
Abstract
Purpose
The present study focuses on a compositional characterization of the humic acid (HA) fraction of several wetland soils using solid-state 13C NMR spectroscopy. The data were analysed using the molecular mixing model (MMM), based on an empirical approach by Nelson and Baldock. The compositional data from HAs obtained with this model were used to obtain a wider assessment of the process of humification from comparison of total soil wetland organic matter composition and HA composition.
Materials and methods
Twenty samples of humic acids (HAs) isolated from a Mediterranean semiarid wetland (‘Tablas de Daimiel’, central Spain) were studied using elemental analysis and cross polarization magic angle spinning (CPMAS)13C nuclear magnetic resonance (NMR) spectroscopy. The NMR data were analysed with the molecular mixing model (MMM) considering up to six generic components (carbohydrate, protein, lignin, lipid, char and ‘carbonyl’). HAs are considered a conceptual mixture of these model components, and the MMM determines the proportions of the characteristic biomolecules contributing to HA composition.
Results and discussion
The composition of the HAs under study depends on local factors such as site vegetation and occurrence of fire. Correlations between the proportions of the six generic components and further comparison with those determined for the unfractionated OM (whole sample, WS), gave information on HA origin and humification mechanisms. In particular, the proportions of char and carbohydrate (R 2 0.637) and contents of lignin and protein (R 2 0.471) in the HAs were negatively correlated (P < 0.05). Significant correlations (R 2 0.439) also existed for char contents in whole sample (WS) compared to HA, and for carbohydrates in WS compared to HA (R 2 0.558). Char proportion grew in HA with respect to the WS, and carbohydrates dropped to a half on average in HA compared to WS.
Conclusions
Two different humification mechanisms could be identified for no-fire and fire areas. In the former, HA-char was preserved selectively from char in the sample, whereas in the latter, char was newly formed by fire effect.
Similar content being viewed by others
Explore related subjects
Discover the latest articles, news and stories from top researchers in related subjects.Avoid common mistakes on your manuscript.
1 Introduction
Humification is assumed to consist of widespread biochemical and abiotic diagenetic reactions leading to recalcitrant, very complex macromolecular compounds (Aiken et al. 1985; Almendros 2008). The resulting humic substances (HS) can originate from alteration of pre-existing biomacromolecules but also from abiotic or enzymatic condensation of soluble biogenic precursors (Almendros 2008). Both processes result in the accumulation of material which is more complex than the corresponding precursors and comparatively much more resistant to microbial attack (Almendros 2008). However, it has also been pointed out, that the recalcitrance of natural organic matter (OM), including HS, depends on association with soil minerals and may not be an intrinsic characteristic of the substrate (Kleber 2010). In any case, humification can be seen as a key step within the wetland carbon cycle, strongly contributing to the important role of wetlands as carbon sinks (Bridgham et al. 2006).
The HS are generally divided according to their solubility in aqueous solution (Hayes et al. 1989). The fraction that is only soluble under alkaline conditions is called the humic acid (HA) fraction. HAs are polydisperse, heterogeneous substances. There are several conceptions concerning their nature and composition: according to an early view, they can be seen as a system of micelles of a polymeric nature, with high average molecular masses (20,000–50,000 Da; Aiken et al. 1985). Some authors have considered that HAs do not exist in soil as a definite entity, being mere mixtures of plant and microbial remains and their degradation products (Burdon 2001). As a matter of fact, specific chemical structures belonging to HS have not been identified (Kelleher and Simpson 2006; Lehmann et al. 2008); an extended discussion on this topic is given by Kleber and Johnson (2010). The polymeric character of HS is also a matter of discussion: some authors have introduced terminological insights which consider that HS are supramolecular associations of self-assembling heterogeneous and relatively small molecules (Piccolo 2001). This view is shared by others (Sutton and Sposito 2005), who consider that HS would contain tightly bonded biomolecules or recognizable fragments of them.
The present study focuses on a compositional characterization of the HA fraction using solid-state 13C NMR spectroscopy. The data (from twenty samples of HAs) were analysed using the molecular mixing model (MMM), based on an empirical approach by Nelson and Baldock (2005) in an attempt to describe the molecular composition of natural OM from quantitative data obtained from 13C nuclear magnetic resonance (NMR) spectroscopy. These authors showed that detrital OM, derived from different sources, can be described in terms of six generic components (carbohydrate, protein, lignin, lipid, ‘pure carbonyl’ and char). The contribution of each to the total OM can be inferred from the respective 13C NMR spectra by way of a linear combination of spectra of the six generic organic components. The approach of Nelson and Baldock (2005) allows a more specific assignment of the C-groups assigned from NMR to various compound classes. In particular, a better discrimination between sources of aromatic C may help differentiate pyrogenic C forms, i.e. char from plant-derived aromatic components. This improves the analysis of the origin of HAs from different precursor macromolecular entities, which is especially appropriate for wetland soils (in most cases peat or histosols) where selective preservation of macromolecular biomass constituents is a classically recognized factor in the C sequestration in these ecosystems.
The compositional data from HAs were used to obtain a wider assessment of the process of humification taking advantage of comparisons between total soil wetland OM composition and HA composition. To our knowledge, although MMM has been applied to HAs only in the study by Buurman et al. 2009, there is no previous research on MMM applied to HAs from wetland soils.
2 Materials and methods
2.1 Study site
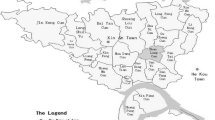
The Tablas de Daimiel is a freshwater wetland in Central Spain on the River Guadiana drainage basin (39° 08′ N, 3°43′ W; Fig. 1). Its detailed description and environmental history were recently reported by Sánchez-Carrillo and Angeler (2010) and Rodríguez-Murillo et al. (2011).

Location of Tablas de Daimiel wetland and sampling points
2.2 Sample preparation and HA extraction
Samples (20) from the upper 20 to 30 cm of the wetland soils from Las Tablas de Daimiel, corresponding to different local environments in the wetland, were collected in March 2007 (Fig. 1). In the case of organic soils or sediments, sampling was carried out collecting 30 cm of the uppermost horizon. Only in sandy sediments with low content of organic C the material was collected in the organc horizon, i.e. 4–5 cm thickness topsoil layer. The samples were air-dried, ground and sieved to pass a 2-mm sieve. The HAs were extracted following a standard procedure described by (Duchaufour and Jacquin 1975). Briefly, the soil sample was shaken with 0.1 M Na4 P2 O7 for 5–6 h. After centrifugation at 4000 rpm for 5 min, the supernatant was separated from the solid pellet. The operation was repeated 3×, and the supernatant solutions were combined. The solid residue was further extracted with 0.1 M NaOH by shaking the mixture for 3–4 h and centrifuging at 4000 rpm for 5 min to separate the supernatant solution from a pellet with the solid phase. The procedure was repeated until the final extract showed a pale colour that did not change in successive extractions (5–10× depending on the concentration and extractability of organic C).
After acidification of the extract with 6 M HCl to pH = 1.5 the HAs were isolated as a precipitate from the supernatant which formed the fulvic fraction, after allowing to settle overnight. The HA fraction was re-dissolved in 0.5 M NaOH, and centrifuged at 20,000 rpm for 5 min to remove clays and particulate OM. The HAs were precipitated again with HCl, centrifuged and dialyzed for purification with distilled water under continuous agitation until free of chlorides (AgNO3 test), which took about 8 days. The 20 soil samples resulted in only 18 HAs extracts, since samples 2 and 4 yielded insignificant quantities of HAs.
2.3 Sample analysis
Total C, H and N content (%) were determined with a Perkin-Elmer 2400 Series 2 CNHS/O elemental analyser. The O content was obtained as the difference between 100% and the sum of measured C, N and H proportions. Based on previous analysis in Tablas de Daimiel wetland, as well as in the examination of the FTIR spectra of the HAs (lack of intense signal in the 600–400 cm−1 range), we assume that ash content is ca. 1% or less, so it should not affect significantly to our results.
Solid-state 13C NMR spectra were acquired with a Bruker DSX 200 (4.7 Tesla) operating at 50.29 MHz for 13C, with HA samples contained in zirconium dioxide rotors of 7 mm diameter in a commercial double bearing probe. The magic angle spinning speed was 6.8 kHz. The recycle delay was 300 ms. Cross polarization contact time was 1 ms, and a ramped 1H–pulse was applied to account for Hartmann-Hahn mismatches. Depending on sample, the number of free-induction decays was between 3000 and 20,000, and the line broadening ranged between 25 and 75 Hz. The chemical shifts were calibrated to tetramethylsilane (0 ppm) using glycine as external reference. For spectral interpretation, the following ranges and assignations were considered (Wilson 1987; Wilson and Hatcher 1988; Preston 1992; Preston et al. 1997; Huang and Feather 1988): 0–46 ppm, alkyl (13, methyl; 21, acetate; 30, polymethylene), 46–110 ppm, O/N-alkyl (56, methoxyl/α-amino; 73, glucopyranosyde-derived; 103, anomeric C in carbohydrate; 105, quaternary aromatic C in tannins); 110–160 ppm, aromatic/unsaturated [ca. 135: unsubstituted, ca. 145, heterosubstituted: guaiacyl (G) lignins/dihydroxyls of tannins; ca. 153: ether-linked (syringyl (S) lignins)/tannins]; 160–200 ppm, carbonyl (172, carboxyl/amide; 198, ketone/aldehyde).
Areas in nine spectral regions were measured with ImageJ software (http://rsb.info.nih.gov/ij/) and normalized to 100.
2.4 Data treatment
The molecular mixing model (MMM) of Nelson and Baldock (2005) was applied to the NMR data as described by Rodríguez-Murillo et al. (2011). The used 13C spectral intensity distribution among the seven chemical shift regions for the six components was obtained from Baldock et al. (2004), who also supplied the appropriate elemental atomic H/C, N/C and O/C ratios for the six model components. Based on these data and the 13C spectral intensity distribution determined from the 13C NMR spectra of our samples, HAs composition was calculated in terms of the six generic biomolecules plus ‘pure carbonyl’ by solving a system of equations for each sample using the generalized reduced gradient nonlinear optimization code in the Solver add-in of Microsoft Excel with two restrictions: (i) the sum of component fractions was forced to be 100, and (ii) the component fractions were forced to be equal to or greater than zero.
3 Results
3.1 Yield of HAs and elemental composition
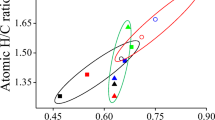
The HA content and composition varied considerably among the different sampling points (Table 1). From the Van Krevelen diagram (Van Krevelen 1950) shown in Fig. 2, we could distinguish up to three different classes of samples: predominantly aliphatic (high H/C atomic ratio), predominantly aromatic, mature (low H/C) and highly oxidized (high O/C atomic ratio).

Van Krevelen’s diagram of humic acids (HAs) from the sampling points of Tablas de Daimiel
The 13C NMR CPMAS spectra of the HAs are depicted in Fig. 3a. The respective spectral intensity distribution, as well as total aromaticity and aliphaticity, are given in Table 2. Two pairs of spectra (WS and HA), from two differentiated sampling points, are given in Fig. 3b.

a NMR spectra of the HAs. F: at least a fire has been recorded in the area of the sampling point. NF no fire recorded. b Comparison between 13C NMR spectra of whole sample and its humic acids: the spectral ranges for the main C-types are indicated
Clear geographic differences were evident between HAs, with the highly aliphatic HA corresponding to the upper Tablas sector, with non-permanent flooding and charophyte vegetation in open water areas whereas comparatively more aromatic HAs dominate in lower Tablas, in which the permanent vegetation is dominated by vascular plants. Our data show an increase in C/N of the HAs compared to the WS (Rodríguez-Murillo et al. 2011) in 16 of the 18 samples. Only for samples 16 and 19 was the C/N of the WS greater than that of the HA.
3.2 Composition of the HAs inferred from the MMM
The HAs composition obtained from applying the MMM is shown in Fig. 4. We did not introduce the experimental C/N values as a condition in our system of equations, as could be done (Nelson and Baldock 2005), because in the MMM scheme, only peptides are significant N-bearing compounds. This has indeed been confirmed in 15N NMR spectroscopic studies (Knicker 2000), despite the fact that in fire-affected soils, as many of our samples are a sizeable fraction of organic N could be present in heterocyclic compounds (Almendros et al. 2003; Knicker 2011), and we would overestimate the proteinaceous contribution assigning all N to this fraction.

HA composition according to molecular mixing model (MMM). HAs are ordered from lower aromaticity (bottom) to higher aromaticity (top). I irregular; S seasonal; P permanent. Flooding regime refers to that before 1973, year of the onset of large perturbation in the hydrological regime of the Tablas. Fires are those recorded before sampling year (2007); at sites 15, 17, and 17′, the existence of fires in the past has been inferred from the occurrence of charred material in the samples. Y means that at least one fire has been recorded and N means no fire recorded in the site
As is the case in WS samples (Rodríguez-Murillo et al. 2011), the six organic components of the HAs were widespread. Structural components as carbohydrate, protein and lipid dominate the HA composition across the wetland, and variable quantities of lignin and char were also present in all the HAs.
The reliability of the MMM applied to HAs was assessed in two independent ways:
-
(i)
Calculating error in total spectrum area as the sum of absolute values of differences between experimental (spectrum) areas and calculated (MMM) areas for each chemical shift region (Nelson and Baldock 2005).
-
(ii)
Calculating H/C and N/C ratios from the composition obtained with the MMM and comparing the values with the corresponding H/C and N/C values from elemental analysis.
Mean error in total area across HA samples was 2.22 ± 0.75, greater than that (0.93) found in WS (Rodríguez-Murillo et al. 2011; Fig. 5a). The spectral regions reproduced most poorly were 95–110 ppm (C1 region of carbohydrate-like structures) and 145–165 ppm (N/O aromatic region). In the former, the mean error for calculated versus measured area was 32% and in the second 4.5%. In the remaining chemical shift regions, differences between calculated and measured spectral intensities were lower. It was estimated that a mean error “x” in determining intensity gives a double error (“2×”) in the composition of the HA.

a Performance of MMM. Area error is the sum of absolute values of differences (%) between experimental and calculated areas for each chemical shift region in each sample. b Performance of MMM. Calculated versus experimental H/C ratios of the samples. The lower line represents the perfect concordance calculated–experimental. c Performance of MMM. Calculated versus experimental N/C ratios of the samples
A strong correlation between MMM-calculated and experimental H/C ratio was obtained, but with some bias: calculated H/C values were bigger than experimental H/C ones, i.e. real samples were more aromatic than MMM estimated composition (Fig. 5b). The opposite happened with the N/C ratio, the calculated values being generally lower than the experimental, but with a less systematic trend in comparison with the H/C ratio case (Fig. 5c). Possible causes of the discrepancy between calculated and experimental ratios are discussed below in Section 4.1.
3.3 Relationship among relative proportions of HA components and comparison between HAs composition and the OM composition of wetland WS
The most significant correlations among HA components (Table 3) were between char and carbohydrate (R −0.798), carbonyl and protein (R −0.781), lipid and lignin (R −0.709), lignin and protein (R 0.686) and carbohydrate and protein (R 0.680). Every significant relationship was negative, except in the case of carbohydrate-protein, carbohydrate-lipid and carbonyl-char. In the wetland WS, the strongest correlation was lignin-protein, but the char-carbohydrate correlation was also noticeable (Rodríguez-Murillo et al. 2011).
The C/N ratio was positively related to lignin (R 0.789) and carbonyl (R 0.522), and negatively related to the other constituents (mainly to protein, R −0.698). No significant relationship was found with char.
In Table 4, the HA composition is compared with the composition of the OM of the corresponding WS sample. The HAs differ from WSs mainly because of the higher aromaticity (relative area of 110–165 ppm region of 13C NMR spectra; Fig. 6a). This increase in aromaticity in HA compared to WS was fairly linear (HA aromaticity =0.912× (WS aromaticity) + 10.39; R 0.825, P < 0.0001) indicating that WT and HA aromaticities are significantly related. In the data set of Mahieu et al. (1999), the mean increase in aromatic C content of HAs versus WSs was 10%, which is in line with our results. The total aromaticity depends strongly upon the sum of lignin content and char content: 94% is explained by lignin + char, which were by far the most significant contributors.

Comparison of components of HAs with components of organic matter in whole samples (WS) according to MMM. a Comparison of aromaticities. b Comparison of char contents. c Comparison of carbohydrate contents
The sum of lignin and char in HAs was strongly related to that of WSs (HA = 0.879× WS + 15.39,. R 0.755. Significant correlation also existed (Fig. 6b) between WS and HA for char alone (R 0.663, P = 0.0027) which increased from WS to HA, and for carbohydrates, which decreased sharply in the HA compared to the WS (Fig. 6c). In the latter, the carbohydrate content of HA was ca. 50% of that of WS (on average), with R 0.747. For the HAs, the content of this compound class was significantly related to that of WS from the different sites. Lipid constituents in WS and HA were barely related R 0.527), and protein, lignin and carbonyl showed non-significant relationships (at P < 0.05) between their contribution to HA and to WS (protein, R 0.401; lignin, R 0.374; carbonyl, R 0.346).
3.4 Influence of environmental factors in HA-WS component relationships
Humification is to a large extent influenced by environmental factors such as aerobic or semi-anaerobic conditions for plant detritus diagenesis. Wetlands are subjected to very diverse local physicochemical processes of variable intensity. In the case of oxidizing environment, the OM tends towards a progressive depletion in carbohydrate, whereas in the case of anaerobic conditions, the reducing environment leads to a selective increase of alkyl and lignin constituents (Filip and Alberts 1994). On the other hand, vegetation fires could produce severe thermal modification of existing OM, as well as the formation of new carbon forms. During a fire, considerable rearrangement of C forms occurs and slightly more refractory and oxidation-resistant organic C forms with increased mean residence time, including black carbon, are formed (Schulze et al. 2000; Almendros et al. 2003).
We analysed the effect of environmental conditions on humification (i.e. as reflected in the composition of the HA fraction), by studying the relationship between HA and WS separately in the different groups of sampling sites, which could be distinguished based on two factors, viz., flooding regime and fire occurrence, by taking advantage of information recorded from Tablas de Daimiel (Cirujano and Álvarez-Cobelas, personal communication) and our own observations (Table 1).
Dividing our sample set into those groups (permanent/non-permanent flooded; fire-affected/not fire-affected) and recalculating the correlations between the aromaticity, char and carbohydrate contents for WS and HA (as the most significantly related variables in the whole set of samples) clearly demonstrated the impact of environmental conditions. Further differentiation between sampling sites with permanent and non-permanent flooding (irregular or seasonal in Table 1), showed strong correlation between the aromaticity of HS and of WS for the non-permanent flooding group (R 0.964, P = 0.0001), but only a weak relationship for the permanent flooding group (R 0.591, P = 0.072). If fire-affected and unaffected areas are differentiated, the aromaticity of HA and of WS are not related in fire areas (R 0.533, P = 0.14), but shows a tight relationship for no-fire areas (R 0.941, P = 0.0002). HA aromaticity is higher and more constrained (30-ca. 45%) in fire areas in comparison with no-fire areas (15-ca. 40%).
Char content of HA and WS are significantly correlated in non-permanently flooded samples (R 0.964, P = 0.0008) and fire-unaffected areas (R 0.940, P = 0.0002) (Fig. 7a), but no significant relationship between char content of HA and of WS char was observed for permanently flooded sites, as well as fire-affected areas (R 0.055) (Fig. 7b). The relationship between carbohydrates in HA and WS was relatively strong and positive for non-permanently flooded areas (R 0.821, P = 0.013) (not shown) and fire-unaffected areas (R 0.733, P = 0.025) (Fig. 8a), but not significant in the case of permanently flooded and fire-affected areas (R 0.644, P = 0.061) (not shown). Insignificant correlation was apparent for the other components (lipid, protein and lignin) of WS and of HA.

Comparison of components of HAs with components of organic matter in whole samples (WS), according to MMM. Effects of fire on char content change. a Comparison of char content in no-fire areas. b Comparison of char content in fire areas

Comparison of components of HAs with components of organic matter in whole samples (WS) according to MMM. Effects of fire on carbohydrate content change
4 Discussion and conclusions
Previously (Rodríguez-Murillo et al. 2011), we studied the composition of OM in wetland soils of Tablas de Daimiel. Based on a combination of solid-state CPMAS 13C NMR spectroscopy with MMM (Nelson and Baldock 2005; Baldock et al. 2004), significant differences were found in the composition of wetland soil OM derived from different areas. The differences were ascribed mainly to the nature of the original detrital OM (non-vascular, vascular, mixed and microbial). Additional perturbations such as fires, allochthonous input and movement of sediment (resuspension) could be invoked to justify the presence of lignin and perhaps of char in areas with non-vascular vegetation. Thus, Tablas de Daimiel wetland can be considered as an interesting, mosaic-like ecosystem, with large variation in soil OM characteristics caused by varying vegetation, environmental conditions and disturbances in the different areas.
Humification processes are also likely to differ in local environments depending on the spatial variability of the microtopography and the duration of flooding. Carbohydrates are the most labile macromolecular components and tend to be depleted during the microbial and abiotic reworking of vegetal detritus. Proteins are more resistant, but are also depleted in part, with an effect of increasing the C/N ratio in the HAs versus that of the whole sample. Progressive accumulation of aromatic structures in the course of HA formation is apparent in the sharp increase in lignin-type structures, as well as carbonyl and aromatic char. A Van Krevelen diagram offers some information on HA characteristics. Decreasing H/C ratio is frequently interpreted as progressive transformation of the OM towards a greater proportion of aromatic carbon, whereas higher H/C implies a molecular composition more similar to that of plant biomass. Besides, aliphatic samples have low C/N, and aromatic high C/N, and this can be related to the C/N of original OM (predominantly non-vascular in aliphatic samples and more vascular in mature samples) and/or different degree of transformation.
Figure 2 illustrates the fact that in the Las Tablas ecosystem there are HAs of very different nature: predominantly aliphatic HAs would correspond with WSs containing OM of aliphatic and alkyl character (non-vascular vegetation and microbial origin) (Rodríguez-Murillo et al. 2011), indicating that the initial signature of detrital OM is kept through HA extraction. In particular, HA extraction preserves (although more weakly) the negative relationships between lignin and protein found in WSs, perhaps reflecting the original balance between vascular/non-vascular vegetation. The main components of HAs, as calculated with MMM, were lipid>char>protein, whereas in whole sample, the main components were protein>lipid>carbohydrate (Table 5). The relative content of carbohydrate is progressively depleted in the course of humification, whereas, as expected, char and—to a lesser extent—lignin, increase in concentration. Increase in lignin—comparing HA and WS—was not uniform in the samples, and lignin, often considered the basis for HA structure in the case of peatland soils, was not the main component identified in our HAs applying the MMM.
4.1 Error assessment in the determination of composition from MMM
An obvious source of error in the determination of HA composition using the MMM could be the occurrence of compounds other than the six generic constituents considered, for instance, tannins. These polyphenols, present in higher plants (Kögel-Knabner 2002), display an intense NMR signal as two peaks in the 95–110 ppm region—which was the worst reproduced signal with MMM—, due to their aromatic quaternary carbon (Preston et al. 1997). The 95–110 ppm region would then show considerable overlapping between anomeric O-alkyl carbons in carbohydrates and aromatic quaternary carbon atoms in tannins and dilignols. Although tannins are usually far less abundant than the other MMM constituents, only 16.7% of carbohydrates, 8.6% of lignin and 6.3% of char peak in the 95–110 ppm region, which is compatible with a small proportion of tannins making a significant contribution to the signal in the 95–110 ppm range. Incomplete agreement between experimental and simulated spectra (i.e. errors in total spectrum area) might also be attributed to experimental errors in 13C NMR spectra of HAs, but, considering the better agreement obtained in many cases using natural organic materials (Nelson and Baldock 2005; Rodríguez-Murillo et al. 2011) instead of HAs, the occurrence in HAs of specific chemical structures with special 13C NMR signatures not considered in the MMM treatment could be an important factor explaining the observed discrepancies. Unfortunately, our data and their analysis can neither prove nor disprove the presence of such structures, which should be quantitatively minor components.
Systematic discrepancies in experimental versus calculated H/C (Fig. 5b) and N/C (Fig. 5c) ratios were also probably due to the above considerations. There was no apparent relationship of differences in experimental and calculated H/C (Fig. 5b) with sample aromaticity or sample char content, although as H/C (experimental) increased (less ‘low proton’ components), differences in H/C were smaller. The existence of a significant proportion of N in heterocyclic structures could explain discrepancies in N/C ratio.
4.2 Humification mechanisms
In the areas permanently flooded in the past, fire has been a recent perturbation of Las Tablas, coinciding with the first desiccation events (around 1960). Such fires, especially in peat zones, have affected humus formation, as could be deduced from the different relationships of char and carbohydrate content for fire and no-fire areas. At first sight, differences in humification could be empirically ascribed to either fire action or flooding regime; the occurrence of fire in our system seems to parallel the effects of the permanent flooding in HA–WS component relationships, but as discussed below, fire effects could explain these relationships, which is not the case for flooding. This coincidence between HA and WS relationships in areas subjected to such different environmental influences (flooding and fire) probably arises from the fact that fires occur mainly in former permanently flooded areas, after desiccation. Fire consumes OM, but, at the same time, increases aromaticity, and the content of refractory substances, such as char, which is a potential contributor to the formation of refractory long-term carbon sinks (Forbes et al. 2006). This is due to selective enrichment of residual heat-resistant, strongly aromatic components and also to endothermic neoformation reactions involving carbohydrates, but also amino acids and unsaturated alkyl chains (González-Pérez et al. 2004).
The concentration of char in HA was unrelated to the accumulation of WS char in fire areas, but depended linearly on WS char in no-fire areas (Fig. 7a, b).
We can hypothesise that, in no-fire areas, HA ‘char’ comes mainly from char formerly introduced into the sediment, as suggested by Haumaier and Zech (1995), and is selectively preserved, whereas in fire-affected areas, char is newly formed in situ as a consequence of fire action. New formation of char in HAs in fire-affected areas could explain the loss of relationship between WS char and HA char in these areas. Then, analysis of HA composition using MMM allows some insight into humification mechanisms, comparing HA and whole sample composition: MMM char determination and constituent comparison between WS and HA allows the detection of two contrasting HA-formation mechanisms, depending on the occurrence (or not) of fire.
References
Aiken GR, McKnight DM, Wershaw RL, MacCarthy P (1985) Humic substances in soil sediment and water. Wiley, New York
Almendros G (2008) In: Chesworth W (ed) Encyclopedia of Soil Science. Springer, Dordrecht, pp 315–323
Almendros G, Knicker H, González-Vila FJ (2003) Rearrangement of carbon and nitrogen forms in peat after progressive isothermal heating as determined by solid-state 13C- and 15N-NMR spectroscopies. Org Geochem 34:1559–1568
Baldock JA, Masiello CA, Gélinas Y, Hedges JI (2004) Cycling and composition of organic matter in terrestrial and marine ecosystems. Mar Chem 92:39–64
Bridgham SD, Megonigal JP, Keller JK, Bliss NB, Trettin C (2006) The carbon balance of north American wetlands. Wetlands 26:889–916
Burdon J (2001) Are the traditional concepts of the structures of humic substances realistic? Soil Sci 166:752–769
Buurman P, Nierop KGJ, Kaal J, Senesi N (2009) Analytical pyrolysis and thermally assisted hydrolysis and methylation of EUROSOIL humic acid samples—a key to their source. Geoderma 150:10–22
Duchaufour P, Jacquin F (1975) Comparaison des processus d’humification dans les principaux types d’humus forestiers. Bull Assoc Fr Etud Sol 1:29–36
Filip Z, Alberts JJ (1994) Microbial utilization resulting in early diagenesis of salt-marsh humic acids. Sci Total Environ 144:121–135
Forbes MS, Raison RJ, Skjemstad JO (2006) Formation transformation and transport of black carbon (charcoal) in terrestrial and aquatic ecosystems. Sci Total Environ 370:190–206
González-Pérez JA, González-Vila FJ, Almendros G, Knicker H (2004) The effect of fire on soil organic matter—a review. Environ Int 30:855–870
Haumaier L, Zech W (1995) Black carbon--possible source of highly aromatic components of soil humic acids. Org Geochem 23:191–196
Hayes MHB, MacCarthy P, Malcolm RL, Swift RS (1989) The search for structure: setting the scene. In: Hayes MHB, MacCarthy P, Malcolm RL, Swift RS (eds) Humic Substances. Wiley, Chichester, pp 3–32
Huang RD, Feather MS (1988) 13C NMR study of some Maillard reaction products arising from D-glucose-DL-alanine interactions. J Agr Food Chem 36:673–676
Kelleher BP, Simpson AJ (2006) Humic substances in soils: are they really chemically distinct? Environ Sci Technol 40:4605–4611
Kleber M (2010) What is recalcitrant soil OM? Environ Chem 7:320–332
Kleber M, Johnson MG (2010) Advances in understanding the molecular structure of soil organic matter: implications for interactions in the environment. Adv Agron 106:77–142
Knicker H (2000) Biogenic nitrogen in soils as revealed by solid state carbon-13 and nitrogen-15 nuclear magnetic resonance spectroscopy. J Environ Qual 29:715–723
Knicker H (2011) Soil organic N - an under-rated player for C sequestration in soils? Soil Biol Biochem 43:1118–1129
Kögel-Knabner I (2002) The macromolecular organic composition of plant and microbial residues as inputs to soil OM. Soil Biol Biochem 34:139–162
Lehmann J, Solomon D, Kinyangi J, Dathe L, Wirick S, Jacobsen C (2008) Spatial complexity of soil organic matter forms at nanometre scales. Nat Geosci 1:238–242
Mahieu N, Powlson DS, Randall EW (1999) Statistical analysis of published 13C CPMAS NMR spectra of soil OM. Soil Sci Soc Am J 63:307–319
Nelson PN, Baldock JA (2005) Estimating the molecular composition of a diverse range of natural organic materials from solid state 13C NMR and elemental analyses. Biogeochemistry 72:1–34
Piccolo A (2001) The supramolecular structure of humic substances. Soil Sci 166:810–832
Preston CM (1992) The application of NMR to organic matter inputs and processes in forest ecosystems of the Pacific northwest. Sci Tot Environ 113:107–120
Preston CM, Trofymow JA, Sayer BG, Niu J (1997) 13C nuclear magnetic resonance spectroscopy with cross-polarization and magic-angle spinning investigation of the proximate analysis fractions used to assess litter quality in decomposition studies. Can J Botany 75:16011–16613
Rodríguez-Murillo JC, Almendros G, Knicker H (2011) Wetland soil organic matter composition in a Mediterranean semiarid wetland (Las Tablas de Daimiel Central Spain): an insight into different carbon sequestration pathways. Org Geochem 42:762–773
Sánchez-Carrillo S, Angeler DG (2010) Ecology of threatened semi-arid wetlands. Springer, Dordrecht
Schulze ED, Wirth C, Heimann M (2000) Managing forests after Kyoto. Science 289:2058–2059
Sutton R, Sposito G (2005) Molecular structure in soil humic substances: the new view. Environ Sci Technol 39:9009–9015
Van Krevelen DW (1950) Graphical-statistical method for the study of structure and reaction processes of coal. Fuel 29:269–284
Wilson MA (1987) NMR techniques and applications in geochemistry and soil chemistry. Pergamon, Oxford
Wilson MA, Hatcher PG (1988) Detection of tannins in modern and fossil barks and in plant residues by high-resolution solid-state 13C nuclear magnetic resonance. Org Geochem 12:539–546
Acknowledgements
This study was supported by project 81/2005 of the Spanish CICYT and Organismo Autónomo Parques Nacionales, and by project CGL2008-04296. We wish to thank M. Álvarez-Cobelas and S. Cirujano for environmental information about Tablas, and the staff of the National Park “Las Tablas de Daimiel” for their help, as well as the Analytical Unit staff of MNCN and our laboratory assistants. The NMR measurements were performed at the Lehrstuhl für Bodenkunde of the TU-München-Weihenstephan, Germany, in the frame of the Acciones Integradas Program, within which the exchange of involved scientists was financed by the DAAD (Germany)-CICyT(Spain).
Author information
Authors and Affiliations
Corresponding author
Additional information
Responsible editor: Jianming Xu
Rights and permissions
About this article

Cite this article
Rodríguez-Murillo, J.C., Almendros, G. & Knicker, H. Humic acid composition and humification processes in wetland soils of a Mediterranean semiarid wetland. J Soils Sediments 17, 2104–2115 (2017). https://doi.org/10.1007/s11368-017-1663-y
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s11368-017-1663-y