Stenzel-Poore MP (2002) The use of flow cytometry to evaluate temporal changes in inflammatory cells following focal cerebral ischemia in mice. Brain Res 932(1–2):110–119
Swirski FK, Nahrendorf M, Etzrodt M, Wildgruber M, Cortez-Retamozo V, Panizzi P, Figueiredo JL, Kohler RH, Chudnovskiy A, Waterman P, Aikawa E, Mempel TR, Libby P, Weissleder R, Pittet MJ (2009) Identification of splenic reservoir monocytes and their deployment to inflammatory sites. Science 325(5940):612–616
Vendrame M, Gemma C, Pennypacker KR, Bickford PC, Davis Sanberg C, Sanberg PR, Willing AE (2006) Cord blood rescues stroke-induced changes in splenocyte phenotype and function. Exp Neurol 199(1):191–200
Weston SA, Parish CR (1990) New fluorescent dyes for lymphocyte migration studies. Analysis by flow cytometry and fluorescence microscopy. J Immunol Methods 133(1):87–97
Acknowledgements
The authors would like to thank the Lisa Muma Weitz Laboratory for Advanced Microscopy & Cell Imaging for their assistance with acquiring the confocal microscopy images. This study was funded by the National Institutes of Health grant RO1 NS052839.
Conflict of interest
The authors have no conflict of interest.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article
Cite this article
Seifert, H.A., Hall, A.A., Chapman, C.B. et al. A Transient Decrease in Spleen Size Following Stroke Corresponds to Splenocyte Release into Systemic Circulation. J Neuroimmune Pharmacol 7, 1017–1024 (2012). https://doi.org/10.1007/s11481-012-9406-8
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s11481-012-9406-8