
Abstract
Purpose
Fe(III) transformation to Fe(II) via the nitrate-dependent iron oxidation process, occurring in anoxic sediments, has an important role in the nitrogen cycle. In this study, laboratory experiments were performed to investigate the denitrification properties driven by nitrate-dependent iron oxidation.
Materials and methods
A 30-day incubation study of sediments and overlying water from Lake Moshui in Wuhan, China, was conducted in an anoxic condition. The nitrate reduction during the incubation was evaluated by the N2O emission and various forms of nitrogen in the overlying water. The denitrification enzyme activity and abundance of nitrate-dependent Fe(II)-oxidising bacteria were determined periodically, and their correlations with nitrogen and iron were analysed to illustrate the denitrification characteristics linked to Fe(II) oxidation in sediments.
Results and discussion
After the Fe(II) and nitrate input, the decrease of Fe(II) and nitrate concentrations was accompanied by an increase in nitrite and N2O production. The contribution of Fe(II) oxidation to the nitrate reduction accounted for 27.7% at the end of the incubation, and the rate of Fe(II) decrease was significantly correlated (P < 0.05) with the production of N2O. In addition, a positive correlation between denitrification enzyme activity and nitrate concentrations was observed. During incubation, the abundance of nitrate-dependent Fe(II)-oxidising bacteria in the sediment ranged from 1.1 × 105 cell g−1 wet sediment to 1.4 × 106 cell g−1 wet sediment, and increased with the increase of Fe(II) input concentration. The nitrate reduction coupled with Fe(II) oxidation was mainly mediated by microbial processes.
Conclusions
Sediment denitrification was enhanced with increasing Fe(II) concentrations, and Fe(II) may play an important role in regulating nitrogen transformation in freshwater lakes.
Similar content being viewed by others

Explore related subjects
Discover the latest articles, news and stories from top researchers in related subjects.Avoid common mistakes on your manuscript.
1 Introduction
Denitrification, a microbial process by which nitrate is reduced stepwise into atmospheric terminal products such as N2O and/or N2 (Lam and Kuypers 2011; Silva et al. 2018), has ecological and geochemical consequences in freshwater systems and wastewater treatment plants (Dunne et al. 2013; Nizzoli et al. 2018; Zhang et al. 2019). Denitrification mostly occurs in the sediment and acts as the prevailing sink for nitrogen in the water column or sediments under anaerobic conditions. Denitrification plays an important role in controlling nitrogen transformation, accounting for more than 70% of natural nitrogen loss (Babbin et al. 2014; Gao et al. 2016).
Organic carbon is a common electron donor for denitrification (Kesserű et al. 2002; Wunderlich et al. 2012; Wu et al. 2013), whereas inorganic substrates, including reduced sulphur (Yang et al. 2012) and iron compounds (Melton et al. 2014), can support denitrification as well. Sulphide oxidation coupled with nitrate reduction has been intensively studied (Jørgensen et al. 2009) and identified as an important pathway of nitrogen cycling in anoxic water (Canfield et al. 2010; Wenk et al. 2013) and potentially in sediments (Sayama et al. 2005; Burgin and Hamilton 2007). However, the possibility of Fe(II) as an electron donor for nitrate reduction in the natural environment had not been investigated until recent decades (Robertson and Thamdrup 2017).
Iron is an abundant redox-active element in sediments, and the transition between Fe(II) and Fe(III) plays an important role on watershed biogeochemistry (Weber et al. 2006a; Melton et al. 2014). Iron can be utilised as an electron donor by many various microorganisms that gain energy from that reaction, and the iron cycle is associated with many other elementary cycles, such as the nitrogen cycle (Laufer et al. 2016). Microbial and partly abiotic mechanisms by which Fe(II) is oxidised with nitrate are summarised as “nitrate-dependent iron oxidation” (Scholz et al. 2016). Water-soluble Fe(II) is generally less reactive, but it is readily utilised by microorganisms as an electron donor (Straub et al. 1996). These microorganisms include aerobic acidophilic (Bonnefoy and Holmes 2012), micro-aerobic neutrophilic (Krepski et al. 2012), and anaerobic bacteria using light (Hegler et al. 2008) or nitrate-dependent Fe(II)-oxidising bacteria (NDFOB) (Straub et al. 1996), which have been found in many different environments including soils, freshwater sediments, marine sediment, and hypersaline sediments (Shelobolina et al. 2012; Rowe et al. 2014; Laufer et al. 2015). Most NDFOB can reduce nitrate to nitrite, followed by further reduction to NO, N2O, and N2 (Chakraborty and Picardal 2013; Melton et al. 2014). The identification of NDFOB strains and investigation of functional enzymes involved in nitrate-dependent iron oxidation could be performed using molecular sequencing and transcription techniques (Beller et al. 2013; Dubinina and Sorokina 2014; Li et al. 2018; Hu et al. 2019). In addition, the critical metabolic components of NDFOB could be identified by whole-genome sequencing (Hu et al. 2017).
Various enzymes involved in the main reaction phase of nitrogen cycling have been discovered in lake sediments, and some scholars have conducted in-depth research on denitrifying enzymes (Cao et al. 2008; Gardner and White 2010). The conversion process of nitrogen compounds to nitrate can be carried out not only in microbial cells but also under enzymatic reactions. A specific enzyme, especially in anaerobic conditions, was involved in each stage of the nitrate transformation process. In contrast to our knowledge of NDFOB, it is largely unknown whether the reaction concerning nitrate reduction coupled with Fe(II) oxidation is related to denitrification enzyme activity (DEA) responsible for the process of nitrate reduction.
With the increase in nitrogen input to a lake, the likelihood of eutrophication increases, resulting in more frequent anoxic conditions in the lake. Hence, more transformation of Fe(III) to Fe(II) occurs, and the role of nitrate-dependent iron oxidation becomes more important. However, an understanding of the internal relationship between nitrogen transformation and nitrate-dependent iron oxidation under increasing nitrate and Fe(II) is still lacking. In this study, a 30-day incubation experiment with the sediments and overlying water from a freshwater lake was carried out. The nitrogen cycle, including water and gas, and its response to Fe(II) oxidation were investigated. We also assessed the possible effect of nitrate-dependent Fe(II) oxidation on denitrification enzyme activity and NDFOB numbers in sediments. Our aim was to better understand the connection between Fe(II) oxidation and the nitrogen cycle through denitrification.
2 Materials and methods
2.1 Sampling
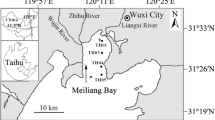
Lake Moshui, a freshwater lake with an area of 3.14 km2 and a mean depth of approximately 1.2 m, is located in Wuhan, Hubei Province, China. The sediments and overlying water of Lake Moshui were taken from eight sites on April 9, 2018 (Fig. 1). The sediments at a depth of 10–15 cm were collected using a Petersen grab dredge and stored in a bag, and 10 L overlying water at each site was stored in a bucket. The pH, EC, dissolved oxygen (DO), and water temperature of the overlying water were measured in situ using a water quality analyser (EXO3, YSI international Co., Ltd., USA). In addition, several 200-mL glass bottles pre-filled with 4 mL hydrochloric acid solution were filled with overlying water and sealed with a rubber stopper for Fe(II) and Fe(III) analysis. The parameter properties in the overlying water and sediments are listed in Tables 1 and 2. All samples were stored at 4 °C before the incubation experiments.

Location of the sampling sites in Lake Moshui (site 1, 30°33′06.4″N 114°12′45.7″E; site 2, 30°32′44.9″N 114°12′44.0″E; site 3, 30°32′32.1″N 114°13′08.4″E; site 4, 30°32′44.4″N 114°13′09.2″E; site 5, 30°32′46.6″N 114°13′30.1″E; site 6, 30°32′38.9″N 114°13′50.9″E; site 7, 30°32′22.7″N 114°14′05.6″E; site 8, 30°32′09.6″N 114°13′38.3″E)
2.2 Sediment slurry incubations
The sediments and overlying water were prepared for the incubation experiment by mixing an equal amount of sample from the eight sites. Then, 100 g of fresh sediments and 400 mL of overlying water were added to 500-mL plastic bottles. One hundred eight bottles were prepared and split into three groups: a control group (CK), with original sediments and overlying water, and two treatment groups, to which NaNO3 was added to obtain a concentration of 32.5 μM nitrate (N group) and NaNO3+FeSO4·7H2O was added simultaneously to obtain the concentrations of 32.5 μM nitrate and 71.25 μM Fe(II) (N+F group). All incubation bottles were then sealed with an inner lid and outer lid to achieve anoxic conditions. Two needles fitted with a three-way valve were inserted into the inner lid for gas and solution sampling. The three groups were then placed on a shaking table (50 r min−1) at 25 °C for 10 min, followed by incubation in a dark incubator at 25 °C.
At each sampling time, 0, 0.5, 1, 3, 5, 7, 10, 14, 18, 22, 26, and 30 days, the incubation bottles were removed from the three groups. Three replicates were used in each group. The gas was sampled before the determination of other parameters. The pH and DO were measured in situ. Similarly, the sediment samples were freeze-dried (− 80 °C) for the following determinations.
An additional experiment was conducted to justify that the dominant reaction was biotic or abiotic. Sterilised (121 °C for 20 min) and unsterilised samples of the sediments and overlying water taken from Lake Moshui were prepared. The subsequent procedure was as described above for the N+F group, and the samples were prepared in triplicate. Gas for N2O flux determination after 1 day and water for nitrate determination after 18 days were taken with the aforementioned syringes.
2.3 Analysis and calculation
Overlying water was filtered (0.45 μm) for the measurements of nitrate, nitrite, and ammonium with standard photometric techniques (State Environmental Protection Administration of the People’s Republic of China 2002), and total nitrogen (TN) concentrations were measured using the oxidation method of alkaline potassium persulfate (Valderrama 1981). Dissolved iron in the overlying water and sediments was measured photometrically using the Ferrozine assay (Viollier et al. 2000). The pH and DO concentrations of the overlying water were determined using a water quality analyser (EXO3, YSI international Co., Ltd., USA). N2O and CO2 concentrations were analysed by a gas chromatograph (7890A, Agilent technologies Co., Ltd., USA) equipped with an electron capture detector. The concentrations of N2O and CO2 in the headspace gas were determined, and the N2O and CO2 fluxes were calculated using the following Eq. 1 (Hayakawa et al. 2013) and Eq. 2 (Wang et al. 2016), respectively.
where FN2O and FCO2 are the N2O and CO2 fluxes (μg N kg−1 day−1), respectively, M is the dry weight of the sediment (kg), Δt is the incubation time (day), ρ is the density of N2O and CO2 under standard temperature and pressure (1.25 g L−1 and 1.977 g L−1, respectively), C is the concentration of N2O and CO2 (ppmv), Vg is the headspace volume of the incubation bottle (L), α is the Bunsen coefficient of N2O (0.549 at 25 °C) (Tiedje 1994), VL is the liquid volume (L), 273 is the conversion factor between degrees Celsius and kelvin, T is the temperature (°C), and 12/44 is the conversion factor from CO2 to C.
The C2H2 inhibition method (Smith and Tiedje 1979) was used to determine the DEA of the sediment. NDFOB inoculation was conducted at the UV sterilisation station. The sediment samples were diluted to five gradients, 10−3, 10−4, 10−5, 10−6, and 10−7, and each dilution had five replicate series. Following the preparation of a serially diluted suspension, vitamins and trace metals (10 mL L−1 each) from stock solutions (Bruce et al. 1999) were added into the incubation tubes containing 1 mL serially diluted suspension. Sodium pyrophosphate (0.1%) was added to the incubation tubes sterilised prior to use. The incubation tubes were then filled with three different bicarbonate-buffered freshwater basal media (Weber et al. 2006c). For incubation of dilution, all incubation tubes were closed with butyl rubber stoppers to achieve anoxic conditions and incubated in a dark biochemical incubator at 25 °C. After 6 weeks of incubation, the growth was detected by the presence of a brownish-red or brownish-green precipitate in the incubation tubes (Weber et al. 2006c). The NDFOB in the incubation tubes were counted according to the MPN method (Cochran 1950).
2.4 Statistical analyses
The correlation between parameters was analysed by Pearson correlation analysis. F statistics were generated in SPSS 20.0 to evaluate the relationship of Fe(II) oxidation and N2O flux. Data from the sediment slurry incubation experiment were analysed for significant differences in N2O cumulative emission of representative days by comparison using a t test in SPSS 20.0.
3 Results
3.1 DO, pH, and nitrogen in the overlying water
The DO concentration in the three groups showed a sharp decrease after 1 day of incubation, followed by a plateau range of 0.6–1.0 mg L−1 (Fig. 2). A decrease during the early days was observed for the pH of all the groups, but little change in pH was found after 10 days of incubation. In general, the pH of the N and the N+F groups showed lower levels than that of the CK group, indicating that more H+ was produced after nitrate input during the incubation.

DO concentrations and pH in the overlying water during the incubation
No sharp decrease in nitrate concentrations was observed in the overlying water of the CK group, whereas an obvious decrease in nitrate concentrations occurred in the N group and N+F group mainly due to denitrification (Fig. 3a). The reduction rate of nitrate in the N group (0.071 day−1, R2 = 0.9300) and N+F group (0.089 day−1, R2 = 0.9797) followed the pseudo-first-order reaction in the first-week incubation, consistent with a previous study by Pyzola (2013). After the 30-day incubation, the nitrate removal efficiencies in the N group and N+F group were 75.7% and 81.9%, respectively.

Nitrogen concentrations in the overlying water during the incubation. NO3--N (a), NO2--N (b), NH4+-N (c) and TN (d)
Contrary to the nitrate decrease, the nitrite concentrations in the N and N+F groups illustrated a rising profile before 22 days (Fig. 3b). Nitrite concentrations for CK remained relatively stable during the incubation. Ammonium concentrations continued to be released from the sediment to the overlying water (Fig. 3c), probably attributed to the ammonification by the microorganisms in the sediments. Ammonium concentrations were higher in the CK group than those in the other two groups from 7 to 30 days. Drastic decreases in TN concentrations after 1 day of incubation were observed in all the samples (Fig. 3d), consistent with the decrease in DO. Despite the initial nitrate input, TN concentrations in the N group and the N+F group were lower than those in the CK group in most cases, suggesting much more rapid denitrification in the sediments.
3.2 Fe(II) and Fe(III) concentrations in the overlying water
The oxidation rate of Fe(II) followed the pseudo-first-order reaction in the N group and the N+F group in the first 5-day incubation (Fig. 4), and the rates were 0.064 day−1 (R2 = 0.8892) and 0.079 day−1 (R2 = 0.9670), respectively. No significant change in Fe(II) concentration was observed for the CK group (P > 0.05). After the 30-day incubation, the decrease in the percentages of Fe(II) concentration in the N+F group (47%) and the N group (43%) was similar but higher than that in the CK group (23%). As the initial Fe(II) concentration in the N+F group was the same as that in the CK group, a lower concentration at the end of incubation meant that more rapid transformation of Fe(II) occurred in the N+F group.

Fe(II) (a) and Fe(III) (b) concentrations in the overlying water during the incubation
A drastic increase in Fe(III) in all groups was observed after a 1-day incubation, resulting from Fe(II) transformation. For the N+F group, the 30-day decrease in Fe(II) concentration was approximately 57 μmol L−1, whereas only a 42 μmol L−1 increase in Fe(III) concentration was found. The increase in Fe(III) concentration was not equal to the decrease in Fe(II) oxidation, probably attributed to the production of poorly soluble Fe(III) minerals.
3.3 N2O emissions
The anoxic conditions promoted denitrification, leading to a peak of N2O fluxes in all groups after 1 day (Fig. 5). A slow decline was illustrated during the latter incubation period. The N2O fluxes were significantly higher in the N and N+F groups than those in the CK group (P < 0.05), indicating that N2O emissions were promoted with nitrate addition. The N2O fluxes in the N and N+F groups ranged from 15.7 to 23.5 μg N kg−1 day−1 after 21 days and were higher than those in the CK group (5.4–7.3 μg N kg−1 day−1). In addition, the N2O emission in the N+F group was higher than that in the N group, and the difference could be mediated by the Fe(II) addition. During the whole incubation period, no significant differences in CO2 fluxes among the N, N+F, and CK groups (P > 0.05) were observed.

Variations in N2O (a) and CO2 (b) fluxes in the overlying water during the incubation
3.4 DEA and NDFOB numbers in the sediment
The denitrification in the sediments was accelerated as the DO decreased drastically during 1 day of incubation, which could be reflected by the DEA (Fig. 6). Similar to the N2O emission, the DEA in the three groups showed a high level after 1 day, indicating an internal connection between N2O production and DEA, and then fluctuated between 6 and 9 mg N kg−1 h−1. Although no significant differences (P > 0.05) in DEA were observed among the three groups, the DEA of the N+F group was generally higher than that of the CK group and the N group.

DEA and NDFOB numbers in the sediments during the incubation
The NDFOB in the sediments was approximately 1 × 105 cells g−1 wet sediment for all the three groups at the beginning of the incubation (Fig. 6). Comparing cell numbers in the three groups, we observed a similar development of cell numbers within the first 3 days: the exponential growth phase appeared shortly after incubation, and all samples reached a cell number of 9.0 × 105 cells g−1 wet sediment. After 7 days, the growth of NDFOB in the N and N+F groups was faster than that in the CK group. The cell numbers in the three groups were then kept stable after 10 days. The cell numbers of NDFOB at the end of incubation in the CK group, N group, and N+F group were 9.5 × 105, 1.1 × 106, and 1.4 × 106 cell g−1 wet sediment, respectively.
4 Discussion
4.1 Transformation of nitrogen coupled with iron oxidation in the overlying water
Nitrate coupled with Fe(II) oxidation results in the production of nitrite, N2O, N2, or ammonium, related to the physicochemical properties of water and the authigenic Fe minerals (Weber et al. 2006b; Scholz et al. 2016; Robertson and Thamdrup 2017). Compared with the N group, the N+F group showed higher nitrate reduction and nitrite production in the overlying water. This finding could be attributed to the following reaction (Scholz et al. 2016):
Based on the pH, nitrate, nitrite, and Fe(II) concentrations of the incubation experiment, the Gibbs free energy was calculated, ΔG = −98.8 kJ/mol, indicating that the oxidation of Fe(II) was favourable in the anoxic sediment slurry incubation and provided powerful support for the continued increase in nitrite concentration in the N and the N+F groups. Lower concentrations of ammonium in the N and N+F groups than in the CK group were probably caused by the anaerobic ammonium oxidation (anammox), as an increase in nitrite production appeared in the two groups. However, anammox, one of the pathways for nitrogen removal in the watershed (Yu et al. 2016), usually played a minor role compared to the denitrification process (Schubert et al. 2006; Han et al. 2014). There was a significant relation between TN and nitrite (R = − 0.791, P < 0.01). An apparent decline in total nitrogen in the N and N+F groups (Fig. 7) meant that a reducing gas such as N2O and N2 was formed, by which nitrogen left the aqueous phase for the gas phase.

Values of NDFOB and DEA in relation to the concentrations of nitrate, nitrite, ammonium, TN, Fe(II), Fe(III), and the cumulative emission of N2O during the incubation. Numbers after three groups represent different incubation days
The consumption of Fe(II) in the N+F group was high during the incubation, especially in the first 3 days, at approximately 8 μmol L−1 day−1. Fe(II) oxidation with oxygen, either abiotically or microbially induced, is a viable explanation for rapid Fe(II) consumption (Scholz et al. 2016). The increased Fe(III) concentration in the overlying water was not equal to the decreased Fe(II) concentration, probably because both soluble and insoluble Fe(II) were oxidised by bacterium to produce various iron minerals (Chaudhuri et al. 2001; Weber et al. 2006a). In the anoxic water, most iron is rapidly re-precipitated (Scholz et al. 2016). There was a negative relation between Fe(III) and TN (R = − 0.589, P < 0.01), whereas a positive correlation between Fe(III) and nitrite (R = 0.549, P < 0.01) was found. Nitrogen transformation and iron cycle are potentially coupled in anoxic environments (Kampschreur et al. 2011), and a positive relation was observed between the Fe(II) and nitrate concentrations (R = 0.483, P < 0.05). The reaction rates of Fe(II) and nitrate were used to establish the slopes of the linear least squares fit of Fe(II) and nitrate, thus estimating the contribution of Fe(II) oxidation to nitrate reduction via the following equation (Eq. 4):
The contribution of Fe(II) oxidation to the reduction of nitrate was estimated to be 27.7% according to the method by Pyzola (2013).
4.2 N2O production driven by iron oxidation
As nitrite accumulation was known to greatly increase N2O emission, N2O fluxes in the N and N+F groups during the incubation were higher than those in the CK group. The sharp increase in N2O emission after 1 day indicated that denitrification occurred rapidly in the anoxic conditions (Hibiya et al. 2003). Compared with the cumulative N2O emissions in the CK group, those in the N group and the N+F group at the end of incubation increased by 157% and 231%, respectively, indicating that nitrate reduction may proceed in a required environment with relatively high concentrations of nitrate related to the high rate of denitrification. A significant positive correlation (R = 0.757, P < 0.01, t test) between N2O fluxes and nitrate concentrations indicated that denitrification was most likely the main source of N2O.
The N2O fluxes were positively correlated with Fe(II) concentrations during incubation (R = 0.560, P < 0.01). N2O cumulative emission in the N+F group was generally higher than that in the N group, indicating that denitrification was stimulated with Fe(II) addition. Organic carbon is often used as the main electron donor for denitrification (Cayuela et al. 2014); however, Fe(II) can also support denitrification by abiotic and biotic pathways (Melton et al. 2014). Hence, the aspect of environmentally unwanted N2O production enhanced by this pathway should be realised when removing nitrogen from nitrogen-rich and iron-rich waters, especially in anoxic conditions (Kampschreur et al. 2011).
4.3 Microbial properties motivated by nitrate-dependent iron oxidation
The DEA and NAFOB numbers in sediments were important for the denitrification as the sediments were beneficial to nitrogen removal in watershed (Dai et al. 2018). The pH in the overlying water ranged from 7.2 to 7.8 during the incubation, providing a favourable environment for DEA. A previous study (Šimek et al. 2002) showed that the optimum pH for denitrifying bacteria growth and DEA is neutral or slightly alkaline. The DEA in the N+F group was higher than that in the CK group, indicating a high denitrification rate mediated by the Fe(II) addition. The DEA showed an apparent decline in all groups after a week incubation. This result was due to the nitrate-consuming reactions, and the lack of nitrate slowed the bacterial denitrification rates. A positive correlation between DEA and N2O emission (R = 0.711, P < 0.01) but a negative correlation between DEA and NDFOB (R = − 0.566, P < 0.01) was observed during the incubation (Fig. 7). Hence, the results did not indicate with certainty that the nitrate reduction coupled with Fe(II) oxidation was catalysed by DEA.
A significant relation between NAFOB and nitrite was found (R = 0.769, P < 0.01), and the relation between NAFOB and Fe(III) was also significant (R = 0.654, P < 0.01). The bacteria in the N group multiplied better than those in the CK group, indicating that NAFOB growth consumed nitrate as the nitrogen source (Straub et al. 1996; Straub and Buchholz-Cleven 1998). The NAFOB numbers in the N+F group were generally larger than those in the CK group, and the N group during incubation as NDFOB gained more energy with the additional Fe(II), and the activity of nitrate reduction coupled to Fe(II) oxidation was promoted to some extent. A previous study (Weber et al. 2009) has shown that Fe(II) acts as a “second substrate” that provides energy for microbial growth and that abundant energy is beneficial to NAFOB growth. A positive correlation between NDFOB numbers and N2O cumulative emission was observed (Fig. 7), illustrating that the nitrate reduction coupled with Fe(II) oxidation was driven by bacteria. NDFOB numbers ranging from 1 × 103 to 5 × 108 cell g−1 sediment (Hauck et al. 2001; Senn and Hemond 2002; Weber et al. 2006b) were reported, consistent with the number of NDFOB (1.1 × 105–1.4 × 106 cell g−1 wet sediment) observed in the sediments during the current incubation.
The nitrate concentration in the sterilised sediment was significantly higher than that in the unsterilised sediment on the 18th day of incubation (P < 0.01, t test) (Fig. 8). Furthermore, there was a significant difference (P < 0.01, t test) in the N2O fluxes and nitrate concentrations between sterilised and unsterilised sediment incubation (Fig. 8). A N2O flux of 55.4 μg N kg−1 day−1 was detected in the sterilised sediment slurry incubation, whereas an abundant N2O flux of 362.9 μg N kg−1 day−1 was observed in the unsterilised sediment after 1 day of incubation. Abiotic denitrification driven by Fe(II) can also occur in sediments, especially in the condition of nitrite accumulation (Kampschreur et al. 2011), and the chemical denitrification of nitrite mediated by Fe(II) oxidation can lead to the formation of N2O (Kampschreur et al. 2011). The sterilised experiment indicated that the abiotic pathway accounted for approximately 15% of the N2O production, and the biotic pathway played a major role in N2O production, i.e., denitrification.

Comparison of N2O (a) concentrations and nitrate (b) fluxes between sterilised and untreated samples of the sediment from Moshui Lake. The bars are labelled with the significant differences between the groups (P < 0.05, t test)
5 Conclusions
The nitrate reduction was accelerated after Fe(II) addition, attributed to Fe(II) used as the second electron donor for denitrification. The contribution of Fe(II) oxidation to the nitrate reduction was 27.7% at the end of the incubation. The Fe(II) and nitrate addition led to an increase in N2O production, especially after 1 day of incubation. Sterilised experiments suggested that nitrate reduction and its subsequent N2O reduction were affected by the combination of biotic and abiotic reactions, and the biotic reaction was dominant. The numbers of NDFOB in the sediment during incubation were 1.1 × 105–1.4 × 106 cell g−1 wet sediment. The current study illustrated that anoxic nitrate-dependent iron oxidation played a non-negligible role in regulating nitrogen transformations, especially in iron-rich freshwater lake sediments.
References
Babbin AR, Keil RG, Devol AH, Ward BB (2014) Organic matter stoichiometry, flux, and oxygen control nitrogen loss in the ocean. Science 344:406–408
Beller HR, Zhou P, Legler TC, Chakicherla A, Kane S, Letain TE, O’Day PA (2013) Genome-enabled studies of anaerobic, nitrate-dependent iron oxidation in the chemolithoautotrophic bacterium Thiobacillus denitrificans. Front Microbiol 4:249
Bonnefoy V, Holmes DS (2012) Genomic insights into microbial iron oxidation and iron uptake strategies in extremely acidic environments. Environ Microbiol 14:1597–1611
Bruce RA, Achenbach LA, Coates JD (1999) Reduction of (per)chlorate by a novel organism isolated from paper mill waste. Environ Microbiol 1:319–329
Burgin AJ, Hamilton SK (2007) Have we overemphasized the role of denitrification in aquatic ecosystems? A review of nitrate removal pathways. Front Ecol Environ 5:89–96
Canfield DE, Stewart FJ, Thamdrup B, De Brabandere L, Dalsgaard T, Delong EF, Revsbech NP, Ulloa O (2010) A cryptic sulfur cycle in oxygen-minimum-zone waters off the Chilean coast. Science 330:1375–1378
Cao YP, Green PG, Holden PA (2008) Microbial community composition and denitrifying enzyme activities in salt marsh sediments. Appl Environ Microbiol 74:7585–7595
Cayuela ML, van Zwieten L, Singh BP, Jeffery S, Roig A, Sánchez-Monedero MA (2014) Biochar’s role in mitigating soil nitrous oxide emissions: a review and meta-analysis. Agric Ecosyst Environ 191:5–16
Chakraborty A, Picardal F (2013) Induction of nitrate-dependent Fe(II) oxidation by Fe(II) in Dechloromonas sp. strain UWNR4 and Acidovorax sp. strain 2AN. Appl Environ Microbiol 79:748–752
Chaudhuri SK, Lack JG, Coates JD (2001) Biogenic magnetite formation through anaerobic biooxidation of Fe(II). Appl Environ Microbiol 67:2844–2848
Cochran WG (1950) Estimation of bacterial densities by means of the “most probable number”. Biometrics 6:105–116
Dai JW, He SB, Zhou WL, Huang JC, Chen S, Zeng XH (2018) Integrated ecological floating bed treating wastewater treatment plant. Environ Sci Pollut Res 25:18793–18801
Dubinina GA, Sorokina AY (2014) Neutrophilic lithotrophic iron-oxidizing prokaryotes and their role in the biogeochemical processes of the iron cycle. Microbiology 83:1–14
Dunne EJ, Coveney MF, Marzolf ER, Hoge VR, Conrow R, Naleway R, Lowe EF, Battoe LE, Inglett PW (2013) Nitrogen dynamics of a large-scale constructed wetland used to remove excess nitrogen from eutrophic lake water. Ecol Eng 61:224–234
Gao J, Hou LJ, Zheng YL, Liu M, Yin GY, Li XF, Lin XB, Yu CD, Wang R, Jiang XF, Sun XR (2016) nirS-Encoding denitrifier community composition, distribution, and abundance along the coastal wetlands of China. Appl Microbiol Biotechnol 100:8573–8582
Gardner LM, White JR (2010) Denitrification enzyme activity as an indicator of nitrate movement through a diversion wetland. Soil Sci Soc Am J 74:1037–1047
Han HJ, Lu XX, Burger DF, Joshi UM, Zhang L (2014) Nitrogen dynamics at the sediment–water interface in a tropical reservoir. Ecol Eng 73:146–153
Hauck S, Benz M, Brune A, Schink B (2001) Ferrous iron oxidation by denitrifying bacteria in profundal sediments of a deep lake (Lake Constance). FEMS Microbiol Ecol 37:127–134
Hayakawa A, Hatakeyama M, Asano R, Ishikawa Y, Hidaka S (2013) Nitrate reduction coupled with pyrite oxidation in the surface sediments of a sulfide-rich ecosystem. J Geophys Res-Biogeo 118:639–649
Hegler F, Posth NR, Jiang J, Kappler A (2008) Physiology of phototrophic iron(II)-oxidizing bacteria: implications for modern and ancient environments. FEMS Microbiol Ecol 66:250–260
Hibiya K, Terada A, Tsuneda S, Hirata A (2003) Simultaneous nitrification and denitrification by controlling vertical and horizontal microenvironment in a membrane-aerated biofilm reactor. J Biotechnol 100:23–32
Hu M, Chen PC, Sun WM, Li FB, Cui JH (2017) A novel organotrophic nitrate-reducing Fe(II)-oxidizing bacterium isolated from paddy soil and draft genome sequencing indicate its metabolic versatility. RSC Adv 7:56611–56620
Hu M, Chen PC, Sun WM, Li FB, Cui JH (2019) Physiological and genomic characterization of a nitrate-reducing Fe(II)-oxidizing bacterium isolated from paddy soil. Geomicrobiol J 36:433–442
Jørgensen CJ, Jacobsen OS, Elberling B, Aamand J (2009) Microbial oxidation of pyrite coupled to nitrate reduction in anoxic groundwater sediment. Environ Sci Technol 43:4851–4857
Kampschreur MJ, Kleerebezem R, de Vet WWJM, van Loosdrecht MCM (2011) Reduced iron induced nitric oxide and nitrous oxide emission. Water Res 45:5945–5952
Kesserű P, Kiss I, Bihari Z, Polyák B (2002) Investigation of the denitrification activity of immobilized Pseudomonas butanovora cells in the presence of different organic substrates. Water Res 36:1565–1571
Krepski ST, Hanson TE, Chan CS (2012) Isolation and characterization of a novel biomineral stalk-forming iron-oxidizing bacterium from a circumneutral groundwater seep. Environ Microbiol 14:1671–1680
Lam P, Kuypers MMM (2011) Microbial nitrogen cycling processes in oxygen minimum zones. Annu Rev Mar Sci 3:317–345
Laufer K, Nordhoff M, Røy H, Schmidt C, Behrens S, Jørgensen BB, Kappler A (2015) Coexistence of microaerophilic, nitrate-reducing, and phototrophic Fe(II) oxidizers and Fe(III) reducers in coastal marine sediment. Appl Environ Microbiol 82:1433–1447
Laufer K, Byrne JM, Glombitza C, Schmidt C, Jorgensen BB, Kappler A (2016) Anaerobic microbial Fe(II) oxidation and Fe(III) reduction in coastal marine sediments controlled by organic carbon content. Environ Microbiol 18:3159–3174
Li S, Li XM, Li FB (2018) Fe(II) oxidation and nitrate reduction by a denitrifying bacterium, Pseudomonas stutzeri LS-2, isolated from paddy soil. J Soils Sediments 18:1668–1678
Melton ED, Swanner ED, Behrens S, Schmidt C, Kappler A (2014) The interplay of microbially mediated and abiotic reactions in the biogeochemical Fe cycle. Nat Rev Microbiol 12:797–808
Nizzoli D, Bartoli M, Azzoni R, Longhi D, Castaldelli G, Viaroli P (2018) Denitrification in a meromictic lake and its relevance to nitrogen flows within a moderately impacted forested catchment. Biogeochemistry 137:143–161
Pyzola S (2013) Nitrate reduction coupled to iron(II) oxidation and manganese(II) in an agricultural soil. Dissertation. University of Kentucky
Robertson EK, Thamdrup B (2017) The fate of nitrogen is linked to iron(II) availability in a freshwater lake sediment. Geochim Cosmochim Acta 205:84–99
Rowe AR, Chellamuthu P, Lam B, Okamoto A, Nealson KH (2014) Marine sediments microbes capable of electrode oxidation as a surrogate for lithotrophic insoluble substrate metabolism. Front Microbiol 5:784
Sayama M, Risgaard-Petersen N, Nielsen LP, Fossing H, Christensen PB (2005) Impact of bacterial NO3− transport on sediment biogeochemistry. Appl Environ Microbiol 71:7575–7577
Scholz F, Löscher CR, Fiskal A, Sommer S, Hensen C, Lomnitz U, Wuttig K, Göttlicher J, Kossel E, Steininger R, Canfield DE (2016) Nitrate-dependent iron oxidation limits iron transport in anoxic ocean regions. Earth Planet Sci Lett 454:272–281
Schubert CJ, Durisch-Kaiser E, Wehrli B, Thamdrup B, Lam P, Kuypers MMM (2006) Anaerobic ammonium oxidation in a tropical freshwater system (Lake Tanganyika). Environ Microbiol 8:1857–1863
Senn DB, Hemond HF (2002) Nitrate controls on iron and arsenic in an urban lake. Science 296:2373–2376
Shelobolina E, Konishi H, Xu HF, Benzine J, Xiong MY, Wu T, Blothe M, Roden E (2012) Isolation of phyllosilicate-iron redox cycling microorganisms from an illite-smectite rich hydromorphic soil. Front Microbiol 3:134
Silva LCF, Lima HS, Sartoratto A, de Sousa MP, Torres APR, de Souza RS, de Paula SO, de Oliveira VM, da Silva CC (2018) Effect of salinity in heterotrophic nitrification/aerobic denitrification performed by acclimated microbiota from oil-produced water biological treatment system. Int Biodeterior Biodegradation 130:1–7
Šimek M, Jı́šová L, Hopkins DW (2002) What is the so-called optimum pH for denitrification in soil? Soil Biol Biochem 34:1227–1234
Smith MS, Tiedje JM (1979) Phases of denitrification hollowing oxygen depletion in soil. Soil Biol Biochem 11:261–267
State Environmental Protection Administration of the People’s Republic of China (2002) Determination methods for examination of water and wastewater, fourth ed. China Environmental Science Press, Beijing
Straub KL, Buchholz-Cleven BEE (1998) Enumeration and detection of anaerobic ferrous iron-oxidizing, nitrate-reducing bacteria from diverse European sediments. Appl Environ Microbiol 64:4846–4856
Straub KL, Benz M, Schink B, Widdel F (1996) Anaerobic, nitrate-dependent microbial oxidation of ferrous iron. Appl Environ Microbiol 62:1458–1460
Tiedje JM (1994) Denitrifiers. In: Bottomley PS, Angle JS, Weaver RW (eds) Methods of soil analysis, Part 2. Microbiological and biochemical properties. Soil Science Society of America Press, Madison, pp 253–254
Valderrama JC (1981) The simultaneous analysis of total nitrogen and phosphorus in natural waters. Mar Chem 10:109–122
Viollier E, Inglett PW, Hunter K, Roychoudhury AN, Cappellen PV (2000) The ferrozine method revisited: Fe(II)/Fe(III) determination in natural waters. Appl Geochem 15:785–790
Wang ML, Hu RG, Zhao JS, Kuzyakov Y, Liu SR (2016) Iron oxidation affects nitrous oxide emissions via donating electrons to denitrification in paddy soils. Geoderma 271:173–180
Weber KA, Achenbach LA, Coates JD (2006a) Microorganisms pumping iron: anaerobic microbial iron oxidation and reduction. Nat Rev Microbiol 4:752–764
Weber KA, Pollock J, Cole KA, O’Connor SM, Achenbach LA, Coates JD (2006b) Anaerobic nitrate-dependent iron(II) bio-oxidation by a novel lithoautotrophic betaproteobacterium, strain 2002. Appl Environ Microbiol 72:686–694
Weber KA, Urrutia MM, Churchill PF, Kukkadapu RK, Roden EE (2006c) Anaerobic redox cycling of iron by freshwater sediment microorganisms. Environ Microbiol 8:100–113
Weber KA, Hedrick DB, Peacock AD, Thrash JC, White DC, Achenbach LA, Coates JD (2009) Physiological and taxonomic description of the novel autotrophic, metal oxidizing bacterium, Pseudogulbenkiania sp. strain 2002. Appl Microbiol Biotechnol 83:555–565
Wenk CB, Blees J, Zopfi J, Veronesi M, Bourbonnais A, Schubert CJ, Niemann H, Lehmann MF (2013) Anaerobic ammonium oxidation (anammox) bacteria and sulfide-dependent denitrifiers coexist in the water column of a meromictic south-alpine lake. Limnol Oceanogr 58:1–12
Wu GX, Zhai XF, Jiang CG, Guan YT (2013) Effect of ammonium on nitrous oxide emission during denitrification with different electron donors. J Environ Sci 25:1131–1138
Wunderlich A, Meckenstock R, Einsiedl F (2012) Effect of different carbon substrates on nitrate stable isotope fractionation during microbial denitrification. Environ Sci Technol 46:4861–4868
Yang XA, Huang S, Wu QH, Zhang RD (2012) Nitrate reduction coupled with microbial oxidation of sulfide in river sediment. J Soils Sediments 12:1435–1444
Yu JH, Fan CX, Zhong JC, Zhang L, Zhang L, Wang CH, Yao XL (2016) Effects of sediment dredging on nitrogen cycling in Lake Taihu, China: insight from mass balance based on a 2-year field study. Environ Sci Pollut Res 23:3871–3883
Zhang HH, Feng J, Chen SN, Zhao ZF, Li BQ, Wang Y, Jia JY, Li SL, Wang Y, Yan MM, Lu KY, Hao HY (2019) Geographical patterns of nirS gene abundance and nirS-type denitrifying bacterial community associated with activated sludge from different wastewater treatment plants. Microb Ecol 77:304–316
Funding
This research was supported by National Key R & D Program of China (No. 2017YFC0505305).
Author information
Authors and Affiliations
Corresponding author
Additional information
Responsible editor: Haihan Zhang
Publisher’s note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
About this article

Cite this article
Cheng, B., Hua, Y., Zhao, J. et al. Nitrogen transformation mediated by nitrate-dependent iron oxidation in anoxic freshwater. J Soils Sediments 20, 1087–1096 (2020). https://doi.org/10.1007/s11368-019-02461-w
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s11368-019-02461-w