
Abstract
Purpose
Lead (Pb) pollution is appearing as an alarming threat nowadays in both developed and developing countries. Excessive Pb concentrations in agricultural soils result in minimizing the microbiological activities which leads to the decrease in crop production. A pot experiment was conducted with the purpose to examine the deleterious effect of Pb on microbiological index under spinach cultivation.
Materials and methods
Pb was added to 5 kg soil in each pot (with 6 seeds/pot) using Pb(NO3)2 at the rate of 0, 150, 300, 450, and 600 mg kg−1 with three replications in completely randomized design. All soil microbial, enzymatic, and chemical properties and plant growth parameters and nutrient uptake were measured by standard methods.
Results and discussion
Both soil and plant measured parameters decreased after the addition of Pb (150, 300, 450, and 600 mg Pb kg−1 soil) treatments with the passage of time (from 15 to 60 days) compared with control (CK). However, high Pb levels had more suppressive effect, therefore, highest Pb level (600 mg Pb kg−1 soil) significantly (P < 0.05) decreased the microbial biomass carbon (5.59-fold); microbial biomass nitrogen (N; 11.71-fold); microbial biomass phosphorus (P; 25.1-fold); dehydrogenase (4.02-fold); phosphatase (9.40-fold); urease (9.26-fold); pH (1.40-fold); spinach shoot (2.17-fold) and root (2.54-fold) length; shoot (2.36-fold) and root (2.69-fold) fresh weight; shoot (3.90-fold) and root (3.50-fold) dry weight; chlorophyll content (5.60-fold); carotenoid content (4.29-fold); plant macronutrients uptake, i.e., N (4.38- and 2.97-fold), P (3.88- and 6.58-fold), K (3.88- and 4.6-fold), Ca (6.60- and 6.70-fold), and Mg (5.57- and 4.45-fold); and plant micronutrient uptake, i.e., Zn (2.39- and 3.05-fold), Cu (3.70- and 2.62-fold), Fe (4.13- and 3.23-fold), and Mn (4.17- and 4.09-fold) in spinach shoot and root, respectively. Conversely, highest Pb level, i.e., 600 mg Pb kg−1 soil significantly (P < 0.05) increased the biomass carbon (C)/nitrogen (N) (4.69-fold) and C/P (6.01-fold) ratios, soil extractable Pb (5.87-fold), and Pb uptake in spinach shoot (3.58-fold) and root (4.38-fold), respectively, at the end of the experiment, i.e., day 60.
Conclusions
Pb contamination significantly decreased the soil microbial and enzymatic activities, pH, spinach plant growth, and nutrients uptake in all the samples spiked with Pb. The degree of the influence increased with the increased Pb concentrations and incubation time, showing that Pb threshold is strongly associated with the extent of Pb concentration and time to accumulate. The soil microbial biomass, enzymatic activities, pH, and spinach physiological indices, could be used as a sensitive indicators to reflect environmental stress in soil ecosystems.
Similar content being viewed by others


Explore related subjects
Discover the latest articles, news and stories from top researchers in related subjects.Avoid common mistakes on your manuscript.
1 Introduction
Contamination of agricultural soils with heavy metals is a widespread global problem (Tandy et al. 2006) and also a major environmental concern over the past several decades. Heavy metal contamination in soil is caused by various sources, such as industrial processes, manufacturing, disposal of industrial and domestic refuse, and agricultural practices (Hassan et al. 2013a). Heavy metals are not subjected to degradation processes and therefore they remain almost indefinitely in the environment, and accumulate in different parts of the food chain (Hassan et al. 2013a). The food chain contamination is one of the most important pathways for the entry of these toxic pollutants into the human body (Khan et al. 2008). Heavy metals at high concentrations generally affect the growth morphology and metabolism of soil biota (Hassan et al. 2013a). Khan et al. (2010) reported that heavy metals could have long-term hazardous impacts on the health of soil ecosystems and adverse influences on soil biological processes. Heavy metals, such as lead (Pb), cadmium (Cd), chromium (Cr), zinc (Zn), and nickel (Ni) are important environmental pollutants, particularly in areas with high anthropogenic pressure (Majid et al. 2012). Among these heavy metals, Pb is the most abundant and ubiquitous and plays a major role in the contamination of soil and atmosphere (Bindler 2011).
Pb has been mined and used by humans for several thousand years (Reimann et al. 2012). The accumulated total world Pb production is estimated to be 300 megatonnes, in it the portion of the anthropogenic Pb fraction ranged from <10 to >90% (Reimann et al. 2012). It has been reported that an increase of 1 × 106 t year−1 occurred because of emerging industrial activities together with the introduction of leaded gasoline (Komarek et al. 2008). The Pb enters the environment via several anthropogenic activities, e.g., during production (including mining and smelting), use (batteries, pigments, ceramics, and plastics), recycling, disposal of Pb compounds, combustion of fossil fuels (coal and former use of leaded gasoline), use of mineral fertilizers and sewage sludge application (Komarek et al. 2008), and further discharge and disperse into nearby agricultural soils, food crops, and stream systems. Considering its low solubility and relative resistance to microbial degradation, Pb and its compounds tend to accumulate in soils and sediments, where, they remain accessible to food chain and to human metabolism (Reimann et al. 2012). The Pb-affected soils contain Pb in the range of 400–800 mg kg−1 where the industrialized areas contain Pb up to 1000 mg kg−1 (Sharma and Dubey 2005). In almost all towns of Pakistan, sewage is directly used for irrigating about 32,500 ha agricultural land (Ensink et al. 2004). The Pb concentration in the agricultural soils of the Pakistan ranges from 20 to 500 mg kg−1 (Nasreen 2006).
As soil is a sink for all types of materials, from which metals ultimately reach to ground water, plants, and animals, commonly referred to as soil–water–plant–animal systems (Murtaza et al. 2010). Microbial community is an integral part of the soil ecosystem because it regulates significant processes, such as nutrient cycling and decomposition of organic matter (Hassan et al. 2013b; Omirou et al. 2011). Microbial biomass is a term used to indicate the total amount of organisms in the soil (Nannipieri et al. 2003). Microbial biomass represent only 5% of the organic matter but plays a critical role in soil fertility, hence, known as a good general measure of soil health (Gonzales-Quiñones et al. 2011), as it has been estimated that 80–90% of the processes in soil are reactions mediated by microbial decomposers (Nannipieri and Badalucco 2003). Generally, microbes and microbe-mediated processes are the most sensitive to perturbations in the soil and can be used as early indicators of changes in the soil environment (Nannipieri et al. 2003). Soil enzymes are pervasive in soil environments; they are vital activators in life processes (Hassan et al. 2013c). Similarly in the soil, they are known to play substantial roles in the biochemical degradation of the organic matter, catalyzing several reactions necessary for the life processes of microorganisms in soils, the decomposition of organic wastes, and nutrient cycling, providing an early indication of the history of a soil and its changes in agricultural management and other external stresses, e.g., heavy metals (Kandeler et al. 2006; Eldor 2007). The Pb in higher concentration has a negative effect on ecological consequences and microbiological parameters which are indicators of soil quality (Filip 2002) and inhibits enzyme activities by reacting with their sulphydril groups (Sharma and Dubey 2005). The short-term exposure of metals to soil results in greater reduction of microbial and enzymatic activity (Rantalainen et al. 2006; Mishra et al. 2006).
Pb toxicity affects the morphology, growth, and photosynthetic processes of plants and causes inhibition of enzyme activities, water imbalance, alterations in membrane permeability, and disturbs mineral nutrition (Sharma and Dubey 2005). The Pb toxicity in plants adversely affects the process of photosynthesis, and its higher concentration in soil imbalance the mineral nutrients in growing plants (Robinson et al. 2008). Significant changes in nutrient contents as well as in internal ratio of nutrients occur in plants under Pb toxicity (Robinson et al. 2008). Pb physically blocks the entry of cations (K+, Ca+2, Mg+2, Mn+2, Zn+2, Cu+2, and Fe+3) and anions (NO3 −) in the root system (Sharma and Dubey 2005). Vegetables are typically common crops in Pakistan that irrigate with sewage and fetch high prices in the nearby urban markets (Murtaza et al. 2010). Leafy vegetables are an important part in the human diet. Spinach (Spinacia oleracea L.) is a leafy vegetable which belongs to the family Chenopodiacae. Spinach has a high nutritional value and is an important source of minerals and is extremely rich in antioxidants (Salk et al. 2008). Spinach is the most important and highly nutritious green leafy winter vegetable grown both worldwide and in Pakistan on a large scale (Waseem and Nadeem 2001). The area under spinach cultivation and spinach production in Pakistan are 7,706 ha and 86,598 tonnes and in Punjab the area under spinach cultivation and production of spinach are 3,110 ha and 86,598 tones (Agricultural Statistics of Pakistan 2010–2011). To our knowledge, effect of Pb pollution on microbial biomass and enzymes activity under spinach cultivation has not been studied before and is still poorly understood. Therefore, the present study was conducted with the objectives (1) to compare the effects of different levels of Pb (0, 150, 300, 450, and 600 mg kg−1) on the dynamics of soil microbial biomass and enzymes activity; (2) to examine the effects of different Pb levels on growth, macro- and micronutrients, and Pb uptake of spinach plant.
2 Materials and methods
2.1 Soil sampling
Samples (0–15 cm depth) of Alfisol soils collected from, Dera Ghazi Khan City of Punjab (30°03″ N and 70°38″ E) were used in this study. Filed-moist soil samples were sieved (<2 mm) and divided into two subsamples. One subsample was used to conduct the pot experiment. The other subsample was air-dried at room temperature and used for chemical analyses.
2.2 Experimental layout
The study was conducted in a randomized complete block design with ten treatments in triplicates. The sieved soil (5 kg, 2 mm) was transferred into 15 earthen pots (28 × 34 cm). Pb was applied as Pb(NO3)2 solution to soil in each pot maintaining the concentrations of 0 (Pb0), 150 (Pb1), 300 (Pb2), 450 (Pb3), and (Pb4) 600 mg kg−1 of soil. Soils mixed with Pb were allowed to equilibrate for 15 days and then seeds of spinach were sown at the rate of six seeds per pot which were thinned up to three seedlings in each pot after germination. Soil samples were collected from each pot at 15, 30, 45, and 60 days after sowing and analyzed for microbial biomass (microbial biomass carbon (MBC), microbial biomass nitrogen (MBN), and microbial biomass phosphorus (MBP)), enzymes activities (dehydrogenase, phosphatase, and urease), extractable Pb and pH. Spinach plants were harvested after 60 days of sowing, and parameters regarding root length, root fresh and dry weight, shoot length, shoot fresh and dry weight, macronutrients in shoot and root, micronutrients in shoot and root, and Pb accumulation in spinach was measured. The treatments of the experiment are listed in Table 1.
2.3 Physicochemical analysis of the soil
Soil particle size distribution was evaluated by the international pipette method (Hassan 2013). Soil pH and electrical conductivity were measured using soil/water ratio (w/v) of 1:2 (Hassan et al. 2013a). The cation exchange capacity was determined by the method of Anderson and Ingram (1993). Total organic carbon (TOC) of the soil was determined by potassium dichromate (K2Cr2O7) oxidation at 170–180 °C followed by titration with 0.5 mol l−1 ferrous sulfate (Walkley and Black 1934). Soil available N was measured by Kjeldahl method after using KCl (2 mol l−1) as an extractor, while the available K content was estimated by flame photometer after its extraction by NH4COO–CH3 (1 mol l−1, pH 7) as described by Jackson (1973). Soil available phosphorus (P) was assayed by NaHCO3 method (Hesse 1972). Total P in soil samples were analyzed by NaOH fusion and colorimetric procedures (Olsen and Somers 1982), while total N was determined by the method of Buresh et al. (1982) by adding 30 ml of concentrated H2SO4 and 10 g of digestion mixture (K2SO4/FeSO4/CuSO4, 10.0:1.0:0.5) in 10 g of soil. Extractable Pb (milligrams per kilogram) in the soil was calculated by using the atomic absorption spectrophotometer, after treatment with AB-DTPA solution (Page et al. 1982). Some pertinent characteristics of the soil are shown in Table 2.
2.4 Analysis of soil microbial biomass
The chloroform fumigation-extraction method was used to measure soil MBC. Soil sample equivalent to 10 g (fresh soil) was fumigated for 24 h at 25 °C with alcohol-free chloroform (CHCl3) in a vacuum desiccator containing soda lime. The fumigated soil was then transferred into a clean empty desiccator and residual CHCl3 was removed from the fumigated soils by repeated evacuations. The fumigated soil was extracted immediately for 30 min by using horizontal shaking at 200 rpm with 50 ml 0.5 M K2SO4 and filtered through a filter paper (Whatman No. 40). The non-fumigated control soil (10 g fresh soil) was extracted simultaneously when fumigation commenced. TOC in the extracts was measured as CO2 by infrared adsorption after combustion at 760 °C using a Shimadzu automatic TOC analyzer (Shimadzu Corp. Japan). MBC was calculated as (Ct1 − Ct0) × 2.22, where Ct1 is the extracted C in milligrams per kilogram from fumigated samples, Ct0 is the extracted C (milligrams per kilogram) from non-fumigated samples and 2.22 is the factor, calculated by 0.45, i.e., 100/45 = 2.22, here 0.45 is an extractable part of microbial C after fumigation (Wu et al. 1990). For MBN, total N in the K2SO4 extract was measured after Kajeldahl digestion. The soil MBN was calculated as (Nt1 − Nt0) × 1.85, where Nt1 is the extracted N (milligrams per kilogram) in fumigated samples, Nt0 is the N (milligrams per kilogram) in non-fumigated samples and 1.85 is a factor which was obtained via 0.54 (i.e., 100/54 = 1.85) which is an extractable part of microbial N after fumigation (Brookes et al. 1985). In the case of MBP, the fumigated and the non-fumigated soil samples were extracted by 0.5 M NaHCO3 (pH 8.5) for 30 min. The concentrations of P were determined using spectrophotometer at 882 nm wave length. The MBP was calculated as (Pt1 − Pt0) × 2.5, where Pt1 is the P (milligrams per kilogram) in fumigated samples, Pt0 is the P (milligrams per kilogram) from non-fumigated samples and 2.5 is a factor, computed by 0.4 (e.g., 100/40 = 2.5), while 0.4 is an extractable part of microbial P after fumigation (Brookes et al. 1985). Soil microbial biomass of the experimental soil is shown in Table 3.
2.5 Analyses of soil enzymes activity
Dehydrogenase activity was determined by the method of Öhlinger (1996). Fleetingly, 20 g air-dried soil was mixed with 0.2 g of CaCO3, and then 6 g of this mixture was placed in three different test tubes. Samples were incubated at 37 °C for 24 h after adding 1 ml of 3% aqueous solution of triphenyl tetrazolium chloride and 2.5 ml of distilled water. Then 10 ml of methanol was added and filtered after shaking. The red color intensity was measured with a spectrophotometer at a wave length of 546 nm. Soil dehydrogenase activity was expressed as mg triphenyl formazan (TPF) kg−1 dry soil 24 h−1. The method of Kandeler and Gerber (1988) was followed to analyze soil urease activity. In summary, 5 g soil was taken into 250 ml conical flask and 10 ml of urea solution was added along with 20 ml buffer solution (citric acid, KOH, and NaOH) having pH 6.7. The solution was filtered after incubation at 37 °C for 24 h and then 3 ml of filtrate was taken into 50 ml flask. Contents were mixed in the flask after adding 20 ml of distilled water and 4 ml of mixed reagent (Phenol + NaOH) in it. Then 4 ml of sodium hypochlorite solution was added, mixed and volume was made up to 50 ml with distilled water. The absorbance of blue color was checked at 578 nm through spectrophotometer. Soil urease activity was expressed as mg NH3–N kg−1 dry soil 24 h−1. Soil phosphatase activity was determined by following the procedure of Alef and Nannipieri (1995). Briefly, 1 g soil mixed with 0.2 ml of toluene, 4 ml of modified universal buffer of pH 11 plus 1 ml of p-nitro phenyl phosphatase solution and then the flask was placed in an incubator at 37 °C for 1 h. Then 1 ml of 0.5 M CaCl2 and 4 ml of 0.5 M NaOH were added and soil suspension was filtered (Whatman no. 2). Yellow color intensity was measured at 400 nm wavelength with a spectrophotometer. Soil phosphatase activity was expressed as milligrams phenol produced per kilograms dry soilper hour. Soil enzymatic activity of the experimental soil is shown in Table 3.
2.6 Plant analysis
2.6.1 Growth parameters
After harvest, roots and shoots were separated from soil and shoot and root lengths were measured using a meter rod. The fresh weight of shoots and roots were determined with the help of analytical balance, thereafter plants were oven dried at 70 °C to a constant dry weight and data for dry weights were recorded. Chlorophyll and carotenoid contents were determined by using the method of Lichtenthaler and Wellburn (1983), in the acetone extract (80%, v/v). Briefly, 0.2 g leaves were homogenized with 10 ml of 80% acetone, and the extract was centrifuged at 3,000×g for 5 min. The upper phase was transferred into a new tube and absorbance was measured at 663, 646, and 470 nm, respectively, for chlorophyll a and b and carotenoid contents.
2.6.2 Macro- and micronutrients in plant
For the determination of plant nitrogen (N), 0.5 g of dry plant material was taken in 100 ml digestion tube. Then pumic boiling granule and 3 g catalyst mixture was added, immediately followed by 10 ml of concentrated H2SO4 addition. After stirring on vortex mixer tube was placed at block digester set at 100 °C for 20 min, then volume was brought to 100 ml. Then 10 ml of 10 N NaOH solution was added and distillated at distillation apparatus for 10 min. The 35-ml distillate was collected and titrated against 0.01 N H2SO4 (Van Schouwenberg and Walinge 1973).
For measuring the plant phosphorous (P) the plant material was digested and then 10 ml of digested filtrate was taken in 100 ml volumetric flask and 10 ml of ammonium vanadomolybdate reagent was added and solution was diluted. The calibration curve was prepared from standards and the absorbance for P concentration was plotted and P concentration was determined from the calibration curve (Buresh et al. 1982).
The plant potassium (K) was determined by using the dry ashing method. For this purpose 1 g of ground plant material was taken in 50 ml porcelain crucibles and placed in cool muffle furnace and increased the temperature gradually to 550 °C, for 5 h after attaining 550 °C temperature. After 5 h, muffle furnace was shut off and left for cooling and crucibles were taken out carefully. The cooled ash was dissolved in 5 ml of 2 N HCl and mixed with plastic rod. After mixing for 20 min, the volume was brought up to 50 ml, and was filtered by using Whatman no. 42 filter paper. The K concentration was determined through colorimeter method (Chapman and Pratt 1961).
The Ca+2, Mg+2, Mn+2, Zn+2, Cu+2, and Fe+3 were determined with dry ashing. For this purpose 1 g of ground plant material was taken in 50 ml porcelain crucibles and placed in cool muffle furnace and increased the temperature gradually to 550 °C, for 5 h after attaining 550 °C temperature. After 5 h, muffle furnace was shut off and left for cooling and crucibles were taken out carefully. The cooled ash was dissolved in 5 ml of 2 N HCl and mixed with plastic rod. After 20 min, the volume was brought up to 50 ml with distilled water and mixed thoroughly and allowed to stand for 30 min. The Ca+2, Mg+2, Mn+2, Zn+2, Cu+2, and Fe+3 were determined by atomic absorption spectroscopy after filtration through Whatman no. 42 filter paper (Chapman and Pratt 1961).
2.6.3 Pb accumulation and uptake in plant
Pb contents in plants were measured by dry ashing. The metal contents in the extract were determined using the atomic absorption spectrophotometer (Page et al. 1982).
2.7 Statistical analyses
The data were statistically analyzed by Statistix 8.1 (Statistix, USA). Parametric statistics of ANOVA analysis was carried out to estimate the effect of Pb contamination on the soil microbial biomass, biomass C/N and C/P ratios, enzymes activity, soil extricable Pb, plant Pb uptake, pH, and plant growth and nutrients uptake. Mean separations were achieved using a least significant difference test at p < 0.05.
3 Results
3.1 Dynamics of soil microbial biomass under Pb contamination
Effect of Pb contamination on the dynamics f soil microbial biomass, i.e., MBC, MBN, and MBN is presented in the Figs. 1, 2, and 3. Data showed that maximum MBC, MBN and MBP contents (131.4, 16.4, and 7.8 mg kg−1) were found in control with no Pb treatment and minimum MBC, MBN, and MBP contents (23.5, 1.4, and 0.31 mg kg−1) were found at highest Pb level, i.e., 600 mg Pb kg−1 soil. Soil microbial biomass decreased within all Pb treatments when compared with the control, and this decrease in the soil microbial biomass increased with the increase in the Pb concentration. The incubation time, also played a vital role, Pb toxicity increased with the increased incubation time, therefore, minimum MBC, MBN, and MBP were observed at the 60th day. Different levels of Pb, i.e., 150, 300, 450, and 600 mg Pb kg−1 soil, significantly (P < 0.05) decreased the MBC (1.16-, 1.32-, 1.56-, and 5.59-fold), MBN (1.60-, 2.41-, 2.6-, and 11.7-fold), and MBP (1.75-, 2.50-, 3.88-, and 25.1-fold) compared with control.

Dynamics of soil microbial biomass carbon (MBC) under different Pb treatments and incubation times. The error bars represent standard error of the means

Dynamics of soil microbial biomass nitrogen (MBN) under different Pb treatments and incubation times. The error bars represent standard error of the means

Dynamics of soil microbial biomass phosphorous (MBP) under different Pb treatments and incubation times. The error bars represent standard error of the means
3.2 Dynamics of biomass C/N and C/P ratio under Pb contamination
Effect of Pb contamination on the dynamics of soil microbial biomass C/N and C/P ratios is shown in the Figs. 4 and 5. Data showed that minimum C/N and C/P ratios (8.01 and 16.8) were found in control with no Pb treatment and maximum C/N and C/P ratios (37.6 and 101) were found at highest Pb level, i.e., 600 mg Pb kg−1 soil. Soil microbial biomass C/N and C/P ratios increased within all Pb treatments when compared with the control, and this increase in the soil microbial biomass C/N and C/P ratios increased with the increase in the Pb concentration. The incubation time, also played a vital role, soil microbial biomass C/N and C/P ratios increased with the increased incubation time, therefore, maximum soil microbial biomass C/N and C/P ratios were observed at the 60th day. Different levels of Pb, i.e., 150, 300, 450, and 600 mg Pb kg−1 soil, significantly (P < 0.05) increased the soil microbial biomass C/N (1.48-, 1.82-, 2.13-, and 4.69-fold) and C/P ratios (1.50-, 1.90-, 2.5-, and 6.01-fold) compared with control.

Dynamics of soil microbial biomass carbon and nitrogen ratio (MBC/MBN) under different Pb treatments and incubation times. The error bars represent standard error of the means

Dynamics of soil microbial biomass carbon and phosphorous ratio (MBC/MBP) under different Pb treatments and incubation times. The error bars represent standard error of the means
3.3 Dynamics of soil enzymes activity under Pb contamination
Effect of Pb contamination on the dynamics of soil enzymes activity, i.e., dehydrogenase, phosphatase, and urease is presented in the Figs. 6, 7, and 8. Data showed that maximum soil enzymes activity, i.e., dehydrogenase, phosphatase, and urease (48.7 mg TPF kg−1 24 h−1, 23.5 mg PNP kg−1 h−1, and 7.5 mg NH4–N kg−1 2 h−1) were found in control with no Pb treatment and minimum dehydrogenase, phosphatase and urease activities (12.1 mg TPF kg−1 24 h−1, 2.5 mg PNP kg−1 h−1, and 0.81 mg NH4–N kg−1 2 h−1) were observed at highest Pb level, i.e., 600 mg Pb kg−1 soil. Soil enzymatic activity decreased within all Pb treatments when compared with the control, and this decrease in the soil enzymatic activity increased with the increase in the Pb concentration. The incubation time, also played a vital role, Pb toxicity increased with the increased incubation time, therefore, minimum dehydrogenase, phosphatase, and urease activities were observed at the 60th day. Different levels of Pb, i.e., 150, 300, 450, and 600 mg Pb kg−1 soil, significantly (P < 0.05) decreased the dehydrogenase (1.29-, 1.66-, 2.25-, and 4.02-fold), phosphatase (1.45-, 1.85-, 2.83-, and 19.4-fold), and urease (1.68-, 2.12-, 2.87-, and 9.25-fold) compared with control.

Dynamics of soil dehydrogenase activity under different Pb treatments and incubation times. The error bars represent standard error of the means

Dynamics of soil phosphtse activity under different Pb treatments and incubation times. The error bars represent standard error of the means

Dynamics of soil urease activity under different Pb treatments and incubation times. The error bars represent standard error of the means
3.4 Changes in extractable Pb in soil
The changes in extractable Pb were determined after every 15 days interval, after the application of different Pb treatments (Fig. 9). The data pertaining to the extractable Pb showed that it was below detection limit in control and was maximum (485.8 mg kg−1) in the highest Pb level, i.e., 600 mg Pb kg−1 soil. Soil extractable Pb increased within all Pb treatments when compared with the control. Nevertheless, this increase in the soil extractable Pb increased with the increase in Pb concentrations. The incubation time, also played a vital role, soil extractable Pb decreased with the increased incubation time, therefore, minimum soil extractable Pb was observed at the 60th day. The high levels of Pb, i.e., 300, 450, and 600 mg Pb kg−1 soil, significantly (P < 0.05) increased the soil extractable Pb contents (2.02-, 2.96-, and 3.86-fold) compared with low level of Pb treatment, i.e., 150 mg Pb kg−1 soil.

Dynamics of soil extractable Pb under different Pb treatments and incubation times. The error bars represent standard error of the means
3.5 Changes in soil pH
Effect of Pb contamination on the soil pH is presented in the Fig. 10. Data showed that maximum soil pH (7.91) was found in control with no Pb treatment and minimum soil pH (5.66) was found at the highest Pb level, i.e., 600 mg Pb kg−1 soil. Soil pH decreased within all Pb treatments when compared with the control. However, this decrease in the soil pH increased with the increase in Pb concentrations. The incubation time, also played a vital role, soil pH decreased with the passage of time, therefore, minimum soil pH was observed at the last day of the experiment, i.e., 60th day. Different levels of Pb, i.e., 150, 300, 450, and 600 mg Pb kg−1 soil, significantly (P < 0.05) decreased the soil pH (1.06-, 1.15-, 1.25-, and 1.40-fold) compared with control.

Dynamics of soil pH under different Pb treatments and incubation times. The error bars represent standard error of the means
3.6 Plant shoot and root, chlorophyll, and carotenoid contents at harvest
Effect of Pb contamination on the growth parameters of spinach plant, i.e., shoot and root length, shoot and root fresh weight, shoot and root dry weight, chlorophyll, and carotenoid contents at harvest is presented in the Fig. 11. Data showed that maximum shoot and root length (26.5 and 13.2 cm), shoot and root fresh weight (17.5 and 7.5 g), shoot and root dry weight (8.4 and 3.91 g), chlorophyll (5.1 mg), and carotenoid contents (9.2 μg) at harvest were found in control (with no Pb treatment) and minimum shoot and root length (12.2 and 5.3 cm), shoot and root fresh weight (7.4 and 2.79 g), shoot and root dry weight (2.15 and 1.12 g), chlorophyll (0.69 mg), and carotenoid contents (2.14 μg) at harvest were found at highest Pb level, i.e., 600 mg Pb kg−1 soil. Growth parameters of spinach plant, i.e., shoot and root length, shoot and root fresh weight and shoot, root dry weight, chlorophyll, and carotenoid contents decreased within all Pb treatments when compared with the control. Nevertheless, this decrease in the growth parameters of spinach plant increased with the increase in Pb concentrations. Different levels of Pb, i.e., 150, 300, 450, and 600 mg Pb kg−1 soil, significantly (P < 0.05) decreased the shoot length (1.06-, 1.18-, 1.42-, and 2.17-fold), root length (1.09-, 1.24-, 1.5-, and 2.54-fold), shoot fresh weight (1.08-, 1.22-, 1.49-, and 2.36-fold), root fresh weight (1.06-, 1.17-, 1.32-, and 2.69-fold), shoot dry weight (1.09-, 1.28-, 1.66-, and 3.90-fold), root dry weight (1.09-, 1.30-, 1.65-, and 3.50-fold), chlorophyll content (1.20-, 1.56-, 2.27-, and 5.60-fold), and carotenoid contents (1.1-, 1.38-, 1.94-, and 4.29-fold) compared with control.

Plant growth parameters under different Pb treatments. The error bars represent standard error of the means. SL shoot length, RL root length, SFW shoot fresh weight, RFW root fresh weight, SDW shoot dry weight, RDW root dry weight, Chl chlorophyll, Carot carotenoid, CK control. Units: SL and RL, centimeters; SFW, RFW, SDW, and RDW, grams; Chl, milligrams; and Carot, micrograms
3.7 Plant macronutrient contents in shoot
Effect of Pb contamination on the macronutrient contents, i.e., N, P, K, Ca, and Mg in the spinach shoot is presented in the Fig. 12. Data showed that maximum N (5.12%), P (1.86%), K (5.76%), Ca (1.45%), and Mg (0.78%) contents were found in control (with no Pb treatment), and minimum N (1.17%), P (0.48%), K (1.7%), Ca (0.22%), and Mg (0.14%) contents were found at highest Pb level, i.e., 600 mg Pb kg−1 soil. The macronutrient contents, i.e., N, P, K, Ca, and Mg in the spinach shoot decreased within all Pb treatments when compared with the control, and this decrease in the macronutrient contents increased with the increase in the Pb concentration. Different levels of Pb, i.e., 150, 300, 450, and 600 mg Pb kg−1 soil, significantly (P < 0.05) decreased the macronutrient contents N (1.07-, 1.24-, 1.69-, and 4.38-fold), P (1.12-, 1.41-, 1.62-, and 3.88-fold), K (1.08-, 1.21-, 1.40-, and 3.88-fold), Ca (1.15-, 1.40-, 1.86-, and 6.60-fold), and Mg (1.18-, 1.53-, 2.23-, and 5.57-fold) in the spinach shoot compared with control.

Macronutrint contents in plant shoot under different Pb treatments. The error bars represent standard error of the means
3.8 Plant macronutrient contents in root
Effect of Pb contamination on the macronutrient contents, i.e., N, P, K, Ca, and Mg in the spinach root is presented in the Fig. 13. Data showed that maximum N (2.67 mg kg−1), P (0.79%), K (2.49%), Ca (0.67%), and Mg (0.49%) contents were found in control (with no Pb treatment) and minimum N (0.9%), P (0.12%), K (0.54%), Ca (0.1%), and Mg (0.11%) contents were found at highest Pb level, i.e., 600 mg Pb kg−1 soil. The macronutrient contents, i.e., N, P, K, Ca, and Mg in the spinach root decreased within all Pb treatments when compared with the control. Different levels of Pb, i.e., 150, 300, 450, and 600 mg Pb kg−1 soil, significantly (P < 0.05) decreased the macronutrient contents N (1.06-, 1.14-, 1.27-, and 2.97-fold), P (1.16-, 1.44-, 2.02-, and 6.58-fold), K (1.07-, 1.18-, 1.51-, and 4.6-fold), Ca (1.19-, 1.53-, 2.16-, and 6.7-fold), and Mg (1.08-, 1.26-, 1.53-, and 4.45-fold) in the spinach root compared with control.

Macronutrint contents in plant root under different Pb treatments. The error bars represent standard error of the means
3.9 Plant micronutrient contents in shoot
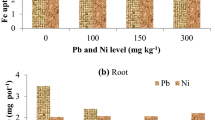
Effect of Pb contamination on the micronutrient contents, i.e., Zn, Cu, Fe, and Mn in the spinach shoot is presented in the Fig. 14. Data showed that maximum Zn (83.5 mg kg−1), Cu (25.1 mg kg−1), Fe (32.2 mg kg−1), and Mn (118.5 mg kg−1) contents were observed in control (with no Pb treatment) and minimum Zn (34.9 mg kg−1), Cu (6.78 mg kg−1), Fe (7.8 mg kg−1), and Mn (28.4 mg kg−1) contents were found at highest Pb level, i.e., 600 mg Pb kg−1 soil. The micronutrient contents, i.e., Zn, Cu, Fe, and Mn in the spinach shoot decreased within all Pb treatments when compared with the control and this decrease in micronutrient contents increased with the increase in the Pb concentration. Different levels of Pb, i.e., 150, 300, 450, and 600 mg Pb kg−1 soil, significantly (P < 0.05) decreased the micronutrient contents Zn (1.04-, 1.14-, 1.43-, and 2.39-fold), Cu (1.06-, 1.15-, 1.54-, and 3.70-fold), Fe (1.07-, 1.20-, 1.60-, and 4.13-fold), and Mn (1.14-, 1.4-, 1.99-, and 4.17-fold) in the spinach shoot compared with control.

Micronutrint contents in plant shoot under different Pb treatments. The error bars represent standard error of the means
3.10 Plant micronutrient contents in root
Effect of Pb contamination on the micronutrient contents, i.e., Zn, Cu, Fe, and Mn in the spinach root is presented in the Fig. 15. Data showed that maximum Zn (29.6 mg kg−1), Cu (9.5 mg kg−1), Fe (15.2 mg kg−1), and Mn (76.5 mg kg−1) contents were found in control (with no Pb treatment), and minimum Zn (9.7 mg kg−1), Cu (3.62 mg kg−1), Fe (4.7 mg kg−1), and Mn (18.7 mg kg−1) contents were found at highest Pb level, i.e., 600 mg Pb kg−1 soil. The micronutrient contents, i.e., Zn, Cu, Fe, and Mn in the spinach root decreased within all Pb treatments when compared with the control, and this decrease in the micronutrient contents increased with the increase in the Pb concentration. Different levels of Pb, i.e., 150, 300, 450, and 600 mg Pb kg−1 soil, significantly (P < 0.05) decreased the micronutrient contents Zn (1.08-, 1.22-, 1.49-, and 3.05-fold), Cu (1.07-, 1.20-, 1.41-, and 2.62-fold), Fe (1.09-, 1.25-, 1.54-, and 3.23-fold), and Mn (1.08-, 1.26-, 1.77-, and 4.09-fold) in the spinach root compared with control.

Micronutrint contents in plant root under different Pb treatments. The error bars represent standard error of the means
3.11 Plant Pb uptake in shoot and root
Pb uptake in shoot and root of spinach plant under different Pb concentrations, i.e., 0–600 mg Pb kg−1 soil is presented in the Fig. 16. The data pertaining to the plant Pb uptake in shoot and root showed that it was below detection limit in control, and was maximum (9.75 and 4.43 mg kg−1) in the highest Pb level, i.e., 600 mg Pb kg−1 soil. The plant Pb uptake in shoot and root increased within all Pb treatments when compared with the control. However, the plant Pb uptake in shoot and root increased with the increase in Pb concentrations. The high levels of Pb, i.e., 300, 450, and 600 mg Pb kg−1 soil, significantly (P < 0.05) increased the plant Pb uptake in shoot (1.68-, 2.54-, and 3.58-fold) and root (1.81-, 2.72-, and 4.38-fold) compared with low level of Pb treatment, i.e., 150 mg Pb kg−1 soil.

Plant Pb uptake in root and shoot under different Pb treatments. The error bars represent standard error of the means
4 Discussion
Results showed that soil microbial biomass, i.e., MBC, MBN, and MBP decreased within all Pb (150, 300, 450, and 600 mg Pb kg−1 soil) treatments with the passage of time (15–60 days) compared with the control (Figs. 1, 2, and 3). However, high Pb levels had more suppressive effect, therefore, highest Pb level, i.e., 600 mg Pb kg−1 soil significantly (P < 0.05) decreased the MBC, MBN, and MBP by 5.59-, 11.71-, and 25.1-fold, respectively at the end of the experiment, i.e., day 60. This might be due to response of highest Pb treatment (600 mg Pb kg−1 soil) was rapid and efficient as compared with the other treatments. Zeng et al. (2007) set an experiment at Hangzhou, China to see the effects of Pb contamination on microbial biomass in a greenhouse pot experiment and they concluded that microbial biomass significantly (P < 0.05) decreased with the increased Pb concentrations. Khan et al. (2010) conducted a study in the greenhouse for a period of 12 weeks at Beijing, China, and found that soil microbial biomass decreased significantly (P < 0.05) in the metal-amended samples, and decrease in the microbial biomass activity increased with the increase in Pb levels and incubation time. Bhattacharyya et al. (2008) conducted a study at West Bengal, India and found that microbial biomass had a significantly (P < 0.05) negative correlation with the increased metal concentrations.
The MBC/MBN and MBC/MBP ratios increased within all Pb (150, 300, 450, and 600 mg Pb kg−1 soil) treatments with the passage of time (15–60 days) compared with the control (Figs. 4 and 5). However, highest Pb level, i.e., 600 mg Pb kg−1 soil had 4.69- and 6.01-fold more MBC/MBN and MBC/MBP ratios, respectively at the end of the experiment, i.e., day 60. This might be with the increase in the level and toxicity of the applied Pb, the size of soil microbial community and C mineralization decreased, which ultimately enhanced the microbial biomass ratios. Moreover, microbial community structure changed accordingly due to increased metal stress in the soil, showing an increase in the fungal to bacterial ratio, as fungi tended to be more resistant to heavy metals than bacteria. Akmal et al. (2005) conducted an incubation experiment for a period of 56 days in Hangzhou, China, and found lowest microbial biomass C/N ratio in control and highest at 1,000 mg kg−1 Pb. Akmal and Jianming (2009) stated that microbial biomass ratios increased with the increased Pb contamination, and maximum microbial biomass ratios were observed at the highest Pb level, i.e., 1,000 mg kg−1 Pb at the last day of the incubation, i.e., after 60 days of heavy metal pollution.
In the current study, soil enzymes activity namely, dehydrogenase, phosphates, and urease decreased within all Pb (150, 300, 450, and 600 mg Pb kg−1 soil) treatments with the passage of time (from 15 to 60 days) compared with the control (Figs. 6, 7, and 8). Nevertheless, high Pb levels had more inhibitory effect, therefore, highest Pb level, i.e., 600 mg Pb kg−1 soil significantly (P < 0.05) decreased the dehydrogenase, phosphates, and urease by 4.02-, 9.40-, and 9.26-fold, respectively at the end of the experiment, i.e., day 60. This was due to high Pb level suppressed the soil microbial community and ultimately enzymes activity. Khan et al. (2010) found that highest reduction in the acid phosphatase (35.6%) and urease (36.6%) activity was found at the high Pb levels and after 2 weeks of incubation. Similarly, Zeng et al. (2007) used six Pb levels viz. 0 (control), 100, 300, 500, 700, and 900 mg kg−1 soil, and they concluded that dehydrogenase, phosphatase, and urease activities significantly (P < 0.05) decreased with the increased Pb concentration, and strong inhibitory influence was observed at and after 500 mg Pb kg−1 soil. Bhattacharyya et al. (2008) found that the soil enzymes namely, β-glucosidase, urease, phosphatase, and arylsulphatase activities had a significantly (P < 0.05) negative correlation with the increased metal concentrations.
The extractable Pb was below detection limit in control and showed an increase with the increased Pb levels, i.e., from 150 to 600 mg Pb kg−1 soil, compared with the control (Fig. 9). Hence, highest extractable Pb (5.87-fold) was found at the 600 mg Pb kg−1 soil. The extractable Pb decreased with the passage of time, i.e., 15–60 days, as a result 1.1-fold less extractable Pb was observed at the 60th day, than at the 15th day. This could be due to the poor mobility of Pb in soil and presence of spinach plants. Liao et al. (2007) after contaminating the soil up to 900 mg kg−1 by using Pb acetate, under Chinese cabbage (Brassica chinensis), found that, available Pb contents were particularly low instead of polluting the soil with higher quantities of Pb after 60 days of plant growth. Similarly, Tiemann et al. (1999) found less bioavailability of Pb than the total quantities and explained that this might be due to Alfalfa plant, which did bind the heavy metal and reduced its bioavailability.
Soil pH decreased within all Pb (150, 300, 450, and 600 mg Pb kg−1 soil) treatments with the passage of time (from 15 to 60 days) compared with the control (Fig. 10). However, highest Pb level, i.e., 600 mg Pb kg−1 soil have 1.40-fold low soil pH, at the end of the experiment, i.e., day 60, compared with other Pb treatments. This happened because the effect of the highest Pb treatment (600 mg Pb kg−1 soil) on the microbial and chemical properties was more drastic as compared with the other treatments. Chander et al. (2001) examined the effect of metals (Pb, Cu, and Zn) on chemical and biochemical properties of soil, and concluded that chemical (e.g., pH) and biochemical properties of soil decreased with the increased load and toxicity of the metals pollution. Wyszkowska and Kucharski (2000) investigated the effects of different levels of Pb, i.e., 0, 2, 4, and 6 cm3 kg−1 on the growth and development of triticale and biochemical properties of the brown soil of Olsztyn, Poland. The authors found that Pb contamination significantly decreased the chemical and biochemical properties of soil.
The growth parameters of spinach plant, i.e., shoot and root length, shoot and root fresh weight, shoot and root dry weight, chlorophyll, and carotenoid contents decreased within all Pb (150, 300, 450, and 600 mg Pb kg−1 soil) treatments, compared with control (Fig. 11). However, high Pb levels had more inhibitory effect, therefore, highest Pb level, i.e., 600 mg Pb kg−1 soil significantly (P < 0.05) decreased the growth parameters of spinach plant, i.e., shoot and root length, shoot and root fresh weight, shoot and root dry weight, chlorophyll, and carotenoid contents by 2.17-, 2.54-, 2.36-, 2.69-, 3.90-, 3.50-, 5.60-, and 4.29-fold, respectively at the end of the experiment, i.e., day 60. The reason of this decline might be Pb toxicity adversely affected the nutrient uptake ability, and chloroplast, hence mechanism of photosynthesis in plants and due to the inhibition of plant photosynthesis all growth parameters decreased significantly. Zeng et al. (2007) after using six Pb levels, i.e., 0 (control), 100, 300, 500, 700, and 900 mg kg−1 soil, concluded that rice biomass and chlorophyll content decreased gradually with the increased Pb concentration and this decrease in plant biomass was highest at 500 to 900 mg kg−1 Pb. Wyszkowska and Kucharski (2000) used four levels of Pb, i.e., 0, 2, 4, and 6 cm3 kg−1 in an experiment at Olsztyn, Poland, and found that Pb contamination significantly decreased the growth and development, and chlorophyll contents of the tested crop, i.e., triticale plant. Hussain et al. (2006) in a pot experiment used two levels of Pb, i.e., 20 and 40 mg l− and concluded that application of Pb metal significantly reduced all growth attributes, i.e., shoot and root length, shoot and root fresh weight, shoot and root dry weight, chlorophyll, and carotenoid contents of mash bean. Similarly, Rout et al. (2001) reported a significant decrease in the carotenoid content of the plants due to heavy metal toxicity.
The macronutrients, i.e., N, P, K, Ca, and Mg in the spinach shoot and root decreased within all Pb (150, 300, 450, and 600 mg Pb kg−1 soil) treatments (Figs. 12 and 13). However, high Pb levels had more inhibitory effect, therefore, highest Pb level, i.e., 600 mg Pb kg−1 soil significantly (P < 0.05) decreased the macronutrients, i.e., N, P, K, Ca, and Mg by 4.38-, 3.88-, 3.88-, 6.60-, and 5.57-fold in the spinach shoot and 2.97-, 6.58-, 4.6-, 6.70-, and 4.45-fold in the spinach root, respectively. This might be due to Pb blocked the plants nutrient elements uptake and because of the blockage low macronutrients were observed in plant shoot and root. Paivoke (2002) after using two levels of Pb, i.e., 0.5 and 9.4 mmol Pb acetate kg−1 dwt soil in a pot experiment of 21 days, found that N, P, K, Mg, Fe, Na, and S contents were negatively correlated with soil Pb. High concentration of Pb in the soil environment significantly imbalanced the internal nutrients in the growing plant, and in most cases Pb blocked the entry of cations, i.e., K+, Ca+2, Mg+2, Mn+2, Zn+2, Cu+2, and Fe+3 (Sharma and Dubey 2005). Walker et al. (1977) examined the effects of two levels of Pb, i.e., 125 and 250 Pb g−1 soil on mineral nutrient contents of corn plants for 24 days in potted soil containing loamy sand in a greenhouse and found that Pb significantly decreased the uptake of K, Ca, Mg, Fe, and NO3- in corn plant. Kibria et al. (2009) investigated the effects of six levels of Pb, i.e., 0, 20, 40, 60, 80, and 100 mg kg−1 soil on growth and mineral nutrition of Amaranthus gangeticus L. and Amaranthus oleracea L. in a pot experiment and found that Pb application in soil significantly decreased N, P, Ca, Zn, Fe, and Mn in shoots and roots.
The micronutrients, i.e., Zn, Cu, Fe, and Mn in the spinach shoot and root decreased within all Pb (150, 300, 450, and 600 mg Pb kg−1 soil) treatments (Figs. 14 and 15). However, high Pb levels had more inhibitory effect therefore highest Pb level, i.e., 600 mg Pb kg−1 soil significantly (P < 0.05) decreased the micronutrients, i.e., Zn, Cu, Fe, and Mn by 2.39-, 3.70-, 4.13-, and 4.17-fold in spinach shoot and 3.05-, 2.62-, 3.23-, and 4.09-fold in the spinach root. This decrease in micronutrient cations was due to the reason that Pb is a divalent cation-like micronutrient cations, and the ionic radii of Pb are also similar to that of micronutrient cations, as a result Pb imbalanced the mineral nutrients. Moreover, Pb blocked the uptake of nutrients, and retarded the root development of spinach and because of the blockage and poor root development the cations uptake was strongly inhibited. Gopal and Rizvi (2008) conducted an experiment to evaluate detrimental effects of two levels of Pb, i.e., 0.1 and 0.5 mM on growth, metabolism, and nutrients uptake of Radish (Raphanus sativus), under glasshouse conditions for 65 days; they found that Pb contamination reduced the concentration of Fe and S in the shoot of Radish. Sarfraz et al. (2007) examined the effect of metal concentrations on yield of rice and concluded that Zn, Cu, Fe, and Mn were found in straw and the nutrients concentration decreased with increased metal concentration in straw. Paivoke (2002) exposed the pea (Pisum sativum L.) plant to Pb, for 21 days in a greenhouse and found nutrients imbalance in pea plant and nutrients contents, e.g., Fe, Na, and S decreased with the increased Pb concentrations. Similarly, Sharma and Dubey (2005) after reviewing stated that high concentration of Pb imbalanced the internal nutrients in the growing plants and blocked the entry of cations, i.e., K+, Ca+2, Mg+2, Mn+2, Zn+2, Cu+2, and Fe+3.
The data related to the plant Pb uptake in shoot and root showed that it was below detection limit in control, and increased with the increase in Pb level, i.e., from 150 to 600 mg Pb kg−1 soil (Fig. 16). Hence, highest Pb uptake in shoot (3.58-fold) and root (4.38-fold) was found at the 600 mg Pb kg−1 soil. The data showed that Pb accumulation in plant increased with the increased Pb levels. Michalska and Asp (2001) observed the accumulation of Pb in spinach roots increased with the increased Pb levels. Paivoke (2002) showed that pea (P. sativum L.) had highest Pb accumulation in shoot and root at the higher Pb treatments. Sarfraz et al. (2007) conducted an experiment to see the metal accumulation in the rice plant, and stated that metal accumulation in the rice straw and grains increased with the increased level of metal concentration.
5 Conclusions
The results reported permit the conclusion that heavy metal pollution is indeed an alarming threat for soil microbiological health and fertility and ultimately to crop production. The Pb contamination had a strong inhibitory effect on both soil and plant growth parameters, and the degree of the influence increased with the increased Pb concentration and incubation time, deducing that Pb threshold is strongly associated with the extent of Pb concentration and time to accumulate. High soil contamination by Pb could be a potential risk to soil microbiological index and agricultural crops or vegetables, and the interactions of plant–soil–microbes, in the long run. When the level of Pb treatments increased to 600 mg kg−1, ecological risk existed evidently to both soil microbiological activities and plant growth. Soil microbial biomass, enzymatic activities, pH, spinach physiological indices, and spinach biomass might be sensitive indicators reflecting environmental stress in the soil system. However, more long- and short-term research is needed to assess the ecological risk of Pb contamination under field conditions.
Change history
14 March 2019
This article has been retracted by the Editors-in-Chief.
References
Agricultural Statistics of Pakistan (2010–2011) Government of Pakistan, Ministry of Food, Agriculture and Livestock (Economic trade and investment wing). Printing Corporation of Pakistan Press, Islamabad, p. 280
Akmal M, Jianming X (2009) Microbial biomass and bacterial community changes by Pb contamination in acidic soil. J Agric Biol Sci 1:30–37
Akmal M, Jianming X, Zhaojun L, Haizhen W, Huaiying Y (2005) Effects of lead and cadmium nitrate on biomass and substrate utilization pattern of soil microbial communities. Chemosphere 60:508–514
Alef K, Nannipieri P (1995) Methods in applied soil microbiology and biochemistry. Academic, San Diego, p 490
Anderson JM, Ingram JSI (1993) Tropical soil biology and fertility. CAB International, Wallingford, pp 50–79
Bhattacharyya P, Chakrabarti K, Chakraborty A, Tripathy S, Powell MA (2008) Fractionation and bioavailability of Pb in municipal solid waste compost and Pb uptake by rice straw and grain under submerged condition in amended soil. Geosci J 12:41–45
Bindler R (2011) Contaminated lead environments of man: reviewing the lead isotopic evidence in sediments, peat, and soils for the temporal and spatial patterns of atmospheric lead pollution in Sweden. Environ Geochem Health 33:311–329
Brookes PC, Landman A, Pruden G, Jenkinson DS (1985) Chloroform fumigation and the release of soil nitrogen: a rapid direct extraction method to measure microbial biomass nitrogen in soil. Soil Biol Biochem 17:837–842
Buresh RJ, Austin ER, Craswell ET (1982) Analytical methods in N-15 research. Fert Res 3:37–62
Chander K, Dyckmans J, Hoeper H, Joergensen RG, Raubuch M (2001) Long term effects on soil microbial properties of heavy metals from industrial exhaust deposition. J Plant Nutr Soil Sci 164:657–663
Chapman HD, Pratt PF (1961) Methods of analysis for soils, plants and water. University of California, Berkeley
Eldor P (2007) Soil microbiology, ecology, and biochemistry. In: Eldor P (ed) Tercera. Academic, Chennai
Ensink JHJ, Simmons RW, van der Hoek W (2004) Wastewater use in Pakistan: the cases of Haroonabad and Faisalabad. In: Scott CA, Faruqui NI, Raschid L (eds) Wastewater use in irrigated agriculture, confronting the livelihood and environmental realities. CAB International, Wallingford, pp 91–99
Filip Z (2002) International approach to assessing soil quality by ecological related biological parameters. Agri Ecosyst Environ 88:169–174
Gonzales-Quiñones V, Stockdale EA, Banning NC, Hoyle FC, Sawada Y, Wherrett AD, Jones DL, Murphy DV (2011) Soil microbial biomass: interpretation and consideration for soil monitoring. Soil Res 49:287–304
Gopal R, Rizvi AH (2008) Excess lead alters growth, metabolism and translocation of certain nutrients in radish. Chemosphere 70:1539–1544
Hassan W (2013) C and N mineralization and dissolved organic matter potentials of two contrasting plant residues: effects of residue type, moisture and temperature. Acta Agr Scand B-S P. doi:10.1080/09064710.2013.846398
Hassan W, Akmal M, Muhammad I, Younas M, Zahaid KR, Ali F (2013a) Response of soil microbial biomass and enzymes activity to cadmium (Cd) toxicity under different soil textures and incubation times. Aust J Crop Sci 7:674–680
Hassan W, Chen W, Huang Q, Mohamed I (2013b) Microcalorimetric evaluation of soil microbiological properties under plant residues and dogmatic water gradients in Red soil. Soil Sci Plant Nut. doi:10.1080/00380768.2013.845735
Hassan W, Chen W, Cai P, Huang Q (2013c) Oxidative enzymes, the ultimate regulator: implications for factors affecting their efficiency. J Environ Qual 42:1–12
Hesse PR (1972) A textbook of soil chemical analysis. Chemical Publishing Co. Inc, New York
Hussain M, Ahmad MSA, Kausar A (2006) Effect of lead and chromium on growth, photosynthetic pigments and yield components in mash bean [Vigna mungo (L.) Hepper]. Pak J Bot 38:1389–1396
Jackson ML (1973) Soil chemical analysis. Prentice Hall of India Pvt Ltd, New Delhi, p 498
Kandeler E, Gerber H (1988) Short-term assay of soil urease activity using colorimetric determination of ammonium. Biol Fertil Soils 6:68–72
Kandeler E, Mosier A, Morgan J, Milchunas D, King J, Rudolph S, Tscherko D (2006) Response of soil microbial biomass and enzyme activities to the trescient elevation of carbon dioxide in a semi-arid grassland. Soil Biol Biochem 38:2448–2460
Khan S, Cao Q, Zheng YM, Huang YZ, Zhu YG (2008) Health risks of heavy metals in contaminated soils and food crops irrigated with wastewater in Beijing, China. Environ Pollut 152:686–692
Khan S, Hesham A, Qiao M, Rehman S, He JZ (2010) Effects of Cd and Pb on soil microbial community structure and activities. Environ Sci Pollut Res 17:288–296
Kibria MG, Islam M, Osman KT (2009) Effects of lead on growth and mineral nutrition of Amaranthus gangeticus L. and Amaranthus oleracea L. Soil Environ 28:1–6
Komarek M, Ettler V, Chrastny V, Mihaljevic M (2008) Lead isotopes in environmental sciences: a review. Environ Int 34:562–577
Liao M, Chen C, Zeng L, Huang C (2007) Influence of lead acetate on soil microbial biomass and community structure in two different soils with the growth of Chinese cabbage (Brassica chinensis). Chemosphere 66:1197–1205
Lichtenthaler HK, Wellburn AR (1983) Determination of total carotenoids and chlorophyll ‘a’ and ‘b’ of leaf extracts in different solvents. Biochem Soc Trans 11:591–592
Majid NM, Islam MM, Rauf RA (2012) Evaluation of Jelutong (Dyera cotulata) as a phytoremediator to uptake copper (Cu) from contaminated soils. Aust J Crop Sci 6:369–374
Michalska M, Asp A (2001) Influence of lead and cadmium on growth, heavy metal uptake and nutrient concentration of three lettuce cultivars grown in hydroponic culture. Comm Soil Sci Plant Anal 32:571–583
Mishra S, Srivastava S, Tripathi RD, Kumar R, Seth CS, Gupta DK (2006) Lead detoxification by coontail (Ceratophyllum demersum L.) involves induction of phytochelatins and antioxidant system in response to its accumulation. Chemosphere 65:1027–1039
Murtaza G, Ghafoor A, Qadir M, Owens G, Aziz MA, Zia MH, Saifullah (2010) Disposal and use of sewage on agricultural lands in Pakistan: a review. Pedosphere 20:23–34
Nannipieri P, Badalucco L (2003) Biological processes. In: Bembi DK, Nieder R (eds) Processes in the soil-plant system. Modelling concepts and applications. The Haworth Press, Binghamton
Nannipieri P, Ascher J, Ceccherini MT, Landi L, Pietramellara G, Renella G (2003) Microbial diversity and soil functions. Eur J Soil Sci 54:655–670
Nasreen S (2006) Monitoring of surface water, ground water, air and soil in Peshawar basin against time the third dimension. (Unpublished) Ph.D. thesis, Centre of Excellence in Geology, University of Peshawar, Pakistan
Öhlinger R (1996) Biomass-P by fumigation–extraction technique. In: Schinner F, Ohlinger R, Kandeler E, Margesin R (eds) Methods in soil biology. Springer, Berlin, pp 62–64
Olsen SR, Sommers LE (1982) Phosphorus. In: Page AL (ed) Methods of soil analysis. Agronomy no. 9, Part 2. Chemical and microbiological properties, 2nd edn, ASA, Madison, WI, USA. pp. 403–430
Omirou M, Rousidou C, Bekris F, Papadopoulou KK, Mekissoglou-Spiroudi U, Ehaliotis C, Karpouzas G (2011) The impact of biofumigation and chemical fumigation methods on the structure and function of the soil microbial community. Microbial Ecol 61:201–213
Page AL, Miller RH, Keeney DR (1982) Methods of soil analysis-chemical and microbiological properties. Part 2. 2nd edn. Amer Soc Agron (ASA) no. 9, Madison, Wisconsin
Paivoke AEA (2002) Soil lead alters phytase activity and mineral nutrient balance of Pisum sativum. Environ Exp Bot 48:61–73
Rantalainen ML, Torkkeli M, Strömmer R, Setälä H (2006) Lead contamination of an old shooting range affecting the local ecosystem-a case study with a holistic approach. Sci Total Environ 69:99–108
Reimann C, Flem B, Fabian K, Birke M, Ladenberger A, Négrel P, Demetriades J, Hoogewerff A, The GEMAS Project Team (2012) Lead and lead isotopes in agricultural soils of Europe—the continental perspective. Appl Geochem 27:532–542
Robinson BH, Bischofberger S, Stoll A, Schroer D, Furrer G, Roulier S et al (2008) Plant uptake of trace elements on a Swiss military shooting range: uptake pathways and land management implications. Environ Pollut 153:668–676
Rout GR, Samantaray S, Das P (2001) Aluminum toxicity in plants: a review. Agronomie 21:3–21
Salk A, Arin L, Deveci M, Polat S (2008) Special vegetable production. Onur Press, Tekirdag, p 488
Sarfraz MS, Mehdi M, Hassan G, Abbas ST (2007) Metal contamination in nullah dek water and accumulation in rice. Pedosphere 17:130–136
Sharma P, Dubey RS (2005) Lead toxicity in plants. Braz J Plant Physiol 17:35–52
Tandy S, Schulin R, Nowack B (2006) The influence of EDDS on the uptake of heavy metals in hydroponically grown sunflowers. Chemosphere 62:1454–1463
Tiemann KJ, Gardea TJ, Gamez G, Dokken K, Sias S (1999) Use of X-ray absorption spectroscopy and esterification to investigate Cr(III) and Ni(II) ligands in alfalfa biomass. Environ Sci Technol 33:150–154
Van Schouwenberg JCH, Walinge I (1973) Methods of analysis for plant material. Agricultural University Wageningen, Wageningen, The Netherlands
Walker WM, Miller JE, Hassett JJ (1977) Effect of lead and cadmium upon the calcium, magnesium, potassium and phosphorus concentration in young corn plants. Soil Sci 124:145–151
Walkley AJ, Black TA (1934) An estimation of the Degt. Jarett method for determining soil organic matter and a proposed modification of the chromic acid titration. Soil Sci 37:29–38
Waseem K, Nadeem MA (2001) Enhancement of spinach production by varying sowing dates, row spacing and frequency of cuttings. Online J Biol Sci 1:902–904
Wu J, Joergensen RG, Pommerening B, Haussod R, Brookes PC (1990) Measurement of soil microbial biomass C by fumigation–extraction—an automated procedure. Soil Biol Biochem 22:1167–1169
Wyszkowska J, Kucharski J (2000) Biochemical properties of soil contaminated by petrol. Polish J Environ Stud 9:479–485
Zeng LS, Liao M, Chen CL, Huang CY (2007) Effects of lead contamination on soil enzymatic activities, microbial biomass and rice physiological indices in soil–lead–rice (Oryza sativa L.) system. Ecotoxic Envieron Saf 67:67–74
Author information
Authors and Affiliations
Corresponding author
Additional information
Responsible editor: Zucong Cai
This article has been retracted by the Editors-in-Chief. After a thorough investigation carried out by the Higher Education Commission of Pakistan and Muhammad Nawaz Sharif University of Agriculture, Pakistan, it was found that one of the co-authors, Mr. Waseem Hassan, plagiarized sections of this article from Mr. Idrees Haider's masters' thesis that was published in 2009 at PMAS Arid Agriculture University, Rawalpindi, Pakistan.
About this article
Cite this article
Hassan, W., David, J. RETRACTED ARTICLE: Effect of lead pollution on soil microbiological index under spinach (Spinacia oleracea L.) cultivation. J Soils Sediments 14, 44–59 (2014). https://doi.org/10.1007/s11368-013-0802-3
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s11368-013-0802-3