
Abstract
Purpose
The purposes of present study were to display the vertical distribution of soil organic carbon (SOC), nitrogen (N), and phosphorus (P) stoichiometry; identify the biogeographic characteristics of SOC, N, and P stoichiometry along an aridity gradient across the desert ecosystem of Hexi Corridor; and determine how biogeographic distribution patterns of SOC, N, and P stoichiometry are related to vegetation, soil texture, geography, and climate.
Materials and methods
We investigated the distribution and characteristics of SOC, N, and P stoichiometry based on samples collected from Hexi Corridor during 2011–2012 with total 400 plots of 80 sites. This region presents a precipitation gradient from about 250 mm in the east to less than 50 mm in the west. The measured variables included belowground and aboveground biomass, pH, bulk density, sand, clay, silt, SOC, N, and P contents. ANOVA analysis, reduced major axis, redundancy analysis, Person’s correlation, and regression analysis were used to analysis the variation of SOC, N, and P stoichiometry and related biogeographic factors.
Results and discussion
In present study, SOC, N, and P contents decreased significantly with increasing soil depth. C/N did not change significantly, while C/P and N/P decreased significantly. SOC and N, SOC and P, and N and P were well constrained within 0–100 cm. SOC, N, and P contents in 0–20 cm were higher than them in other studies. Vegetation, soil texture, climate, and geography could explain 91.6% of the total variance of soil stoichiometry. The impact of latitude and longitude on SOC, N, and P stoichiometry was mainly caused by the redistribution of precipitation, while the impact of altitude mainly resulted from the variation of temperature. With increasing aridity, SOC, N, and P contents and C/N/P ratios reduced consistently with inconsistent decrease rates.
Conclusions
Our results suggested that the interaction of vegetation structure, soil condition, and shortage of precipitation should be the main driver for the lower contents and much shallower distributions of SOC, N, and P of Hexi Corridor. The increasing aridity should be the critical factor that is responsible for the decrease of SOC, N, and P contents and C/N/P ratios. This study contributes to the understanding of soil stoichiometry in the desert ecosystem.
Similar content being viewed by others

Explore related subjects
Discover the latest articles, news and stories from top researchers in related subjects.Avoid common mistakes on your manuscript.
1 Introduction
Soil organic carbon (SOC), nitrogen (N), and phosphorus (P) play fundamental roles in the cycling of nutrient and the maintenance of ecosystem structure and function (Stevenson and Cole 1999; Griffiths et al. 2012). Through atmosphere deposition and fertilization, terrestrial ecosystems are undergoing unprecedented increases in nutrient element inputs, including N and P (Peñuelas et al. 2012; Tipping et al. 2014). The unbalanced inputs of N and P can critically affect ecological stoichiometry of soil and organisms, which would further alter ecosystem functioning (Sardans et al. 2012; Peñuelas et al. 2013). For instance, excess available N could alter plant nutrient stoichiometry and accelerate P cycles; however, P addition could result in large gaseous N loss in P-poor soils (Wang et al. 2010; Peñuelas et al. 2012; He and Dijkstra 2015). Although derived from different sources, soil N and P are closely related to SOC due to the constrained proportions of these elements required by organisms, which closely links with the variation of greenhouse gas emission and further contributes to the global climate change (Liu et al. 2013; Tang et al. 2017). Therefore, a clear understanding of SOC, N, and P contents and their distributions is necessary to evaluate current and potential ecosystem productivity and assess the potential climate change feedbacks (Jennings et al. 2009; Liu et al. 2013).
Robust linkages in stoichiometry of C, N, and P from molecular to global scales make it possible to track changes in the structure and nutrient cycling of ecosystems (Sterner and Elser 2002; Wardle et al. 2004; Zechmeister-Boltenstern et al. 2015). A considerable number of studies focused on soil C, N, and P stoichiometry and tried to decipher the related factors of soil nutrient stoichiometry (Delgado-Baquerizo et al. 2017; Luo et al. 2017; Yu et al. 2017). For example, a well-constrained C/N ratio in 0–10 cm organic-rich soil was reported at regional and global scales (Cleveland and Liptzin 2007; Tian et al. 2010). However, due to biogeographic heterogeneity of glacial history, climate, geologic parent material, topography, and biotic diversity, stoichiometry of C, N, and P also exhibits different scenarios, making it difficult to accurately evaluate the biogeography of soil stoichiometry (Jenny 1941; Taylor and Townsend 2010; Tian et al. 2010; Tang et al. 2017). Ratios of C/P and N/P in China are lower than global average value, and ratio of C/N/P is different between China and global scale, depending on the variations of vegetation types, soil types, and biogeographic climate (Cleveland and Liptzin 2007; Tian et al. 2010; Yu et al. 2017). It is also predicted that future climate would be much dryer and warmer in drylands worldwide, resulting in altered water availability and temperature and posing great threaten on the balance of C, N, and P cycles and the key services provided by dryland ecosystems (Gao and Giorgi 2008; Dai 2013; Delgado-Baquerizo et al. 2013; Feng and Fu 2013; He and Dijkstra 2014; Jiao et al. 2016; Berdugo et al. 2017). In an extremely arid environment, SOC and N decreased with increasing aridity, but the inorganic P increased (Delgado-Baquerizo et al. 2013). In more arid and warmer sites, SOC, N, and P had lower values, while ratios of C/N, C/P, and N/P showed different varying trends (Jiao et al. 2016). These results indicated the decoupling of soil C, N, and P stoichiometry with aridity gradient in arid ecosystems. However, based on these inconsistent results, it is difficult to draw a universal conclusion. Thus, it is crucial to evaluate the responses of soil stoichiometry to environmental changes at different biogeographic scales (Zhang et al. 2013; O'Rourke et al. 2015; Stockmann et al. 2015).
Hexi Corridor, locating at the northwestern China, is defined as the transitional zones between desert and grassland or between desert and oasis and is crucial for maintaining the stable ecological system and local productivity. The desert ecosystem of Hexi Corridor is characterized by small shrubs or subshrubs with shallow roots, thick soil layer (from 0.2 to 1.5 m), and a large amount of gravel (0.5–6 cm in diameter) with increasing soil depth (Wang et al. 2013, 2014). The mean annual precipitation in this area, from east to west, ranges from about 250 mm to less than 50 mm, providing a natural gradient for studying the patterns of soil C, N, and P stoichiometry and the potential response of soil stoichiometry to the ongoing global climate changes. However, due to the insufficient field observations and large biogeographic heterogeneity, the research on soil stoichiometry in the desert ecosystem of Hexi Corridor remains an important and scientific gap. Meanwhile, the relationship between biogeographic factors and the distribution of soil stoichiometry is still poorly understood.
In present study, we hypothesized that SOC, N, and P stoichiometry across the desert ecosystem of Hexi Corridor is mediated by various processes at regional scale. The aims of our research were to clarify the vertical and biogeographic distributions of SOC, N, and P stoichiometry across the desert ecosystem of Hexi Corridor and to identify the relationship between biogeographic distribution patterns of SOC, N, and P stoichiometry and vegetation, soil texture, geography, and climate. The information obtained from this research would contribute to a better understanding of soil stoichiometry characteristics of desert ecosystem and facilitate the management of desert ecosystem under global climatic changes.
2 Materials and methods
2.1 Study area
The study area locates at the central region of Hexi Corridor in Gansu Province, northwestern China (94° 21′–102° 18′ E, 38° 23′–41° 45′ N). Altitude ranges from 1190 to 2560 m. Mean annual temperature (MAT) ranges from 5 to 9 °C. Mean annual precipitation (MAP) varies from about 250 mm to less than 50 mm from east to west with 70–80% of falling during June to August, while the aridity index ranges from 1.3 to 17.5. Aridosols and entisols are the main soil types with pH ranges from 8.3 to 9.3. Since 2000, grazing has been prohibited in the research area. The predominant species are small shrubs and subshrubs, including Asterothamnus centraliasiaticus, Reaumuria songarica, Salsola passerina, and Sympegma regelii, and ephemeral plant species such as Suaeda glauca, Bassia dasyphylla, and Artemisia scoparia (Wang et al. 2014).
2.2 Sampling and measurements
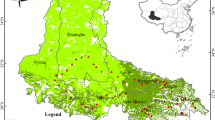
Soil samples were collected in August of 2011 and 2012. Eighty sites were selected across the study area, which represent the vegetation structure, soil order, and precipitation gradient in the desert ecosystem of Hexi Corridor (Fig. 1). Global positioning system (GARMIN eTrex Vista, Taiwan, China) was used to record geographical coordinate and elevation of each site. Based on the records of 25 weather stations across the Hexi Corridor, spatial interpolation was used to obtain detail data of MAT and MAP in the whole area (Zhang et al. 2017). At each site, a 100 m line transect was set up. Along each transect, ten plots marked with 1 to 10 (1 m × 1 m) were established. Five odd-numbered plots in each site were selected for soil sampling, and data from these plots were used to calculate average value of each site. At each plot, soil samples were randomly excavated at seven depths with three replicates: 0–5, 5–10, 10–20, 20–30, 30–50, 50–70, and 70–100 cm. Soil samples in each soil depth were mixed evenly. Samples for soil bulk density of each soil layer were obtained using a cutting ring (volume of 100 cm3) and oven-dried for 24 h at 105 °C. Samples for measuring SOC, N, and P contents were air-dried, grounded, and then sieved using a 100-mesh sieve.

Distribution of sampling sites in desert ecosystem of Hexi Corridor, northwestern China
SOC was determined by the potassium dichromate volumetric method in conjunction with external heating method. N was measured using a vario MACRO CUBE (Elementar Analysensysteme, Germany). P was measured by the colorimetry method after H2SO4-H2O2 digestion.
2.3 Statistical analyses
The degree of aridity was characterized by precipitation and temperature according to the study by de Martonne:
where IDM is the de Martonne aridity index; P is the mean annual precipitation in millimeters; and T is the mean annual temperature in degree (de Martonne 1926; Tabari et al. 2014). The higher the IDM value, the lower the degree of aridity would be.
To examine the correlations of SOC, N, and P, the reduced major axis (RMA) analysis was performed with SMART package (http://www.bio.mq.edu.au/ecology/SMART). In RMA analysis, we used a scaling approach (Y = aXb) to examine the scaling relations among SOC, N, and P. The redundancy analysis (RDA), Person’s correlation, and regression analysis were used to explore the influence of plant biomass, geography, climate, and soil texture on soil C/N/P. The soil order was classified into four types: Sierozems, Gray desert soil, Gray brown soil, and Desert eolian sand soil. ANOVA was used to test effects of soil depths and soil orders on SOC, N, P, and their ratios. All data satisfied restrictions of homogeneity of variance (P > 0.05). The above analyses were conducted using SPSS 18.0 (IBM, Chicago, IL, USA), Canoco 4.5 (Microcomputer Power, Ithaca, NY, USA) and Origin 9.0 (OriginLab, USA).
3 Results
3.1 Distribution of SOC, N, and P stoichiometry
As is shown in Table 1, the maximum contents of SOC, N, and P were in 0–5 cm. With increasing soil depth, SOC, N, and P decreased significantly; however, there were no significant differences of SOC and P within 0–20 cm soil layer and of N within 0–30 cm soil layer. Moreover, the contents of SOC, N, and P did not show significant differences among 30–50, 50–70, and 70–100 cm. With increasing soil depth, ranges of SOC, N, and P contents were 182.59–322.03, 22.80–37.12, and 13.26–16.59 mmol kg−1, respectively. The ratio of C/N showed no significant difference with ranges from 7.96 to 8.71. Ratios of C/P and N/P exhibited no significant differences within 0–50 cm with ranges from 12.16 to 17.91 and from 1.48 to 2.09, respectively. Across the seven depths, C/N/P changed from 18:2:1 to 12:2:1.
In 0–30 cm, proportions of SOC, N, and P were 79, 78, and 76% of the whole profile (0–100 cm), respectively. Contents of SOC, N, and P were 254.94 ± 22.72, 31.18 ± 2.51, 14.81 ± 0.62 mmol kg−1, and the coefficients of variations were 0.80, 0.72, and 0.38, respectively. Ratios of C/N, C/P, and N/P were 8.19 ± 0.23, 16.25 ± 0.88, and 2.04 ± 0.10, and the coefficients of variations were 0.25, 0.48, and 0.43, respectively. Ratio of C/N/P was 16:2:1.
As is shown in Table 2, SOC, N, and P contents showed maximum values in Sierozems; however, there were no significant differences among Gray desert soil, Gray brown soil, and Desert eolian sand soil. Desert eolian sand soil had maximum ratio of C/N, while the ratio of C/N did not display significant differences in the other three soil orders. Ratios of C/P and N/P in different soil orders were similar with SOC, N, and P contents. C/N/P in Sierozems, Gray desert soil, Gray brown soil, and Desert eolian sand soil were 9:1:1, 7:1:1, 6:1:1, and 6:1:1, respectively.
3.2 Relations of SOC, N, and P
In 0–30 cm, Pearson’s correlation coefficients of SOC and N, SOC and P, and N and P were 0.97, 0.73, and 0.75, respectively (Table 3). Coefficients were 0.97, 0.73, and 0.74 for SOC and N, SOC and P, and N and P in 0–10 cm, respectively, and 0.96, 0.74, 0.76 in 0–100 cm for them, respectively. The RMA slopes for SOC and N, SOC and P, N and P were 0.90, 0.60, 0.67, respectively, which were significantly different from 1 (P < 0.05) (Table 4).
3.3 Factors related to SOC, N, and P stoichiometry
Results of RDA are shown in Fig. 2 and Table 5 as well as Tables S1 and S2 in the Electronic Supplementary Material. The eigenvalues of soil stoichiometry and biogeographic factors for the first and second axes were 0.92 and 0, respectively (Table 5). The first axis explained 91.6% of the variance of soil stoichiometry and 99.9% of the variance of soil stoichiometry and biogeographic factors relations. However, the second axis only explained 0.1% of the variance of soil stoichiometry and explained 0.1% of the variance of soil stoichiometry and biogeographic factors relations. SOC, N, P, C/P, and N/P were located on the right of the first axis (Fig. 2). According to the arrow length, for the 12 factors, the effects of silt, sand, BB (belowground biomass), MAT, altitude, AB (aboveground biomass), MAP, clay, latitude, BD, longitude, and pH on soil stoichiometry weakened orderly. Among them, silt, clay, MAP, AB, BB, longitude, altitude, and MAP that are located on the right of the first axis indicated the positive effects on SOC, N, P, C/P and N/P (Tables S1 and S2, Electronic Supplementary Material). Correlation analysis and regression analysis showed that soil C/N was mainly affected by MAT and biomass.

RDA biplot of biogeographic factors and soil stoichiometry (AB and BB represent aboveground biomass and belowground biomass, respectively)
The regression analysis showed that IDM had significant positive influence on soil stoichiometry (P < 0.001) (Fig. 3). The regression coefficients (R2) of IDM and SOC, N, C/P, P, N/P, and C/N were 0.42, 0.33, 0.29, 0.24, 0.13, and 0.09, respectively.

Responses of SOC, N, and P stoichiometry on aridity degree
4 Discussion
4.1 Distribution of SOC, N, and P stoichiometry in the desert ecosystem
Nutrient stoichiometry is essential to nutrient cycling, litter decomposition, and habitat variability, which regulate the ecosystem functions and services (Sterner and Elser 2002; Zechmeister-Boltenstern et al. 2015). In present study, contents of SOC and N showed significant decrease trend with increasing soil depth and 80% of them concentrated in 0–30 cm. This verified that soil surface layer (0–30 cm) interacts directly with atmosphere and is the most sensitive layer to land-use changes (Batjes 1996; Wang et al. 2014; Cao et al. 2017). In 0–20 cm, the proportion of SOC of Hexi Corridor was 62% and higher than it in global shrublands (33%), global grasslands (42%), and Tibetan grasslands (49%) (Jobbágy and Jackson 2000; Yang et al. 2010). The proportion of N (61%) in 0–20 cm of Hexi Corridor was higher than it in Tibetan grasslands (43%), and the proportion of P of Hexi Corridor accounted for 60% (Jobbágy and Jackson 2001). Moreover, contents of SOC at China national scale with 133 samples (2047 mmol kg−1) and in China temperate desert with 319 samples (775 mmol kg−1) were extremely higher than it in present study (Tian et al. 2010). The contents of N at China national scale and in temperate desert were about four and three times than it in present study, while the content of P in present study showed less difference with them (Tian et al. 2010). These results indicated the much shallower distributions and lower contents of SOC, N, and P in the desert ecosystem of Hexi Corridor.
The above results may be ascribed to the complicated interactive relationship of climate, soil conditions, and vegetation. Hexi Corridor is typically featured with limited precipitation, thinner soil layer thickness, and greater gravel content, especially below 30 cm of soil layer (Wang et al. 2014). As a result, the dominant plants in this area are small shrubs or semi-shrubs with lower root biomass (58.01 g m−2) and shallower distribution (more than 90% centered in 0–30 cm) of root (Wang et al. 2014). This typical phenomenon, together with the shortage of precipitation that limited the leaching of N and P downward to soil, created the lower vegetation production and limited SOC inputs and modified the redistribution of soil nutrients directly (Jobbágy and Jackson 2000; Jobbágy and Jackson 2001; He et al. 2014; He and Dijkstra 2014). However, even though soil P in present study showed a decrease trend with increasing depth, the less variation (13.26–16.59 mmol kg−1) of P was consistent with previous results conducted at national and regional scales (Tian et al. 2010; He et al. 2014; Cao et al. 2017). The main reason accounted for this is the relatively stable vertical distribution of soil P due to the weathering of parent material that provides the major source of soil P, and soil P located at the bottom of soil profile should be conveyed by deep-rooted plants from deeper soil to the surface (Jobbágy and Jackson 2001; He et al. 2014; Cao et al. 2017).
The non-significant difference of C/N with increasing depth indicated that the variation of SOC and N was well coordinated (Cleveland and Liptzin 2007; Tian et al. 2010). Actually, this is not surprising because plants are the major source of C and N to soil (Griffiths et al. 2012). Unlike the ratio of C/N, ratios of C/P and N/P that decreased with increasing soil depth were mainly controlled by the relatively stable soil P across soil depth. Moreover, because of the lower contents of SOC, N, and P, ratios of soil C/N, C/P, and N/P were lower than other studies; meanwhile, the ratio of C/N/P differs significantly with the Redfield ratio (106:16:1) (Cleveland and Liptzin 2007; Tian et al. 2010).
For a specific soil order, soil chemical properties, such as SOC, N, and P, can reflect the combined effects by climate, parent material, organisms, and time (Tian et al. 2010). Contents of SOC, N, and P in present study have no significant differences among Gray desert soil, Gray brown soil, and Desert eolian sand soil with C/N/P changed from 9:1:1 to 6:1:1, indicating the slower soil development in the desert ecosystem of Hexi Corridor.
4.2 Relations of SOC, N, and P in the desert ecosystem
Based on 400 soil profiles across the desert ecosystem, we found the well-constrained relation of SOC and N in 0–10, 0–30, and 0–100 cm with high correlation coefficients of 0.97, 0.97, and 0.96, respectively. It disaccorded with previous study that only verified the well-constrained relation of SOC and N in organic rich soil layer (0–10 cm) (Tian et al. 2010). In present study, SOC and P, N, and P also showed relatively constrained relations in 0–10, 0–30, and 0–100 cm. It suggested that, in this arid region, even though C/N/P ratios were quite lower than them at national and global scale, the well-constrained relations among C, N, and P existed not only in soil surface but also in the deeper soil layer, at least up to 100 cm (Cleveland and Liptzin 2007; Tian et al. 2010). Thus, it is necessary to evaluate the distribution of C, N, and P stoichiometry in different scales, especially in the regional scale with the first-hand samples. In addition, despite the vertically well-constrained relations of SOC, N, and P, the horizontally allometric scaling relationships of SOC, N, and P showed increase rate in this order: SOC > N > P.
4.3 Factors related to soil C, N, and P stoichiometry in the desert ecosystem
RDA results verified that SOC, N, and P stoichiometry was significantly related to plants, soil texture, geography, and climate (Burke et al. 1989; Batjes 1996; Jobbágy and Jackson 2000). Similar to other regions, SOC, N, and P stoichiometry of Hexi Corridor also showed positive correlations with silt and clay, due to the acceleration effects of fine soil particles on forming aggregates, improving soil quality, and stabilizing SOC, N, and P (Schimel et al. 1994; Yang et al. 2008; Su et al. 2010). Moreover, the positive relationships between biomass and SOC, N, and P stoichiometry of Hexi Corridor suggested that biomass, especially for the belowground biomass, is one of the determine factors on SOC, N, and P stoichiometry.
In present study, SOC, N, P, C/P, and N/P showed significant variations along the narrow longitude and latitude gradients. This result might be largely caused by the replacement of precipitation along longitude and latitude gradients (Table S2, Electronic Supplementary Material). With a wider range of altitude gradient, the increase of SOC, N, P, C/P, and N/P might be ascribed to the decreasing MAT (R2 = 0.53) than the increasing MAP (R2 = 0.12) (Fig. S1, Electronic Supplementary Material). This may be attributed to the increasing temperature, especially in the arid region, which would result in a considerable decline in water use efficiency by increasing evapotranspiration, low biomass production, and low nutrient contents (Schimel et al. 1994; Jobbágy and Jackson 2000; Canadell et al. 2007). These results suggested that, although the biogeochemical cycles of present study region are charactered with limited precipitation, its effects became weaken along altitude gradient.
In present study, SOC, N, and P contents and C/N/P ratios consistently increased with increasing IDM. It could be contributed by the increasing soil water availability, vegetation biomass, and biological weathering that caused by the decreasing aridity degree. Aridity degree has direct or indirect effects on relevant biogeochemical processes of SOC, N, and P cycles (Bray 1997; Farooq et al. 2012; Sardans and Peñuelas 2012; Ledger et al. 2013). The consistent variations of SOC, N, and P contents and C/N/P ratios with increasing IDM were disaccord with the study conducted in all continents except Antarctica and the study of Inner Mongolia (Delgado-Baquerizo et al. 2013; Jiao et al. 2016). These discrepancies implied that with increasing aridity, SOC, N, and P contents and C/N/P ratios in different regions would behave differently. In addition, it suggested that the aridity may be the critical factor to the inconsistent variations of these three elements. However, because of the complex interactions of factors, it still needs more quantitative studies for such specific region.
5 Conclusions
In conclusion, due to the complex effects of vegetation, soil texture, geography, and climate, SOC, N, and P in the desert ecosystem of Hexi Corridor had lower contents and much shallower distributions. Moreover, the well-constrained SOC, N, and P occurred from soil surface to the deeper soil layer. The main climate factor for soil stoichiometry variation along latitude and longitude gradients was precipitation, while temperature was responsible for its variation along altitude gradient. Our results found that the increasing aridity could reduce SOC, N, and P contents and C/N/P ratios and suggested that aridity should be the critical factor to determine the inconsistent variations of SOC, N, and P that primarily link to biological processes.
References
Batjes NH (1996) Total carbon and nitrogen in the soils of the world. Eur J Soil Sci 47:151–163
Berdugo M, Kéff S, Soliveres S, Maestre FT (2017) Plant spatial patterns identify alternative ecosystem multifunctionality states in global drylands. Nat Ecol Evol 1:0003
Bray EA (1997) Plant responses to water deficit. Trends Plant Sci 2:48–54
Burke IC, Yonker CM, Parton WJ, Cole CV, Flach K, Schimel DS (1989) Texture, climate, and cultivation effects on soil organic matter content in U.S. grassland soils. Soil Sci Soc Am J 53:800–815
Canadell JG, Kirschbaum MUF, Kurz WA, Sanz MJ, Schlamadinger B, Yamagata Y (2007) Factoring out natural and indirect human effects on terrestrial carbon sources and sinks. Environ Sci Pol 10:370–384
Cao Y, Zhang P, Chen YM (2017) Soil C:N:P stoichiometry in plantations of N-fixing black locust and indigenous pine, and secondary oak forests in Northwest China. J Soils Sediments 2:1–12
Cleveland CC, Liptzin D (2007) C:N:P stoichiometry in soil: is there a “Redfield ratio” for the microbial biomass? Biogeochemistry 85:235–252
Dai A (2013) Increasing drought under global warming in observations and models. Nat Clim Change 3:52–58
Delgado-Baquerizo M, Maestre FT, Gallardo A, Bowker MA, Wallenstein MD, Quero JL, Ochoa V, Gozalo B, García-Gómez M, Soliveres S, García-Palacios P, Berdugo M, Valencia E, Escolar C, Arredondo T, Barraza-Zepeda C, Bran D, Carreira JA, Chaieb M, Conceição AA, Derak M, Eldridge DJ, Escudero A, Espinosa CI, Gaitán J, Gatica MG, Gómez-González S, Guzman E, Gutiérrez JR, Florentino A, Hepper E, Hernández RM, Huber-Sannwald E, Jankju M, Liu J, Mau RL, Miriti M, Monerris J, Naseri K, Noumi Z, Polo V, Prina A, Pucheta E, Ramírez E, Ramírez-Collantes DA, Romão R, Tighe M, Torres D, Torres-Díaz C, Ungar ED, Val J, Wamiti W, Wang D, Zaady E (2013) Decoupling of soil nutrient cycles as a function of aridity in global drylands. Nature 502:672–676
Delgado-Baquerizo M, Eldridge DJ, Maestre FT, Ochoa V, Gozalo B, Reich PB, Singh BK (2017) Aridity decouples C:N:P stoichiometry across multiple trophic levels in terrestrial ecosystems. Ecosystems 21:459–468
Farooq M, Hussain M, Wahid A, Siddique KHM (2012) Drought stress in plant: an overview. In: Aroca R (ed) Plant responses to drought stress. Springer, Berlin, pp 1–33
Feng S, Fu Q (2013) Expansion of global drylands under a warming climate. Atmos Chem Phys Discuss 13:14637–14665
Gao XJ, Giorgi F (2008) Increased aridity in the Mediterranean region under greenhouse gas forcing estimated from high resolution simulations with a regional climate model. Glob Planet Chang 62:195–209
Griffiths BS, Spilles A, Bonkowski M (2012) C:N:P stoichiometry and nutrient limitation of the soil microbial biomass in a grazed grassland site under experimental P limitation or excess. Ecol Process 1:6
He MZ, Dijkstra FA (2014) Drought effect on plant nitrogen and phosphorus: a meta-analysis. New Phytol 204:924–931
He MZ, Dijkstra FA (2015) Phosphorus addition enhances loss of nitrogen in a phosphorus-poor soil. Soil Biol Biochem 82:99–106
He MZ, Dijkstra FA, Zhang K, Li XR, Tan HJ, Gao YH, Li G (2014) Leaf nitrogen and phosphorus of temperate desert plants in response to climate and soil nutrient availability. Sci Rep 4:6932
Jennings E, Allott N, Pierson DC, Schneiderman EM, Lenihan D, Samuelsson P, Taylor D (2009) Impact of climate change on phosphorus loading form a grassland catchment: implication for future management. Water Res 43:4316–4326
Jenny H (1941) Factors of soil formation. McGraw-Hill, New York
Jiao F, Shi XR, Han FP, Yuan ZY (2016) Increasing aridity, temperature and soil pH induce soil C-N-P imbalance in grasslands. Sci Rep 6:19601
Jobbágy EG, Jackson RB (2000) The vertical distribution of soil organic carbon and its relation to climate and vegetation. Ecol Appl 10:423–436
Jobbágy EG, Jackson RB (2001) The distribution of soil nutrients with depth: global patterns and the imprint of plants. Biogeochemistry 53:51–77
Ledger ME, Brown LE, Edwards FK, Milner AM, Woodward G (2013) Drought alters the structure and functioning of complex food webs. Nat Clim Chang 3:223–227
Liu ZP, Shao MA, Wang YQ (2013) Spatial patterns of soil total nitrogen and soil total phosphorus across the entire loess plateau region of China. Geoderma 197-198:67–78
Luo WT, Li MH, Sardans J, Lü XT, Wang C, Penuelas J, Wang ZW, Han XG, Jiang Y (2017) Carbon and nitrogen allocation shifts in plants and soils along aridity and fertility gradients in grasslands of China. Ecol Evol 7:6927–6934
de Martonne E (1926) Traité de Géographie Physique, 3 tomes. Paris
O'Rourke SM, Angers DA, Holden NM, Mcbratney AB (2015) Soil organic carbon across scales. Glob Change Biol 21:3561–3574
Peñuelas J, Poulter B, Sardans J, Ciais P, van der Velde M, Bopp L, Boucher O, Godderis Y, Hinsinger P, Llusia J, Nardin E, Vicca S, Obersteiner M, Janssens IA (2013) Human-induced nitrogen-phosphorus imbalances alter natural and managed ecosystems across the globe. Nat Commun 4:2934
Peñuelas J, Sardans J, Rivas-ubach A, Janssens IA (2012) The human-induced imbalance between C, N and P in Earth’s life system. Glob Chang Biol 18:3–6
Sardans J, Peñuelas J (2012) The role of plants in the effects of global change on nutrient availability and stoichiometry in the plant-soil system. Plant Physiol 160:1741–1761
Sardans J, Rivas-Ubach A, Peñuelas J (2012) The elemental stoichiometry of aquatic and terrestrial ecosystems and its relationships with organismic lifestyle and ecosystem structure and function: a review and perspectives. Biogeochemistry 111:1–39
Schimel DS, Braswell BH, Holland EA, McKeown R, Ojima DS, Painter TH, Parton WJ, Townsend AR (1994) Climatic, edaphic, and biotic controls over storage and turnover of carbon in soils. Glob Biogeochem Cycles 8:279–293
Sterner RW, Elser JJ (2002) Ecological stoichiometry: the biology of elements from molecules to the biosphere. Princeton University Press, Princeton
Stevenson FJS, Cole MA (1999) Cycles of soils: carbon, nitrogen, phosphorus, sulfur, micronutrients, 2nd edn. Wiley, New York
Stockmann U, Padarian J, McBratney A, Minasny B, de Brogniez D, Montanarella L et al (2015) Global soil organic carbon assessment. Glob Food Secur-Agr 6:9–16
Su YZ, Wang XF, Yang R, Lee J (2010) Effects of sandy desertified land rehabilitation on soil carbon sequestration and aggregation in an arid region in China. J Environ Manag 91:2109–2116
Tabari H, Hosseinzadeh Talaee P, Mousavi Nadoushani SS, Willems P, Marchetto A (2014) A survey of temperature and precipitation based aridity indices in Iran. Quarter Int 345:158–166
Tang XL, Xia MP, Guan FY, Fan SH (2017) Spatial distribution of soil nitrogen, phosphorus and potassium stocks in Moso bamboo forests in subtropical China. Forests 7:267
Taylor PG, Townsend AR (2010) Stoichiometric control of organic carbon-nitrate relationships from soils to the sea. Nature 464:1178–1181
Tian HQ, Chen GS, Zhang C, Melillo JM, Hall CAS (2010) Pattern and variation of C:N:P ratios in China’s soils: a synthesis of observational data. Plant Soil 98:139–151
Tipping E, Benham S, Boyle JF, Crow P, Davies J, Fischer U, Guyatt H, Helliwell R, Jackson-Blake L, Lawlor AJ, Monteith DT, Rowe EC, Toberman H (2014) Atmospheric deposition of phosphorus to land and fresh-water. Environ Sci-Proc Imp 16:1608–1617
Wang Y, Law R, Park B (2010) A global model of carbon, nitrogen and phosphorus cycles for the terrestrial biosphere. Biogeosciences 7:2261–2282
Wang M, Su YZ, Yang R, Yang X (2013) Allocation patterns of above- and below-ground biomass in desert grassland in the middle reaches of Heihe River, Gansu Province, China. Chin J Plant Ecol 37:209–219 (in Chinese)
Wang M, Su YZ, Yang X (2014) Spatial distribution of soil organic carbon and its influencing factors in desert grasslands of the Hexi Corridor, Northwest China. PLoS One 9:e94652
Wardle DA, Walker LR, Bardgett RD (2004) Ecosystem properties and forest decline in contrasting long-term chronosequences. Science 305:509–513
Yang YH, Fang JY, Tang YH, Ji CJ, Zheng CY, He JS, Zhu B (2008) Storage, patterns and controls of soil organic carbon in the Tibetan grasslands. Glob Change Biol 14:1592–1599
Yang YH, Fang JY, Guo DL, Ji CJ, Ma WH (2010) Vertical patterns of soil carbon, nitrogen and carbon: nitrogen stoichiometry in Tibetan grasslands. Biogeosci Discuss 7:1–24
Yu ZP, Wang MH, Huang ZQ, Lin TC, Vadeboncoeur MA, Searle EB, Chen HYH (2017) Temporal changes in soil C-N-P stoichiometry over the past 60 years across subtropical China. Glob Change Biol 24:1308–1320
Zechmeister-Boltenstern S, Keiblinger KM, Mooshammer M, Peñuelas J, Richter A, Sardans J, Wanek W (2015) The application of ecological stoichiometry to plant-microbial-soil organic matter transformations. Ecol Monogr 85:133–155
Zhang ZS, Song XL, Lu XG, Xue ZS (2013) Ecological stoichiometry of carbon, nitrogen, and phosphorus in estuarine wetland soils: influences of vegetation coverage, plant communities, geomorphology, and seawalls. J Soils Sediments 13:1043–1051
Zhang K, Su YZ, Yang R (2017) Biomass and nutrient allocation strategies in a desert ecosystem in the Hexi Corridor, northwest China. J Plant Res 130:699–708
Acknowledgments
We sincerely appreciate Dr. Yongle Chen from Shenzhen University and Dr. Jingjing Du from Zhengzhou University of Light Industry for checking the English language of our manuscript.
Funding
This study was supported by the National Key Research and Development Program of China (No. 2017YFC0504304).
Author information
Authors and Affiliations
Corresponding author
Additional information
Responsible editor: Yongfu Li
Electronic supplementary material
ESM 1
(DOCX 196 kb)
Rights and permissions
About this article

Cite this article
Zhang, K., Su, Y. & Yang, R. Variation of soil organic carbon, nitrogen, and phosphorus stoichiometry and biogeographic factors across the desert ecosystem of Hexi Corridor, northwestern China. J Soils Sediments 19, 49–57 (2019). https://doi.org/10.1007/s11368-018-2007-2
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s11368-018-2007-2