
Abstract
Purpose
The farming practices of ridge-furrow with film mulching (RFFM) have been extensively reported as a valuable technique for improving crop productivity and enhancing water-use efficiency. However, research on the high-yield mechanism of RFFM measures under drought stress from the perspective of microbial diversity has been scared.
Materials and methods
Field experiments were conducted to analyze the effects of conventional flat planting (FP), no film mulching (NR), white polyethylene film mulching (WR), black polyethylene film mulching (BR), and biodegradable film mulching (BDR) with 40 and 80 cm furrow width levels on soil temperature, soil water storage (SWS), microbial carbon utilization, and yield of winter oilseed rape.
Results and discussion
The RFFM significantly increased the SWS as well as rapeseed yield and yield components. However, these treatments did not cause obvious differences in soil temperature. A relative high value of average color change rate (AWCD) was observed during the incubation time of 24–216 h, and it reached the peak value at 240 h. The AWCD in the furrow with 80 cm width was greater than that of 40 cm. The RFFM also could improve the utilization intensities of soil microorganism on the carbon sources of carbohydrates, amino acids, and miscellaneous in rapeseed rhizosphere. This result was consistent with that of the principal component analysis. The Shannon–Wiener diversity index, Simpson’s diversity index, and Shannon–Wiener evenness in BDR40, BR40, and BR80 were significantly higher than other treatments, respectively. In addition, correlation analysis results suggested that the indicator of affecting the diversity of carbon source was mainly due to the water storage capacity.
Conclusions
The methods of film mulching have a remarkable effect on microbial diversity. The yield of BR and BDR were significantly higher than other treatments. Compared with FP, RFFM increased rapeseed yield by 9.61–86.10%. In the year of less rainfall, production has decreased in comparison with the first season, while BDR80 guarantees continuous high yield and substantially promoted carbon utilization and soil microbial diversity. Soil microbial diversity is positively correlated with soil water content, which is beneficial to increase production. Wider furrow (80 cm) is recommended a sustainable high-yield method due to convenient labor in the region in Southwest China.
Similar content being viewed by others

Explore related subjects
Discover the latest articles, news and stories from top researchers in related subjects.Avoid common mistakes on your manuscript.
1 Introduction
In Southwest of China, most of farming lands are mainly mountainous and hilly areas with shallow soil layer and inevitable soil erosion problem, which limit in achieving high crop yield. Moreover, unpredictable rainwater, high evaporation rate, and low water utilization efficiency (Qin et al. 2014; Mo et al. 2017) further aggravate the situation and reduce agricultural productivity significantly. Recently, numerous agronomic management strategies such as drip irrigation (Pascual-Seva et al. 2018), manure application (Gu et al. 2018), mulching with crop straw and plastic film (Bandopadhyay et al. 2018; Dong et al. 2018; Yadav et al. 2018), and rainwater harvesting (Wang et al. 2009; Zhao et al. 2014; Zhang et al. 2017a, b, c; Wu et al. 2018) have been developed and adopted to enhance the efficiency of rainwater utilization and crop productivity.
The farming systems of ridge-furrow with film mulching (RFFM) are one of the most effective techniques to enhance rainwater utilization and increase crop productivity; these have recently been highly recommended as the feasible strategies (Zhou et al. 2012; Zhao et al. 2014; Mo et al. 2016; Zhang et al. 2017a, b, c; Liu et al. 2018; Qin et al. 2018). Ridge-furrow micro-water harvesting can collect runoff from ridges and concentrate water in furrows (Qin et al. 2014; Zhang et al. 2017a, b, c), and plastic-film mulching can reduce soil water evaporation and increase soil temperature in dryland farming systems simultaneously (Gu et al. 2016, 2017), thereby increase agricultural production (Mo et al. 2016; Liu et al. 2018; Wu et al. 2018). RFFM practices also decline weed competition, soil erosion, and soil compaction (Thankamani et al. 2016). Moreover, RFFM has been reported as a very promising and useful technique to increase the production of wheat (Dong et al. 2018; Liu et al. 2018), tomato (Moreno and Moreno 2008), maize (Mo et al. 2016; Li et al. 2017; Qin et al. 2018), and upland rice (Yadav et al. 2018) compared with conventional flat planting. The mechanisms responsible for dryland crop productivity under RFFM practices are predominantly attributable to the following aspects. Firstly, the shape of alternating ridges and furrows contribute to collecting rainwater (Mo et al. 2017; Qin et al. 2018), distributing soil moisture (Li et al. 2017; Liu et al. 2018), and mulching further mitigated the amount of water loss due to evaporation (Qin et al. 2014; Zhao et al. 2014). Secondly, mulching is effective in enhancing the topsoil temperature to ensure seedling emergence and establishment in the early growing season (Zhang et al. 2017a, b, c; Gu et al. 2018). However, the co-application effects of different furrow widths and mulching on crop production have not been sufficiently investigated.
Previous works mainly emphasized on the impacts of RFFM practices on water availability, soil temperature and yield (Li et al. 2017; Gu et al. 2018; Liu et al. 2018; Qin et al. 2018), soil biochemical processes like soil organic carbon (SOC) (Huang et al. 2008; Wang et al. 2016a, b; Dong et al. 2018), soil microbial biomass carbon (MBC) (Muñoz et al. 2017; Jin et al. 2018), and CO2 efflux (Zhang et al. 2017a, b, c; Yu et al. 2018), while their influences on the soil microecological environment and the diversity of soil microbial community were neglected. Soil microorganisms play a crucial role in agro-ecosystem function and sustainability because of their roles in nutrient biogeochemical cycling, soil structure forming, and crop productivity (Soong et al. 2018). Microbial diversity could be used as a sensitive determinant of soil quality, and any alteration or variation in microbial diversity in response to agricultural practices can provide valuable information for the evaluation of soil function (Fierer 2017). It was demonstrated that agricultural practices changed the soil microenvironment, by causing a shift in soil microorganisms (Zhang et al. 2018). Moreover, the disturbances caused by crops, plastic-film mulching, and water availability also alter soil microbial community composition and metabolic functional diversity. And to describe the alterations, community-level physiological profiles have been frequently employed (Huang et al. 2008; Bandopadhyay et al. 2018; Martínez-García et al. 2018). However, until now, the interaction between ridge-furrow micro-water harvesting and plastic-film mulching on soil microbial metabolic characteristics still remained poorly understood. Therefore, this study was conducted to understand the influence of RFFM farming systems on soil microenvironment and soil microbial diversity.
Oilseed rape (Brassica napus L.) is not only extensively employed to produce vegetable oil for human but is also seen as one of the promising renewable energy resources (Rathore et al. 2017; Keane et al. 2018). The winter oilseed rape is one of the most essential oil crops in China and is predominantly planted in the southwest areas. Some studies have shown that RFFM systems can increase rapeseed yield in arid and semiarid areas (Gu et al. 2017, 2018); however, few researchers have focused in studying the changes in microbial functional diversity in rapeseed ecosystems under RFFM practices. Therefore, the objectives of this study were to (1) evaluate the effects of RFFM planting patterns on the soil water availability, soil temperature, and yield and yield components of winter oilseed rape; and to (2) assess the effects of different RFFM cultivations on soil organic carbon, soil microbial biomass, respiration, functional diversity, and carbon utilization patterns of the microbial community. Results of this study will provide a better understanding of soil quality maintenance, crop production, and carbon management under the plastic film mulching and ridge-furrow technology in dryland farming systems.
2 Materials and methods
2.1 Experimental site description
A field experiment was performed during the two successive seasons of 2016–2017 and 2017–2018 in the Yunyang Experimental Station (108° 54′ E, 30° 55′ N; altitude of 700 m), Southwest University, Chongqing, China. The region has an average annual temperature of 18.7 °C, mean sunshine duration of 1500 h, and an annual mean precipitation of approximately 900 mm, and the rain period predominantly prolongs from June to September. Local soil type is clay loam in texture and Dystric Purpil-Udic Cambosols according to the Chinese Soil Taxonomy (CRGCST 2001). A mean soil bulk density in the 0–60 cm soil layer is 1.44 g cm−3, and the chemical properties of 0–20-cm soil layer at the beginning of the experiment are organic matter of 12.58 g kg−1, total nitrogen of 0.98 g kg−1, total phosphorus of 0.56 g kg−1, total potassium of 29.15 g kg−1, available nitrogen of 70 mg kg−1, available phosphorus of 15.53 mg kg−1, available potassium of 306.11 mg kg−1, and a pH of 6.15 (soil:water = 1:5), respectively. The temperature and rainfall during the rapeseed growing seasons are described in Fig. S1 (Electronic Supplementary Material—ESM) recorded at the standard meteorological observation station near the experimental site. The preceding crop was sorghum in both seasons.
2.2 Experimental design
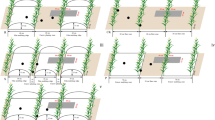
All the treatments were arranged in a randomized complete block design with three replications in both seasons (2016–2017 and 2017–2018). Two furrow widths and four types of film mulching were tested for soil microbial, carbon utilization, seed yield, and yield components of winter rapeseed. Nine treatments were established in this experiment including (1) FP: conventional flat planting without mulching, as a control group; (2) NR40: 40-cm furrow width without film mulching; (3) NR80: 80-cm furrow width without film mulching; (4) WR40: 40-cm furrow width with white transparent polyethylene film mulching; (5) WR80: 80-cm furrow width with white transparent polyethylene film mulching; (6) BR40: 40-cm furrow width with black polyethylene film mulching; (7) BR80: 80-cm furrow width with black polyethylene film mulching; (8) BDR40: 40-cm furrow width with biodegradable film mulching; (9) BDR80: 80-cm furrow width with biodegradable film mulching. All the films used in this test were 0.008 mm thick and 80 cm wide. In ridge-furrow treatments, ridge width and height were 40 and 15 cm, respectively. Plots were 5.2 m long × 4 m wide (size, 20.8 m2) with a 1-m-wide strip between plots. For the NR40, WR40, BR40, and BDR40 treatments, there were seven ridges and six furrows in each plot and two rows of rapeseed were planted in each furrow, while five ridges and four furrows in each plot were established in NR80, WR80, BR80, and BDR80 treatments and three rows of rapeseed were maintained in each furrow (Fig. S2—ESM). After plowing and leveling the experimental field, compound fertilizer (N:P:K = 14:16:15) at 225 kg ha−1 was applied as base fertilizer to ensure sufficient fertilizer supply to rapeseed in accordance with local farming practices. Rapeseed oil seeds “Sanxiayou No. 5” were manually sown in rows spaced 40 cm and holes spaced 33 cm apart. Seedlings were thinned by hand, and plant density was determined as two plants per hole at the fifth-leaf stage. No irrigation and no more fertilizer were applied during the growth of winter oilseed rape. All other agronomic practices were consistent with the local.
2.3 Sampling and measurements
2.3.1 Determination of soil temperature and moisture
At the bud stage of rapeseed, soil surface temperature (20 cm depth) for each plot was recorded manually using a curved pipe geothermometer (WQG-16, Tianjin 8th Manufactory, China) which was placed in the center between two plants in the furrows.
Soil gravimetric water contents (SWC, %) were measured by a 0.08-m-diameter portable auger in all plots, from the surface to 60 cm depth at 20-cm intervals at bud stage of rapeseed. In each plot, three random sampling points were chosen in the middle part between two plants in the furrows. The gravimetric water content of the soil samples was determined by oven-drying at 105 °C. Soil water storage (SWS) was calculated according to the following formula:
where ρb is soil bulk density and SD is soil depth.
2.3.2 Measurement of carbon source utilization in rhizosphere soil
The five-spot-sampling method was applied for collecting the rhizosphere soil samples from rapeseed plants in all plots in March 2018. The roots were dug up cautiously, and the non-rhizosphere soil which was attached to the root surface loosely was removed (Shen et al. 2016). The rhizosphere soils (0–0.2 cm away from roots) were gently brushed down with a sterilized brush. All samples were placed into sterile plastic bags, then placed in an ice box, and then transported to the laboratory within 5 h after collection and were stored at − 4 °C for the analysis of carbon substrate utilization profiles within a week.
The potential metabolic diversity of carbon of microbial communities in rhizosphere soils was determined using commercial Biolog EcoPlate™ (Biolog Inc., CA, USA) (Martínez-García et al. 2018; Zhang et al. 2018). Each plate consists of 96 micro-holes containing three replicates, each of one control well without any carbon source and 31 separate sole carbon substrates. The carbon sources were classified into six guilds namely carbohydrates, carboxylic acids, amino acids, polymers, amines/amides, and miscellaneous (Table S1—ESM). Fresh rhizosphere soil samples (equivalent to 5 g of dry soil) were suspended in 45 mL of sterile 0.85% NaCl solution in a brown bottle and shaken for 30 min at 150 rpm with an orbital shaker at 25 °C. After settling for 10 min, the supernatant was diluted to a final 10−3 using a 0.85% NaCl solution. Subsequently, 150 μL of the diluted soil suspensions were transferred into each of the wells of the EcoPlate. The plates were incubated at 28 °C in a humidity-saturated and dark environment for 240 h. The color development in each well over time was quantified at 590 nm using an EL808 Ultra Microplate Reader (BioTek Instruments, Winooski, VT, USA) at the regular time intervals of 24 h. The absorbance of each carbon source subsequently was corrected by subtracting the absorbance of the blank. Wells that had negative absorbance values were set to zero for the further analyses. Optical density values obtained at 120 h/incubation in which for the microbial activity had stabilized at this time were used for assessing microbial community function, functional diversity, and posterior analyses (Huang et al. 2008).
Average well color development (AWCD), representing the overall carbon substrate utilization potential of cultural microbial communities across all wells per plate (Garland and Mills 1991), was calculated as:
where Ai is the absorbance reading of well i and A0 represents the absorbance reading of the control well.
The functional diversity of microbial communities in rhizosphere soil in RFFM tillage system was indicated by the Shannon–Wiener diversity index (H′, Eq. (3)) (Keylock 2005):
Simpson’s diversity index (D, Eq. (4)) was calculated as (Hunter and Gaston 1988):
Shannon–Wiener evenness (E) represented the equitability of activities across all utilized substrates (Eq. (5)) (Keylock 2005):
where pi is the ratio of the absorbance of a particular well i to the sums of absorbance of all 31 well at 120 h. Substrate richness (S), reflecting a positive response to the utilization of substrates, was calculated as the number of wells with a corrected absorbance at 590 nm higher than 0.25.
2.3.3 Analysis of soil biochemical properties
Each plot was randomly placed with three soil respiration collars (polyvinylchloride with 10 cm diameter and 5 cm height) between the two holes before sowing and kept them clean during the experiment. Soil-surface CO2 efflux within the collars was measured from 9:00 a.m. to 11:00 a.m. by using the Li-6400-09 soil chamber attached to the Li-6400 portable gas exchange system (Li-Cor, Lincoln NE, USA). The CO2 drawdown-rise cycle was replicated three times, and the integrated CO2 efflux values were averaged for each plot. Soil respiration rate was measured on the same day as soil sample collections at the bud stage of rapeseed in March 2018. The SOC content was determined by the potassium dichromate volumetric method (Nelson and Sommers 1996). Soil microbial biomass carbon (MBC) was analyzed by the chloroform fumigation–extraction method described previously (Vance et al. 1987).
2.3.4 Seed yield and yield components
Experimental plots were harvested when approximately 2/3 of seeds were brown or dark brown to black. Plants were randomly sampled from a 1 m × 1 m area of each plot and slowly uprooted by hands; thus, the taproot and large lateral roots were retained. Then, the yield components and seed yield were measured. For each plant, the following quantifications and observations were done: the plant height (cm), siliques per plant, seeds per silique, branches per plant, and thousand-seed weight (g). The remaining plants in each plot were individually harvested to estimate the seed yield (kg ha−1) per unit area after air-drying. The plant height was measured from the cotyledonary node to the ligule of the uppermost amply expanded leaf, and the average height was judged from all measured plants.
2.4 Statistical analyses
All the presented data are means ± standard deviations of a specified number of replicates. The recorded data were subjected to analysis of variance to evaluate the effects of ridging, covering, and their interactions on all measured parameters using the Statistical Packages for the Social Sciences software version 22.0 (SPSS Inc., USA). Significant differences between different treatments were compared by the least significant difference (LSD) test at a 0.05 probability level. Graphics were made using OriginPro 9.0 software (OriginLab Corp., Northampton, MA, USA). Correlation analysis was performed to determine the relationship between the diversity indexes of soil microbial communities, carbon source utilization, soil carbon content, and other factors. Principal component analysis (PCA) was conducted to compare the relative similarity in the carbon metabolic patterns of different rhizosphere soils using Canoco software (v5.0).
3 Results
3.1 Soil hydrothermal characteristic, yield, and agronomic traits
Ridging and mulching practices significantly increased SWS to a depth of 60 cm (P < 0.05, Fig. 1). The BDR treatment had the maximum SWS at a ridge-furrow ratio of 40 cm: 80 cm with an increased rate of 11.92% compared to the control (FP). Under NR treatment, SWS were increased by 3.18 to 5.63% as compared with FP. SWS in the furrow width of 80 cm was higher than that in the furrow width of 40 cm under the same ridge covering materials except for BR treatments. In contrast to SWS, ridging and mulching practices had no significant effect on soil temperature when compared to FP (Fig. 1).

Mean values of soil water storage amount in soil profile of 0–60 cm and soil temperature (20 cm depth) under different ridging-mulching film practices (NR, no film mulching in the ridge; WR, ridges covered with white transparent polyethylene film mulch; BR, ridges covered with black polyethylene film mulch; BDR, ridges covered with biodegradable film mulch; FP, traditional flat planting). Two different ridge-furrow ratios of 40 cm:40 cm and 40 cm:80 cm planting levels were used. Means (n = 3) followed by different lowercase letters indicate significant differences at the P < 0.05 level between treatments according to the LSD’s test. Error bars indicate the standard deviation of the mean
Ridging and mulching practices also significantly affected rapeseed yield and its agronomic traits as compared with conventional flat sowing in both seasons (Table 1). As compared with FP and NR, WR, BR, and BDR treatments considerably improved plant height, siliques per plant, seeds per silique, branches per plant, and thousand-seed weight of rapeseed under both furrow widths. In the first season, the rapeseed yield under NR40, NR80, WR40, WR80, BR40, BR80, BDR40, and BDR80 treatments were significantly improved by 14.23, 13.41, 46.92, 37.97, 86.10, 69.39, 56.67, and 61.61%, respectively, than that of FP treatment. The increase rate of grain yield in the second season was lower than that in the first season with a maximum value of 72.37% in BDR80 treatment. Additionally, higher furrow width led to relatively lower yield performance.
3.2 Metabolic characteristics of soil microbial carbon sources
As shown in Fig. 2, AWCD values were negligible in the first 24 h and increased sharply thereafter, suggesting that during initial 24 h, the carbon utilization rate from the Biolog EcoPlate was intensely low; however, it was rapidly raised after 24 h. AWCD levels in ridge-furrow and mulching practices were substantially higher than that in the FP, which manifested that the carbon metabolism abilities of microbial communities were stronger in ridge-furrow and mulching film systems. The maximum AWCD value within the incubation period of 24–216 h was obtained in WR80 treatment. A declining trend in AWCD was observed for all treatments after 192 h of culture except for FP and BR80, and the maximum reduction was found in NR treatment. Compared with the 40-cm furrow width treatment, the AWCD value was higher in the 80-cm furrow width, indicating a stronger metabolic activity of microbial community in the rhizosphere soils.

The changes in AWCD of all carbon sources in rhizosphere soil microbial communities within incubation time under different ridging-mulching film practices (NR, no film mulching in the ridge; WR, ridges covered with white transparent polyethylene film mulch; BR, ridges covered with black polyethylene film mulch; BDR, ridges covered with biodegradable film mulch; FP, traditional flat planting). Two furrow widths of 40 cm:40 cm and 40 cm:80 cm planting levels were used
3.3 Carbon source utilization of soil microbial
The microbial could use all six substrate categories, and relative substrate utilization varied from 9.29 to 23.53%. However, the patterns of C-substrate utilization were remarkably affected by the ridge-furrow and mulching (Fig. 3). From the metabolic fingerprint of Biolog EcoPlate, amino acids, carboxylic acids, and miscellaneous-consuming microorganisms were dominant irrespective of treatments in the different film covering material and furrow width of rapeseed production system. The utilization proportion of carbohydrates in ridge-furrow and mulching treatments were considerably higher than FP (P < 0.05). Planting with furrow width of 80 cm was conducive to the carbohydrate utilization by soil microbes in comparison with a furrow width of 40 cm under the same type of film treatment. Among them, the highest proportion in carbohydrate utilization was 18.32% in BR80 treatment followed by BDR80 treatment (18.04%). Although, the carbohydrate utilization proportion in NR treatment was 1.41–3.26% higher than that of FP treatment, and the increase was lower than that of other film mulching treatments. The miscellaneous utilization in BDR40 treatment accounted for 20.73% of the total carbon source, followed by WR80 (19.09%). It maintained at 16–17% under the treatments of non-film, black film, and degradation film covering, while was only 12–14% under the treatment of white film covering. The change in utilization of amines/amides was opposite to miscellaneous. Consequently, the rain-harvesting planting could enhance the utilization intensities of carbohydrates, amino acids, and miscellaneous by soil microorganisms in the rhizosphere of rapeseed.

Relative substrate utilization for six substrate categories in each treatment. Soil samples were tested from rhizosphere of rapeseed treated with different ridging and mulching practices (NR, no film mulching in the ridge; WR, ridges covered with white transparent polyethylene film mulch; BR, ridges covered with black polyethylene film mulch; BDR, ridges covered with biodegradable film mulch; FP, traditional flat planting). Two furrow widths of 40 cm:40 cm and 40 cm:80 cm planting levels were used
As illustrated in Fig. 4a, d-cellobiose and α-d-lactose accounted for a low (< 5.0%) proportion of carbohydrate sources. An uneven distribution of carbohydrate utilization by FP treatment was observed with 31.14% of N-acetyl-d-glucosamine and 35.44% of d-mannitol. However, the soil microbial utilization of carbohydrate carbon sources was relatively uniform under WR, BR, and BDR treatments, suggesting that ridging and mulching practices improved the carbohydrate utilization. The utilization rates of the carboxylic acid carbon sources in each treatment were approximately between 10 to 20% except for α-ketobutyric acid and 2-hydroxybenzoic acid (Fig. 4b). l-arginine, l-asparagine, and l-serine were preferentially utilized by all rhizosphere soil communities (Fig. 4c). The rapeseed rhizosphere soils had the highest utilization of Tween 40 and Tween 80 in polymer-carbon sources (Fig. 4d). However, α-cyclodextrin utilization in the WR, BR, and BDR treatments was improved compared to FP. Based on the comprehensive analysis of the utilization of 31 carbon sources in different treatments, the ridging and mulching practices enhanced the diversity of microbial carbon source utilization and avoided the simplification of carbon source utilization.

The distribution of 31 carbon sources in corresponding to (a) carbohydrates, (b) carboxylic acids, (c) amino acids, (d) polymers, (e) amines/amides, and (f)miscellaneous, respectively, in rapeseed rhizosphere soil microbes carbon metabolism under different ridging and mulching practices (NR, no film mulching in the ridge; WR, ridges covered with white transparent polyethylene film mulch; BR, ridges covered with black polyethylene film mulch; BDR, ridges covered with biodegradable film mulch; FP, traditional flat planting). Two furrow widths of 40 cm:40 cm and 40 cm:80 cm planting levels were used. Carbon source: A2: β-methyl-d-glucoside, A3: d-galactonic acid γ-lactone, A4: l-arginine, B1: pyruvic acid methyl ester, B2: d-xylose, B3: d-galacturonic acid, B4: l-asparagine, C1: Tween 40, C2: i-erythritol, C3: 2-hydroxybenzoic acid, C4: l-phenylalanine, D1: Tween 80, D2: d-mannitol, D3: 4-hydroxybenzoic acid, D4: l-serine, E1: α-cyclodextrin, E2: N-acetyl-d-glucosamine, E3: γ-hydroxybutyric acid, E4: l-threonine, F1: glycogen, F2: d-glucosaminic acid, F3: itaconic acid, F4: glycyl-l-glutamic acid, G1: d-cellobiose, G2: glucose-1-phosphate, G3: α-ketobutyric acid, G4: phenylethylamine, H1:α-d-lactose, H2: d,l-α-glycerol phosphate, H3: d-malic acid, H4: putrescine
As shown in Table 2, compared to FP, the cultivation method of ridging and mulching had remarkably increased the functional diversity indexes of rhizosphere microorganisms. The indexes of H′, E, and D in BR40 were relatively higher and increased by 9.00, 4.68, and 2.89%, respectively, compared with FP treatment, indicating that the species richness, the individuals distributed, and evenness of microorganisms in rapeseed rhizosphere were relatively high under BR40. Furthermore, the rate of substrate utilization by soil microbial community was found to be consistently greater in the BDR treatments than in the FP treatment.
3.4 Soil carbon content, soil sucrose activity, and soil respiration
As shown in Table 3, the ridging planting treatments considerably affected the SOC content and soil sucrase activity (P < 0.05) while the mulching planting treatments dramatically influenced the SOC content and soil sucrase activity (P < 0.05), but the interaction between ridging × mulching was non-significant (P > 0.05). Increasing furrow width was conducive to the accumulation of SOC in the soil; however, soil sucrase activity showed the opposite trend. There was no significant difference in SOC in the same furrow width under different film mulching (P > 0.05). Compared with FP, the ridging and mulching practices did not increase SOC content. The WR80 treatment had the highest MBC of 186.05 mg kg−1, about 1.25–1.58 times higher than the MBC of the other treatments. Soil respiration could reveal the effectiveness of easily mineralized organic carbon in soil. Soil respiration activity in the WR40, BDR40, and BDR80 treatments was significantly higher than FP by 55.94, 62.83, and 52.87%, respectively (P < 0.05).
3.5 Principal component analysis
The PCA results showed distinct differences between ridging and mulching treatments (Fig. 5). PCA reduced the initial dimension of the dataset to two principal components, which explained 70.0% of the data variation. The PC1 accounted for 53.6% of the total variance, and PC2 described 16.4%. The loading scores from PCA showed similarity degree to carbon source utilization. The higher loading scores were, the larger effects of carbon source on the principal components were. FP and NR treatments were widely separated from each other on PC1 axis; however, PC1 could not separate in WR, BR, and BDR treatments, suggesting that FP and NR treatments had little effect on the utilization of microbial carbon sources and rhizosphere soil in ridging and mulching practices had a similar degree of similarity to carbon source utilization. Phenylethylamine, putrescine, itaconic acid, and γ-hydroxybutyric acid were the decisive carbon sources in NR treatment. While for WR treatment, the carbon sources of carbohydrates (d-xylose and i-erythritol), amino acids (l-asparagine and l-serine), and carboxylic acids (d-galacturonic acid and d-glucosaminic acid) were the crucial contributor. In addition, d-cellobiose, and N-acetyl-d-glucosamine in carbohydrates, l-arginine in amino acids, and glycogen and Tween 80 in polymers played a prominent role in the separation of the two major components. From the results of carbon source analysis above, it was obvious that the carbon sources utilized by rapeseed rhizosphere soil microbial communities were carbohydrates, amino acids, carboxylic acids, and polymers.

Principal component analysis of different carbons sources of soil microbial diversity in rapeseed rhizosphere in different ridging and mulching practices (NR, no film mulching in the ridge; WR, ridges covered with white transparent polyethylene film mulch; BR, ridges covered with black polyethylene film mulch; BDR, ridges covered with biodegradable film mulch; FP, traditional flat planting). Two furrow widths of 40 cm:40 cm and 40 cm:80 cm planting levels were used. Carbon source: A2: β-methyl-d-glucoside, A3: d-galactonic acid γ-lactone, A4: l-arginine, B1: pyruvic acid methyl ester, B2: d-xylose, B3: d-galacturonic acid, B4: l-asparagine, C1: Tween 40, C2: i-erythritol, C3: 2-hydroxybenzoic acid, C4: l-phenylalanine, D1: Tween 80, D2: d-mannitol, D3: 4-hydroxybenzoic acid, D4: l-serine, E1: α-cyclodextrin, E2: N-acetyl-d-glucosamine, E3: γ-hydroxybutyric acid, E4: l-threonine, F1: glycogen, F2: d-glucosaminic acid, F3: itaconic acid, F4: glycyl-l-glutamic acid, G1: d-cellobiose, G2: glucose-1-phosphate, G3: α-ketobutyric acid, G4: phenylethylamine, H1:α-d-lactose, H2: d,lα-glycerol phosphate, H3: d-malic acid, H4: putrescine
4 Discussion
4.1 Effect of cultivation systems on soil hydrothermal characteristics and rapeseed production
The RFFM farming systems can tremendously improve soil water availability and thermal conditions in the crop root zone compared with flat planting patterns by collecting runoff rainwater from ridges to furrows, as well as preventing soil evaporation and reducing water loss (Mo et al. 2016; Zhang et al. 2017a, b, c). Similarly, in the present study, the average soil SWS (0–60 cm) at the bud stage of rapeseed increased significantly (P < 0.05, Fig. 1) for WR40, WR80, BR40, BR80, BDR40, and BDR80, i.e., by 4.66, 6.41, 6.25, 0.99, 5.56, and 11.92% compared with FP, respectively. The rainfall infiltrated laterally into the ridges, and it was ultimately accumulated in the furrows, where it was available for the plant under water stress conditions (Gu et al. 2017; Zhang et al. 2017a, b, c). Previous studies have demonstrated that a variety of ridge-mulching materials enhanced the rainwater utilization from soil and improved rapeseed productivity (Gu et al. 2017, 2018). The benefits were intensified as the furrow widths increased (Fig. 1), due to reducing water losses from soil surface by mulching, which agreed with the results of Li et al. (2017).
Plastic film mulching considerably improves the rhizosphere soil temperature and alleviates the low-temperature stress on the plant during winter (Mo et al. 2016; Qin et al. 2018). Generally, the influence of film mulching on soil temperature relies on the balance of reflecting the ability of the mulch materials and transmission of solar energy (Liu et al. 2018). In this study, the warming effects of RFFM planting practices were not obvious; contrarily, slight cooling effects were detected in the furrow widths of 40 cm. Compared with the FP, the NR80, WR80, and BDR80 had negligible effects on soil temperature (P > 0.05), while the other treatments conspicuously decreased the soil temperatures (P < 0.05). The reason for this was probably that black plastic film could absorb most of the solar radiation (Qin et al. 2018), lowering the soil temperature more than biodegradable film and white plastic film. In addition, a higher plant coverage and lower sunlight transmission in the rapeseed bud stage intercepted the majority of solar energy reaching the soil, which might lead to the results that the RFFM had no obvious effect on soil temperature. Mo et al. (2017) also stated that the soil coverage with black film slightly diminished topsoil temperature in comparison with that of the control, for it blocked the passages of most solar radiation.
In dryland farming systems, crop productivity predominantly relies on rainfall, and the yield extensively correlates with rainfall at critical growth stages (Liu et al. 2018). The RFFM planting patterns lower soil evaporation, boost the infiltration of rainwater into the deep soil (Ramakrishna et al. 2006), and increase soil water retention, which improve crop growth, development, and grain yield (Gu et al. 2016; Li et al. 2017). In this study, seed yield was markedly higher in the mulching treatments than FP by 9.61–86.10% over the two growing seasons (Table 1). These results coincided with previous works in wheat (Dong et al. 2018; Liu et al. 2018), tomato (Moreno and Moreno 2008), maize (Mo et al. 2016, 2017; Li et al. 2017; Qin et al. 2018), and upland rice (Yadav et al. 2018). Surface cover, improvement in rainwater harvesting, and soil moisture conservation under film mulching were greater than that of conventional practice (Lin et al. 2016).
4.2 Microbial responses to RFFM practices
In the present study, Biolog EcoPlate™ was employed to evaluate the community level physiological profiles of soil microorganism communities as suggested by previous works (Huang et al. 2008; Shen et al. 2016), which would offer metabolic characterization on the basis of 31 carbon sources. These results showed that the AWCD levels in RFFM farming systems were dramatically higher than that in FP, and the maximum value within the incubation period of 24–216 h was obtained at WR80 treatment (Fig. 2). Wang et al. (2016a, b) also reported that the AWCD of soil microorganisms in white film mulching was more sensitive than in black film mulching. The greater potential metabolic functional diversity of soil microbial communities in RFFM treatments (Table 2) was in agreement with the studies by Huang et al. (2008), indicating that more carbon source could be utilized under ridge-furrow with film mulching. In addition, evidence of enhanced degradative activities and breathability of the biodegradable film than polyethylene suggested that it was beneficial to the abundance of the diversity of microbial carbon source utilization (Moreno and Moreno 2008; Bandopadhyay et al. 2018).
PCA analysis of community-level physiological profiles demonstrated that microorganisms of the RFFM treatments had better availability to utilize different carbon sources (Fig. 5). The utilization of amino acids, carboxylic acids, and miscellaneous influenced the differentiation of carbon sources to use patterns of microbial communities in the different film covering material and furrow width of rapeseed production system. Huang et al. (2008) have tested the variability of soil microbial communities in the subtropical zone and found that separation of the mulch and non-mulch treatments were mainly affected by carbohydrate and carboxylic acids. Our results displayed that film types and furrow widths dominated the carbon source utilization by soil microorganism in RFFM. Favorable light, moisture, and temperature conditions and presence of roots modulate microbial activity under RFFM (Lin et al. 2016; Gu et al. 2018). Nonetheless, these responses might be most likely dependent on the amount of warming under film mulching: where in warmer seasons, mulching might push temperatures above optima, restricting soil microbial activities, whereas during low temperature regimes, mulching-induced soil temperature closer to microbial optima and increased their activities (Moreno and Moreno 2008). The RFFM tillage systems cause the heterogeneity of the soil microenvironment due to the enhanced temperature and humidity in the soil, which forms a unique rhizosphere soil microclimate environment (Zhang et al. 2008). Therefore, the RFFM tillage systems improved microbial carbon source utilization diversities and avoided their simplification, but dehumidification should be cautiously considered especially in the Southwestern region of China where there is a high rainfall in the early stage of rapeseed planting (Fig. S1—ESM).
Soil microbial functional diversity correlated with soil organic carbon has been proposed by Huang et al. (2008). However, rather than supporting the assumption that the soil total organic carbon, soil microbial biomass carbon, soil respiration, or soil sucrose activity control the functional diversity of soil microbial community, these studies suggested that SWS was more indicative of the change of functional diversity of soil microbial community (Table 4). Good correlations between SWS and carbohydrates and polymers were also observed. Therefore, increase in SWS was contributed to increasing soil microbial diversity in RFFM farming systems. However, to the best of our knowledge, this is the first work which has examined the close correlation between SWS and the shift in the diversity of soil microbial function. Further investigations of the relationship should be necessary to corroborate this conclusion.
4.3 Effect of RFFM practices on soil organic carbon metabolism
Suitable tillage management measures are important for the improvement of soil enzyme activity, promotion of the decomposition and transformation of soil organic matter, and enrichment of the functional diversity of rhizosphere soil microorganisms (de Moraes Sá et al. 2018; Dong et al. 2018; Martínez-García et al. 2018). RFFM contributed to a decrease in the SOC through increasing soil carbon mineralization and microbial activity (Wang et al. 2016a, b; Dong et al. 2018), suggesting that RFFM may not be a long-term solution in managing the soil. Conversely, other studies stated that RFFM could lead to the positive balances of soil carbon because of the increase in root growth and exudate secretion, and microbial decomposition (Bandopadhyay et al. 2018). Compared with the FP, RFFM practices resulted in the decrease of SOC concentration or no change in this study (Table 3), indicating that RFFM favored a more quick conversion of SOC by assuring suitable microecological environment for microorganisms. This finding was inconsistent with previous studies (Cuello et al. 2015). The MBC only took about 1–5% of SOC, but it had a significant influence on the rate of nutrient cycling in agricultural ecosystems (Cuello et al. 2015). In this study, the remarkably higher MBC concentrations in RFFM practices compared to FP were observed (Table 3). Previous investigation showed that RFFM practices increased MBC in the topsoil compared with the conventional plat planting, and the higher topsoil temperature and moisture conditions played a crucial role in the changes of MBC in dryland farming systems (Dong et al. 2018). It has also been suggested that RFFM practices could enhance soil biochemical properties and soil sucrase activity (Zhou et al. 2012). However, it should be noted that a great variety of microorganisms decreased because of changes in microclimate under continuous RFFM practices, and consequently, the content of MBC also decreased (Li et al. 2004).
The RFFM practices could profoundly alter the ways of crop root distribution. In this investigation, the CO2 efflux rate was 11.49–62.84% higher in RFFM compared with FP (Table 3). Majority of roots predominantly centered under ridge-furrow soil layer (Gao et al. 2014) and then SWS increased (Wu et al. 2018), and a relatively higher soil temperature in the furrow soil area promoted gas molecule activity and gas release rate (Davidson and Janssens 2006), which may enhance the soil respiration by strengthening the rhizosphere activities in ridge-furrow cropping system (Zhang et al. 2017a, b, c).
5 Conclusions
This study elucidated ridge-furrow with film mulching with two furrow widths can enhance the soil moisture storage and availability in the furrow and can regulate the soil temperature. Black plastic and biodegradable film mulching increased more rapeseed yield than white plastic film mulching. This approach also significantly improved the diversity of soil microbial carbon source utilization and further validated the effect of soil water ecological process optimization, namely, the increase of soil water storage capacity on it. However, for the field planting of winter rapeseed in Southwest China, it was also necessary to consider the dehumidification in the rainy season. The diversity of soil microbial carbon source utilization, soil organic carbon, and soil microbial biomass carbon were higher in a furrow width of 80 cm than in a furrow with a width of 40 cm. White film mulching increased the microbial carbon source utilization diversity and microbial biomass carbon, while black film and biodegradable film mulching had a stronger biological activity and increased the carbohydrates, amino acids, and miscellaneous carbon source utilization ability. Taken together, these results indicated that ridge-furrow with film mulching planting rather than conventional plat planting is a good practice for maintaining microbial carbon metabolism and altering microbial functional diversity. The challenge in the future will be to better understand the variation in soil microbial community over time under different mulching treatments in different locations and elucidate the expression of functional genes within each community.
References
Bandopadhyay S, Martin-Closas L, Pelacho AM, DeBruyn JM (2018) Biodegradable plastic mulch films: impacts on soil microbial communities and ecosystem functions. Front Microbiol 9:819
CRGCST (Cooperative Research Group on Chinese Soil Taxonomy) (2001) Chinese soil taxonomy. Science Press, Beijing, pp 166–167
Cuello JP, Hwang HY, Gutierrez J, Kim SY, Kim PJ (2015) Impact of plastic film mulching on increasing greenhouse gas emissions in temperate upland soil during maize cultivation. Appl Soil Ecol 91:48–57
Davidson EA, Janssens IA (2006) Temperature sensitivity of soil carbon decomposition and feedbacks to climate change. Nature 440:165–173
de Moraes Sá JC, Potma Gonçalves DR, Ferreira LA, Mishra U, Inagaki TM, Ferreira Furlan FJ, Moro RS, Floriani N, Briedis C, de Oliveira Ferreira A (2018) Soil carbon fractions and biological activity based indices can be used to study the impact of land management and ecological successions. Ecol Indic 84:96–105
Dong QG, Yang Y, Yu K, Feng H (2018) Effects of straw mulching and plastic film mulching on improving soil organic carbon and nitrogen fractions, crop yield and water use efficiency in the Loess Plateau, China. Agric Water Manag 201:133–143
Fierer N (2017) Embracing the unknown: disentangling the complexities of the soil microbiome. Nat Rev Microbiol 15:579–590
Gao Y, Xie Y, Jiang H, Wu B, Niu J (2014) Soil water status and root distribution across the rooting zone in maize with plastic film mulching. Field Crop Res 156:40–47
Garland JL, Mills AL (1991) Classification and characterization of heterotrophic microbial communities on the basis of patterns of community-level sole-carbon-source utilization. Appl Environ Microbiol 57:2351–2359
Gu X, Li Y, Du Y (2016) Continuous ridges with film mulching improve soil water content, root growth, seed yield and water use efficiency of winter oilseed rape. Ind Crop Prod 85:139–148
Gu X, Li Y, Du Y (2017) Biodegradable film mulching improves soil temperature, moisture and seed yield of winter oilseed rape (Brassica napus L.). Soil Tillage Res 171:42–50
Gu X, Li Y, Du Y (2018) Effects of ridge-furrow film mulching and nitrogen fertilization on growth, seed yield and water productivity of winter oilseed rape (Brassica napus L.) in Northwestern China. Agric Water Manag 200:60–70
Huang Z, Xu Z, Chen C (2008) Effect of mulching on labile soil organic matter pools, microbial community functional diversity and nitrogen transformations in two hardwood plantations of subtropical Australia. Appl Soil Ecol 40:229–239
Hunter PR, Gaston MA (1988) Numerical index of the discriminatory ability of typing systems: an application of Simpson's index of diversity. J Clin Microbiol 26:2465–2466
Jin X, An T, Gall AR, Li S, Filley T, Wang J (2018) Enhanced conversion of newly-added maize straw to soil microbial biomass C under plastic film mulching and organic manure management. Geoderma 313:154–162
Keane BJ, Ineson P, Vallack HW, Blei E, Bentley M, Howarth S, McNamara NP, Rowe RL, Williams M, Toet S (2018) Greenhouse gas emissions from the energy crop oilseed rape (Brassica napus): the role of photosynthetically active radiation in diurnal N2O flux variation. GCB Bioenergy 10:306–319
Keylock CJ (2005) Simpson diversity and the Shannon-Wiener index as special cases of a generalized entropy. Oikos 109:203–207
Li F, Song Q, Jjemba PK, Shi Y (2004) Dynamics of soil microbial biomass C and soil fertility in cropland mulched with plastic film in a semiarid agro-ecosystem. Soil Biol Biochem 36:1893–1902
Li W, Wen X, Han J, Liu Y, Wu W, Liao Y (2017) Optimum ridge-to-furrow ratio in ridge-furrow mulching systems for improving water conservation in maize (Zea may L.) production. Environ Sci Pollut Res 24:23168–23179
Lin W, Liu W, Xue Q (2016) Spring maize yield, soil water use, and water use efficiency under plastic film and straw mulches in the Loess Plateau. Sci Rep 6:38995
Liu G, Zuo Y, Zhang Q, Yang L, Zhao E, Liang L, Tong YA (2018) Ridge-furrow with plastic film and straw mulch increase water availability and wheat production on the Loess Plateau. Sci Rep 8:6503
Martínez-García LB, Korthals G, Brussaard L, Jørgensen HB, De Deyn GB (2018) Organic management and cover crop species steer soil microbial community structure and functionality along with soil organic matter properties. Agric Ecosyst Environ 263:7–17
Mo F, Wang J, Xiong Y, Nguluu SN, Li F (2016) Ridge-furrow mulching system in semiarid Kenya: a promising solution to improve soil water availability and maize productivity. Eur J Agron 80:124–136
Mo F, Wang J, Li F, Nguluu SN, Ren H, Zhou H, Zhang J, Kariuki CW, Gicheru P, Kavagi L, Cheruiyot WK, Xiong Y (2017) Yield-phenology relations and water use efficiency of maize (Zea mays L.) in ridge-furrow mulching system in semiarid east African Plateau. Sci Rep 7:3260
Moreno MM, Moreno A (2008) Effect of different biodegradable and polyethylene mulches on soil properties and production in a tomato crop. Sci Hortic 116:256–263
Muñoz K, Buchmann C, Meyer M, Schmidt-Heydt M, Steinmetz Z, Diehl D, Thiele-Bruhn S, Schaumann GE (2017) Physicochemical and microbial soil quality indicators as affected by the agricultural management system in strawberry cultivation using straw or black polyethylene mulching. Appl Soil Ecol 113:36–44
Nelson DW, Sommers LE (1996) Total carbon, organic carbon, and organic matter: laboratory methods. In: Sparks DL et al (eds) Methods of soil analysis. Part 3, SSSA Book Ser. No. 5. ASA/SSSA, Madison, Wisconsin pp 961–1010
Pascual-Seva N, San Bautista A, López-Galarza S, Maroto JV, Pascual B (2018) Influence of different drip irrigation strategies on irrigation water use efficiency on chufa (Cyperus esculentus L. var. sativus Boeck.) crop. Agric Water Manag 208:406–413
Qin S, Zhang J, Dai H, Wang D, Li D (2014) Effect of ridge-furrow and plastic-mulching planting patterns on yield formation and water movement of potato in a semi-arid area. Agric Water Manag 131:87–94
Qin X, Li Y, Han Y, Hu Y, Li Y, Wen X, Liao Y, Siddique KHM (2018) Ridge-furrow mulching with black plastic film improves maize yield more than white plastic film in dry areas with adequate accumulated temperature. Agric For Meteorol 262:206–214
Ramakrishna A, Tam HM, Wani SP, Long TD (2006) Effect of mulch on soil temperature, moisture, weed infestation and yield of groundnut in northern Vietnam. Field Crop Res 95:115–125
Rathore R, Dowling DN, Forristal PD, Spink J, Cotter PD, Bulgarelli D, Germaine KJ (2017) Crop establishment practices are a driver of the plant microbiota in winter oilseed rape (Brassica napus). Front Microbiol 8:1489
Shen Y, Chen Y, Li S (2016) Microbial functional diversity, biomass and activity as affected by soil surface mulching in a semiarid farmland. PLoS One 11:e159144
Soong JL, Marañon-Jimenez S, Cotrufo MF, Boeckx P, Bodé S, Guenet B, Peñuelas J, Richter A, Stahl C, Verbruggen E, Janssens IA (2018) Soil microbial CNP and respiration responses to organic matter and nutrient additions: evidence from a tropical soil incubation. Soil Biol Biochem 122:141–149
Thankamani CK, Kandiannan K, Hamza S, Saji KV (2016) Effect of mulches on weed suppression and yield of ginger (Zingiber officinale Roscoe). Sci Hortic 207:125–130
Vance ED, Brookes PC, Jenkinson DS (1987) An extraction method for measuring soil microbial biomass C. Soil Biol Biochem 19:703–707
Wang Y, Xie Z, Malhi SS, Vera CL, Zhang Y, Wang J (2009) Effects of rainfall harvesting and mulching technologies on water use efficiency and crop yield in the semi-arid Loess Plateau, China. Agric Water Manag 96:374–382
Wang J, Lv S, Zhang M, Chen G, Zhu T, Zhang S, Teng Y, Christie P, Luo Y (2016a) Effects of plastic film residues on occurrence of phthalates and microbial activity in soils. Chemosphere 151:171–177
Wang YP, Li XG, Fu T, Wang L, Turner NC, Siddique KHM, Li F (2016b) Multi-site assessment of the effects of plastic-film mulch on the soil organic carbon balance in semiarid areas of China. Agric For Meteorol 228-229:42–51
Wu J, Xiao K, Zhao C, Yu A, Feng F, Li L, Chai Q (2018) Ridge-furrow cropping of maize reduces soil carbon emissions and enhances carbon use efficiency. Agric Ecosyst Environ 256:153–162
Yadav GS, Das A, Lal R, Babu S, Meena RS, Patil SB, Saha P, Datta M (2018) Conservation tillage and mulching effects on the adaptive capacity of direct-seeded upland rice (Oryza sativa L.) to alleviate weed and moisture stresses in the North Eastern Himalayan Region of India. Arch Agron Soil Sci 64:1254–1267
Yu Y, Tao H, Yao H, Zhao C (2018) Assessment of the effect of plastic mulching on soil respiration in the arid agricultural region of China under future climate scenarios. Agric For Meteorol 256-257:1–9
Zhang GS, Chan KY, Li GD, Huang GB (2008) Effect of straw and plastic film management under contrasting tillage practices on the physical properties of an erodible loess soil. Soil Tillage Res 98:113–119
Zhang F, Li M, Zhang W, Li F, Qi J (2017a) Ridge-furrow mulched with plastic film increases little in carbon dioxide efflux but much significant in biomass in a semiarid rainfed farming system. Agric For Meteorol 244-245:33–41
Zhang F, Zhang W, Li M, Zhang Y, Li F, Li C (2017b) Is crop biomass and soil carbon storage sustainable with long-term application of full plastic film mulching under future climate change? Agric Syst 150:67–77
Zhang Y, Han H, Zhang D, Li J, Gong X, Feng B, Xue Z, Yang P (2017c) Effects of ridging and mulching combined practices on proso millet growth and yield in semi-arid regions of China. Field Crop Res 213:65–74
Zhang Z, Liang S, Wang J, Zhang X, Mahamood M, Yu J, Zhang X, Liang A, Liang W (2018) Tillage and crop succession effects on soil microbial metabolic activity and carbon utilization in a clay loam soil. Eur J Soil Biol 88:97–104
Zhao H, Wang R, Ma B, Xiong Y, Qiang S, Wang C, Liu C, Li F (2014) Ridge-furrow with full plastic film mulching improves water use efficiency and tuber yields of potato in a semiarid rainfed ecosystem. Field Crop Res 161:137–148
Zhou L, Jin S, Liu C, Xiong Y, Si J, Li X, Gan Y, Li F (2012) Ridge-furrow and plastic-mulching tillage enhance maize-soil interactions: opportunities and challenges in a semiarid agroecosystem. Field Crop Res 126:181–188
Acknowledgments
This work was supported by the project of the Special Fund for Agro-scientific Research in the Public Interest (No. 201503127) and the National Natural Science Foundation of China (No. 31271673, No. 31700364, No. 31871583). The authors extremely thank the anonymous reviewers for their helpful comments and Dr. Mohsin Tanveer from School of Land and Food, University of Tasmania, Hobart, Australia, for improving the language of this manuscript.
Author information
Authors and Affiliations
Corresponding authors
Additional information
Responsible editor: Weixin Ding
Publisher’s note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Electronic supplementary material
ESM 1
(DOC 1495 kb)
Rights and permissions
About this article

Cite this article
Zhang, K., Xing, Y., Wang, G. et al. Ridge-furrow with film mulching practice ameliorates soil microbial metabolic activity and carbon utilization in rhizosphere soil of rapeseed (Brassica napus L.). J Soils Sediments 19, 2764–2776 (2019). https://doi.org/10.1007/s11368-019-02243-4
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s11368-019-02243-4