
Abstract
Purpose
There is a growing concern regarding the effects of endocrine disrupting compounds on humans and wildlife. Since some of them have the ability to bind to particulate matter, high concentrations can be reached in sediments of surface waters. The objective of this study was an assessment of 50 sediment samples from predominantly small rivers in the German federal state of Hesse with emphasis on endocrine activity.
Materials and methods
The freshwater mudsnail Potamopyrgus antipodarum was exposed to whole sediments for 28 days with a subsequent evaluation of embryo production. Additionally, in vitro screens with sediment eluates for the detection of estrogenic (yeast estrogen screen, YES) and androgenic (yeast androgen screen, YAS) activities were conducted. Biotest results were compared with chemical analytical data (metals, polycyclic aromatic hydrocarbons, polychlorinated biphenyls, organotin compounds), which were available for about half of the studied sites.
Results and discussion
Seventy-eight percent of the sediments caused a significantly increased reproduction in the Potamopyrgus test, 66% were active in YES, and 68% were active in YAS. More than half of the sediments caused an increased reproduction in vivo and concurrently showed an estrogenic activity in vitro. No significant correlations could be established between biotest results and chemical analytical data.
Conclusions
The results indicate that sediment contamination with endocrine active compounds is widespread, plays an important role in aquatic ecosystems, and should be the subject of further ecotoxicological investigation.
Similar content being viewed by others

Explore related subjects
Discover the latest articles, news and stories from top researchers in related subjects.Avoid common mistakes on your manuscript.
1 Introduction
Among environmental chemicals, endocrine disrupting compounds (EDCs) have gained more and more attention in recent years. They can influence the reproduction of organisms and thus are able to endanger the survival of a population in the field. For this reason, they have important ecological relevance and have become a subject of major concern among scientists in recent years (Sumpter 2005; LeBlanc 2007; Oehlmann et al. 2007).
Two of the best known cases of endocrine disruption in wildlife are (i) the formation of imposex in female prosobranch snails in tributyltin (TBT) polluted areas (Oehlmann et al. 1998; Santos et al. 2005; Wirzinger et al. 2007) and (ii) the demasculinization of male or "super-feminization" of female alligators in Lake Apopka in Florida, USA, associated with a pesticide spill as well as several decades of extensive agricultural activity around the lake and related pesticide and nutrient runoff (Guillette et al. 1994).
EDCs are a heterogeneous group of compounds, including both synthetic (e.g. alkylphenols, phthalates, bisphenol A, polychlorinated biphenyls (PCBs), 17α-ethinylestradiol) and natural compounds (e.g. estrone, 17β-estradiol, phytoestrogens). They are widely distributed and can be detected all over the world—even far from emission sources—due to long-range transport in air and water. EDCs interfere with endocrine systems by blocking, mimicking, or altering hormones and their signaling pathways. Since hormones act even in small doses, low environmental concentrations of endocrine active compounds may be sufficient to influence the function of the endocrine system.
Molluscs have often been proposed as suitable models to assess endocrine disruption (Oehlmann et al. 2000, 2007; Jobling et al. 2004; Ketata et al. 2008). The New Zealand mudsnail Potamopyrgus antipodarum (Hydrobiidae, Mollusca) used in this study is assumed to be a promising model organism for such investigations (Matthiessen 2008), since it responded with increased reproduction when exposed to (xeno)estrogens in various studies (Duft et al. 2003a; Jobling et al. 2004; Giudice and Young 2010). Due to an ovoviviparous reproduction, the embryos in the brood pouch can be easily counted (Duft et al. 2007).
In this study P. antipodarum was employed to investigate the estrogenic activity of sediment samples collected at 50 sampling sites in the German federal state of Hesse (see “Supplementary Material”). Since sediments are a habitat and a food source for various benthic animals and these benthic animals are in turn a major food source for other ecologically relevant trophic levels, an ecotoxicological assessment of sediments is an essential aspect for the evaluation of the ecological status of surface waters. Compounds can bind to sediment particles, whereby accumulated substances can achieve much higher concentrations than in the water phase (Peck et al. 2004). Additionally, sediment-bound substances can be remobilized into the water column under specific conditions such as changes in physico-chemical parameters or during flood events (Rubinos et al. 2010). Thus, they are both sinks and sources for environmental pollutants.
In addition to chemical analyses (to measure contamination) and in situ biological assessments (to investigate benthic community alterations), bioassays (to observe toxic effects on organisms) are an important tool to determine the quality of sediments (Chapman 1990). An advantage of bioassays is the investigation of the effects of all compounds occurring in the sediment, while analytical analyses examine only a limited number of substances. Moreover, in bioassays only bioavailable compounds which are able to enter the body are considered. Another advantage of exposure to whole sediments is the possibility of measuring interactive effects of complex contaminant mixtures containing compounds with antagonistic or additive effects.
The aim of this study was to evaluate the quality of sediments from 50 sampling sites in Hesse with the emphasis on endocrine contamination. For this purpose, a bioassay with P. antipodarum and in vitro assays (yeast estrogen screen (YES) and yeast androgen screen (YAS)) with sediment eluates were conducted. After a 28-day exposure period of the snails to whole sediments in the laboratory, mortality and reproduction success were determined.
2 Materials and methods
2.1 Study area and study sites
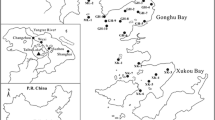
The study area extends throughout Hesse, a federal state in the western part of Germany (Fig. 1). The 50 study sites are located in 44 surface waters, mainly small rivers with a width of about 1–3 m (see “Supplementary Material”). They were proposed by the Hessian Agency for the Environment and Geology (HLUG) and cover a wide range of qualities regarding macroinvertebrate communities and chemical contamination. The southern part of the study area is one of the most densely populated and economically strongest regions of the country. Sampling site 1, which is situated in a nature reserve, serves as a reference site. Its ecological status was evaluated as very good with regard to assessments of fish, diatom, macrophyte, and benthic macroinvertebrate communities (HMUELV 2009). Chemical analytical investigations confirmed the largely uncontaminated state of this site (M. Heß, personal communication). Upstream of the sampling site the river does not receive wastewater treatment plant effluents and is not impacted by other sources of contamination such as agriculture.

Map of the study area and sampling sites
2.2 Sediment sampling procedure
At each site, samples were collected over a 10-m section from the upper 3 cm of the sediment in May 2007. A shovel was used to take the samples, whereby fine particles, which swirled within the water during this process, were collected with a small container. Sediments were collected in glass vessels, which were pre-cleaned with deionized water, ethanol, and acetone and heated to 150°C for sterilization purposes before use. Sediment samples were transported cooled (10°C) and in darkness to the laboratory, where they were stored frozen (−23°C) until the test start. Thereby, the infauna was eliminated, samples were preserved, and modification of substances was minimized.
2.3 Physical characterization of sediment samples
Grain size analysis was performed by fractionation of a dry sediment sample (ca. 100 g) through a series of sieves (mesh sizes: 20, 63, 125, 250, 500, 630, 1000, 2000, 4000 μm) and weighing each sediment fraction (DIN 18123–5 1996). The amount of organic matter was determined by loss on ignition (LOI: % of dry weight) using an oven (Carbolite, Ubstadt-Weiher), according to DIN 38414-S3 (1985). Dry sediment samples (about 5 g) were heated at 550°C for 60 min followed by weighing. The samples were heated for a further 30 min, and this step was repeated to constant weight. Since there is a significant positive correlation between LOI and total organic carbon (TOC) content (Ankley et al. 1994), LOI is also a measure for TOC.
2.4 Bioassay with P. antipodarum
The animals used in the experiment were taken from an in-house breeding stock, which was built up with species collected from the Emsbach creek (Limburg-Lindenholzhausen, Germany; 50°22′41.46′′N, 8°8′14.32′′E) in July 2007. Until tests start 8 months later, snails were kept in glass aquaria at 16 ± 1°C, a light/dark period of 16:8 h, and in aerated artificial freshwater (deionized water; pH 7.9–8.4 adjusted with 1 N NaOH and 1 N HCl; conductivity 770 μS cm−1 adjusted with TropicMarin® salt (Tropic Marin, Wartenberg/Angersbach, Germany)). They were fed ad libitum with finely ground TetraPhyll® (Tetra Werke, Melle, Germany) three times a week.
The experiment was conducted in a climate room at culture conditions in spring 2008. Test vessels (1-L glass beakers covered with plastic caps) were filled with well-homogenized native sediments up to a height of 1 cm before 800 mL of artificial water was added. The water was aerated via glass pipettes, and deionized water was added after evaporation several times a week. Additionally, controls with artificial sediment (97% quartz sand (quick-mix, Osnabrück, Germany) and 3% leaves of Alnus glutinosa) were used. Leaves were collected at a site situated in a rural region with no known sources of contamination. They were thoroughly washed with deionized water, air dried, and finely ground before being mixed with the quartz sand. Each sediment sample and the control were replicated twice. Test vessels were arranged randomly.
After a 1-day equilibration period, 22 snails with a shell height of 3.3–3.9 cm were inserted into each vessel. Preliminary tests showed that snails of this size produce similar numbers of embryos. During the test, snails were fed with a fish food suspension (TetraPhyll®, 0.25 mg per animal) three times a week.
After 28 days, the snails were picked out of the sediment, and mortality was recorded. Animals were put into small tubes, deep-frozen with liquid nitrogen, and stored at −20°C until preparation. Previous tests showed that freezing does not influence the test results. In order to assess the reproductive success of the snails, shells were cracked with a small pincer, and the brood pouch was opened with tweezers. Afterwards the number of embryos, differentiated in shelled and unshelled, was determined. The analysis was conducted blind without knowledge of the treatment. Embryo number was investigated for 20 snails per replicate (40 snails per sediment) or a reduced number in case of increased mortality. Reproduction was not evaluated for a study site if mortality exceeded 20%, since at this point sediments exhibit a toxic effect on test organisms which also affected reproduction.
Physico-chemical water parameters were measured at the beginning and at the end of the experiment in each test vessel (temperature, conductivity, pH, oxygen saturation, ammonium, nitrite), in one replicate per sediment (nitrate) or in random replicates (total hardness). Additionally, temperature, conductivity, pH, ammonia, and nitrite were measured after 14 days of exposure.
2.5 Sediment elution
Samples of homogenized, freeze-dried sediments (50 g) were extracted by sonication (15 min; Sonorex RK 52 H, Bandelin electronic, Berlin, Germany) with 100 mL water. During sonication the sediment was stirred several times. Laboratory tap water was used, since this was the least contaminated with endocrine active compounds in preliminary tests among different tested waters. The eluates were then centrifuged for 5 min at 14,500 rpm, and the supernatant was analyzed in the yeast screens.
2.6 Yeast estrogen screen and yeast androgen screen
The genetically modified yeast cells used in the YES contain the human estrogen receptor alpha (ERα) as well as an expression plasmid carrying the reporter gene lacZ and an estrogen responsive element. When a ligand binds to the estrogen receptor, the reporter gene lacZ is expressed and produces β-galactosidase. This enzyme metabolizes an added chromogenic substrate resulting in a photometrically measurable color change. The YAS follows the same principle, differing only in the used modified yeast cells expressing the human androgen instead of the estrogen receptor and the corresponding responsive element.
The YES was conducted according to Routledge and Sumpter (1996) and Rutishauser et al. (2004), the YAS according to Sohoni and Sumpter (1998). Modifications described by Wagner and Oehlmann (2009) were applied to test water samples.
Seventy-five microliters of eluate samples (corresponding to 37.5 mg of sediment) were added to 96-well microtiter plates in eight replicates. Laboratory tap water served as a negative control. In each assay a standard curve with 17β-estradiol (YES) or testosterone (YAS) was included as well as a blank without yeast cells. Incubation occurred for 24 h at 30°C and 750 rpm.
Measurements and data analysis, including calculation of estradiol equivalents (EEQs) or testosterone equivalents (TEQs), were conducted as described previously (Wagner and Oehlmann 2009). Results are expressed as nanograms EEQ or TEQ per kilogram sediment. If eluates were toxic to yeast cells (20% reduced cell density in comparison to negative control), equivalents were not calculated, and estrogenicity and androgenicity were not evaluated. Limit of quantification (LOQ) was calculated as: LOQ = mean + 3 times the SEM.
2.7 Chemical analytical data
Chemical analytical data of the suspended particulate matter including metals, polychlorinated biphenyls (PCBs), polycyclic aromatic hydrocarbons (PAHs), and organotin compounds (OTCs); Table 1) were provided by HLUG for about half of the study sites. The data are averages of several measurements between 2005 and 2008.
Metals, except for mercury, were analyzed according to DIN EN ISO 17294–2 (2005) using inductively coupled plasma–mass spectrometry (ICP–MS). Atomic absorption spectrometry (AAS) was used for mercury analysis. PCB analysis was conducted according to DIN 38414-S20 (1996). After soxhlet extraction with toluol/acetone of a freeze dried sample, analysis was performed using high resolution gas chromatography–high resolution mass spectrometry (HRGC–HRMS). For PAH analysis, samples were prepared in the same way as for PCB analysis (freeze-drying, soxhlet extraction with toluol/acetone). Analytical investigations were performed using HRGC–MS. For the analysis of OTCs, extraction with hexane/acetone was followed by analysis using HRGC–MS.
Chemical data were compared with biotest results. In order to determine the contamination at a study site, quotients of substance concentrations and the corresponding quality limit values were added. Quotients of ≤ 1 were ignored, since substances represent, by definition, no hazard if limit values are not exceeded. Quality limit values are concentrations of a particular pollutant, which should not be exceeded in the environment by reasons of health and environmental protection. The different references of quality limit values result from their different availability. Contamination levels are listed in Table 2.
2.8 Statistical analyses
Data analyses were performed using GraphPad Prism®, version 5.00 (San Diego, CA, USA). In order to check for significant differences in the reproduction between the artificial control sediment and the field sediments, parametric tests were performed. Since all sampling sites were completely distinct from each other and represent independent samples, a Student's t-test was used to assess whether the means of the control and the various sediments were statistically significantly different. The data of the two replicates were pooled, whereby a sample size of n = 40 was achieved. In the case of the yeast screens, the non-parametric Mann–Whitney U-test was applied to analyze significant differences between the negative control and sampling sites, as data did not pass the normality test (D'Agostino and Pearson omnibus normality test). The non-parametric Spearman correlation analysis was employed to study possible correlations between EEQs, TEQs, and embryo numbers as well as between chemical analytical data and biotest results. For correlations as well as significant differences, three levels were considered statistically significant: p < 0.05 (★), p < 0.01 (★★), and p < 0.001 (★★★).
3 Results
3.1 Physical characterizations of sediment samples
Mean particle size of the sampled sediments ranged from 0.017 to 2.44 mm, and loss on ignition (LOI) from 0.395 to 29.5% (for details see “Supplementary Material”).
3.2 Biotest with P. antipodarum
3.2.1 Mortality
In the control group only one snail per replicate did not survive the 28-day experiment, which corresponds to a mortality rate of 4.6%. Also, for most of the native sediments (92%), no or only low (< 10%) mortality occurred. For three samples a significantly increased mortality was observed. These are the sediments from sampling sites 9 (15.9%), 19 (75.0%), and 37 (52.3%). Furthermore, an increased mortality appeared for the sediment sample from sampling site 45 (22.7%), although this was not statistically significant.
3.2.2 Reproduction
The mean embryo number determined for 40 snails at the test start was 12.3 ± 0.567 (mean ± SEM), of which 5.75 ± 0.370 embryos were unshelled (data not shown).
After 28-day exposure, the total number of embryos as well as the number of newly produced embryos without shell was determined for 40 snails per sediment. For six samples (5, 7, 15, 27, 36, 44) one replicate failed, so that only one replicate with 20 snails could be analyzed. For 39 sediments (78%) a significant increase of the embryo number was determined, and for only one sediment a significant decrease of embryo production was noticed (Fig. 2a). The highest stimulation of embryo production was observed for sediment sample 7, where the snails produced about 60% more embryos than the control animals. In comparison to the reproductive output at the test start, the embryo number increased within 28 days by 145% (from 12.3 to 30.1) for sediment sample 7 and by 52.8% (from 12.3 to 18.8) in the control group.

Sediment test with Potamopyrgus antipodarum. Total number of embryos (a) and number of unshelled embryos (b) per female after 28 days of exposure to field sediments. Results are expressed in relative changes compared to the control group (absolute control values, a: 18.8 ± 0.766, b: 7.75 ± 0.488). Significant differences to the control sediment: ★ p < 0.05, ★★ p < 0.01, ★★★ p < 0.001 (Student's t-test); SEM standard error of the mean, † mortality > 20%
Regarding unshelled embryos, an even more distinct stimulation of embryo production occurred (see Fig. 2b). Compared to the control, the exposure to 43 of the sediments (86%) resulted in a significantly increased number of unshelled embryos, while a decreased reproduction was observed for none of the sediments. For sediment sample 39, the highest value with an increase by 80.6% compared to the control was observed (14.0 compared to 7.75).
For the field reference sediment a slight, but not significant, increase of the total embryo number compared to the control was noticed (see Fig. 2a), while the increase of newly produced embryos without a shell was significant (Student's t-test, p < 0.01) (see Fig. 2b).
3.3 Yeast estrogen screen and yeast androgen screen
3.3.1 Yeast estrogen screen
In the YES (Fig. 3a) an estrogenic activity was observed for 33 of the 50 investigated sediment eluates. For two samples (12, 31) eluates were cytotoxic, and thus an estrogen activity could not be determined. Sample 41 exhibited the highest EEQ value with 40.9 ng kg−1. The average value of all 48 sediment extracts (excluding the cytotoxic samples) was 7.77 ng EEQ kg−1. LOQs varied between 0.164 and 3.73 ng EEQ kg−1.

Yeast estrogen screen (a) and yeast androgen screen (b) with sediment eluates. Estrogenicity expressed as EEQs (estradiol equivalents), androgenicity expressed as TEQs (testosterone equivalents). Significant differences to the negative control: ★
p < 0.05, ★★
p < 0.01, ★★★
p < 0.001 (Mann–Whitney U-test); SEM standard error of the mean, NC negative control,  = cytotoxicity, x = below LOQ (limit of quantification)
= cytotoxicity, x = below LOQ (limit of quantification)
3.3.2 Yeast androgen screen
An androgenic activity was observed for 34 (68%) sediments (see Fig. 3b). TEQs were higher than EEQs: the maximum value was 93.4 ng TEQ kg−1 (sample 44) and the average value 32.0 ng TEQ kg−1. Four sediment eluates (samples 3, 5, 12, 29) exerted a toxic effect on the yeast cells, and androgenicity could therefore not be determined. LOQs ranged from 6.23 to 14.4 ng TEQ kg−1.
3.4 Comparison of in vivo and in vitro test results
Comparing the in vitro and in vivo results, 54% of the sediments had a promoting effect on snail reproduction and also showed an estrogenic activity in the YES. 81.8% of the samples which were active in the YES caused an increased snail reproduction compared with the control. Moreover, eluates of 69.2% of the sediments which were active in the Potamopyrgus experiment also activate the estrogen receptor in the YES. Despite these coincidences, the Spearman correlation between EEQs and number of embryos (total and unshelled) was not significant (total: r = −0.097, p = 0.525; unshelled: r = 0.221, p = 0.146). There was no significant correlation between TEQs and the embryo number (total: r = 0.233, p = 0.133; unshelled: r = −0.148, p = 0.344). Here, 56% of the sediments showed an androgenicity in the YAS and also caused an increased reproduction in the Potamopyrgus test.
4 Discussion
4.1 Biotest with P. antipodarum
A higher portion of the field sediments turned out to have a promoting effect on the embryo production in comparison to the artificial control sediment (see Fig. 2). In several studies, P. antipodarum responded with an increased reproduction when exposed to xenoestrogens. The xenoestrogens bisphenol A (BPA) and octylphenol (OP) caused a stimulatory effect on embryo production in laboratory experiments with spiked sediments at environmentally relevant concentrations (Duft et al. 2003a). Also an aqueous exposure to 17α-ethinylestradiol (EE2), BPA, and OP as well as to an estrogenic sewage effluent resulted in an increased reproductive output (Jobling et al. 2004). Schmitt et al. (2008) observed a stimulation of embryo production in snails following an exposure to estrogenic UV screens. Other studies reported a stimulated reproduction after exposure to field sediments and suggested an estrogenic pollution as the likely reason (Oetken et al. 2005; Schmitt et al. 2010). A possible explanation for the increase of embryo numbers for the majority of the sediments in the present study is therefore the presence of estrogenic substances in the field sediments.
Also, for the field reference sediment (site 1) a significantly increased number of unshelled embryos was observed (see Fig. 2b). This was also the case for other study sites far away from known contamination sources such as wastewater treatment plant dischargers and agricultural areas. Estrogenicity of sediments from largely uncontaminated sampling sites can be explained with the presence and impact of phytoestrogens. These compounds can also exhibit biological activity in animals and humans (Murkies et al. 1998). Hence, an influence on the snail reproduction of naturally occurring phytoestrogens is possible.
Another possible explanation for the increased reproduction in the natural reference sediment and other uncontaminated sediments is an impaired reproduction of the snails in the artificial control sediment if it does not provide optimal conditions for the development and reproduction of the snails. However, in 53.1% of the investigated sediments significantly more embryos were produced than in the natural reference sediment (site 1), which itself showed a low but significant estrogenic activity in the YES. This shows that, in comparison to a natural reference sediment, the exposure to the majority of the sediments led to an increased reproductive output, which can be ascribed to an estrogenic contamination of the sediments.
Only a few sediments caused a decreased embryo number. This observation can be explained by either the occurrence of other endocrine active substances—androgens or anti-estrogens—or a general (reproductive) toxic effect of the sediments. Since study site 20, the only one with a significantly decreased embryo number, exhibited a high metal contamination with exceedances of the quality limit values for arsenic, zinc, lead, and cadmium (compare Table 2), a toxic effect is likely. At other study sites similar or even higher contaminations were measured, and the embryo numbers, however, were increased in comparison to the control. This could be explained by the simultaneous occurrence of estrogenic active compounds and an associated superposition of the effects.
In a sediment test with the xenoandrogens tributyltin (TBT) and triphenyltin (TPT), a decline in the number of embryos in P. antipodarum was observed by Duft et al. (2003b). This shows that mudsnails respond to androgen exposure with decreasing embryo number. In the present study, YAS was conducted in order to investigate the androgenic activity of the sediment samples. The comparison of the results of YAS and the Potamopyrgus biotest is discussed in section 4.3.
For whole sediment biotests, an important question is whether the results are influenced by sediment properties such as texture and organic carbon content. In the present study, sediment characteristics exhibited a large variability. However, no influence of different sediment properties on the development of the animals was evident. Correlation analyses between sediment grain size or LOI and the recorded endpoints (mortality, reproduction) showed no statistically significant relationships. The occurrence of the species in a wide range of habitats indicates that P. antipodarum tolerates different sediment characteristics (Alonso and Castro-Díez 2008). Furthermore, the mudsnail shows a wide tolerance to physico-chemical conditions, such as water velocity, temperature, and salinity, and even a high tolerance to desiccation has been reported (Alonso and Castro-Díez 2008). Its robustness and its tolerance to different conditions and circumstances, and concomitant sensitivity to pollutants, make P. antipodarum a suitable test organism for the study of sediments.
4.2 Yeast estrogen screen and yeast androgen screen
4.2.1 Yeast estrogen screen
Sixty-six percent of the sediments showed an estrogenic activity in YES (see Fig. 3a). In order to investigate the most bioavailable substances, aqueous extracts were applied in the assay. An additional extraction with apolar solvents would probably extract further estrogenic active compounds and increase the estrogenicity of the samples. However, it is likely that most of the estrogen active substances have already been removed by the water. Legler et al. (2002) analyzed the estrogenic potency of different solvent fractions of sediment extracts and found that polar fractions showed higher estrogenicity than non-polar ones. Also Houtman et al. (2006) and Viganò et al. (2008) determined highest estrogenic activity in polar fractions of sediment extracts. This suggests that most of the estrogenic compounds present in the investigated sediments were of a hydrophilic nature.
In comparison to sediments from other European regions, the estrogenic activities measured in the present study are rather low. Peck et al. (2004) determined similar YES-EEQ values (21.3–29.9 ng EEQ kg−1) in UK rivers, while values reported in other studies exceed ours by several orders of magnitude. Sediments from three large European river basins (in the Czech Republic, Belgium/the Netherlands, and Spain) exhibited estrogenicities of up to 669 ng EEQ kg−1 (Schmitt et al. 2010), and also sediments collected from UK estuaries showed estrogenic potencies up to 13 μg EEQ kg−1 (Thomas et al. 2004). Grund et al. (2011) determined up to 1.3 μg EEQ kg−1 in sediments from the Danube River in Southern Germany. Also, when the ER-CALUX assay is applied, high values have been determined: 459 ng EEQ kg−1 was measured in sediments from a Dutch harbor (Houtman et al. 2006), up to 340 ng EEQ kg−1 in the Rhine/Meuse estuary in the Netherlands (Houtman et al. 2004), and up to 1.2 μg EEQ kg−1 in other Dutch freshwater sediments (Legler et al. 2003).
It is notable that most other studies assessed sediments of large rivers, whereas this study focusses on small surface waters. This might be an indication that the endocrine pollution of small surface waterbodies is lower than of large ones. However, when comparing EEQ values it must be taken into account that in the other studies sediments were extracted with various solvents, while in the present study aqueous eluates were used. This might also be a reason why the activities in this study are comparatively low.
The vast majority of substances that are responsible for the measured estrogenicity—such as natural and synthetic steroid estrogens, as well as phthalates, PCBs, and alkylphenols—enter the environment via wastewater treatment plants and agricultural runoff. However, for the natural reference sediment a weak estrogenic activity was observed in the YES and also for other study sites away from such contamination sources estrogenicity was measured. One cause may be phytoestrogens. Most studies investigating the estrogenicity of phytohormones report an ERβ selectivity and only a weak affinity for the ERα, which was used in this study (Kuiper et al. 1998; An et al. 2001; Liang et al. 2009). Kohno et al. (2007) studied the estrogenic activity of ethanolic and aqueous extracts from herbal tea mixtures using the same yeast screen as in the present study. The authors could observe a distinct activation of the ERα through the aqueous extracts, more than through the ethanolic extracts, which can be explained with the presence of phytoestrogens extractable with water. In the present study, an activation of the ERα by phytohormones in aqueous sediment eluates is therefore possible.
4.2.2 Yeast androgen screen
The androgenic activity of the sediment eluate samples measured in YAS varied between < LOQ and 93.4 ng TEQ kg−1 (see Fig. 2b). In other studies, partially high androgenicity of environmental samples was also determined. Kirk et al. (2002) investigated the endocrine activity of wastewater during treatment in several sewage plants using the same yeast assay as in the present study. Levels of androgenic activity (up to 4033 ng L−1 DHT (dihydrotestosterone) equivalents in one final effluent) were in general considerably higher than the estrogenic activity values. Thomas et al. (2002) determined high levels of androgenic activity in solvent extracts of sediments (up to 15.3 μg kg−1 DHT equivalents) compared to the androgenicity of surface water (up to 9 ng kg−1 DHT equivalents) and pore water samples (up to 187 ng kg−1 DHT equivalents).
In the past, more interest has been paid to estrogenic than to androgenic activity of environmental samples. This is probably due to the predominance of effects observed in wildlife which are attributed to the exposure to estrogen active compounds. The occurrence of androgenic effects is mostly linked to specific chemicals or effluents. One example is the worldwide observed masculinization of female gastropods (imposex) caused by TBT (Oehlmann et al. 1998; Wirzinger et al. 2007; Galante-Oliveira et al. 2011; Vasconcelos et al. 2011). Furthermore, the masculinization of fish (Howell et al. 1980; Bortone and Cody 1999) or a male biased sex ratio of fish populations (Larsson et al. 2000) exposed to paper/pulp mill effluents have been frequently reported.
Thomas et al. (2002) identified six natural steroids/steroid metabolites in an effluent and estimated that these are responsible for 99% of the in vitro effluent androgenicity in YAS. It can, therefore, be assumed that naturally produced steroids caused the androgenicity of sediment samples in the present study as well. Kirk et al. (2002) suggested that humans are the major source of wastewater androgenic activity. In the raw effluent of wastewater treatment plants in Australia, relatively high concentrations of androsterone and etiocholanolone, which are metabolites of the steroid hormone testosterone, were measured (Tan et al. 2007). Since androgens have greater synthesis and excretion rates as well as higher circulating concentrations in humans than estrogens, concentrations of androgens in wastewater and the aquatic environment may be expected to be much higher than those of estrogens. But since reported endocrine effects in wildlife are predominantly caused by the exposure to estrogens, androgens play a much smaller role. In order to investigate further the possible androgenic effects on wildlife, the development of androgen-specific biomarkers would be helpful.
4.3 Comparison of in vitro and in vivo test results
In our study, in vitro and in vivo tests were combined to analyze the endocrine potential of the sediment samples. Despite the coincidence regarding the estrogenic potential of the sediments in the in vivo and in vitro test, no statistically significant correlation could be found between EEQs/TEQs and snail reproduction. Furthermore, for the only sediment sample with a significantly reduced embryo number (site 20) no androgenicity was measured in the YAS.
In the yeast screens only substances which are able to bind to the human ERα or the human androgen receptor (AR) cause an effect. Although mollusks respond very specifically to exogenous estrogens and androgens (Matthiessen 2008), the role of steroids in the reproduction has not yet been elucidated (Lafont and Mathieu 2007). It seems unlikely that exactly the same substances exert an effect at human steroid receptors and in the Potamopyrgus reproduction test, even though some consistencies exist, which may be one reason for the lack of correlation. Furthermore, the inverted U-shaped concentration response for EDC effects in the mudsnail described in earlier studies has to be considered (Duft et al. 2003a; Jobling et al. 2004; Schmitt et al. 2008). Thus, an increased reproduction rate after (xeno)estrogen exposure can be observed in an intermediate concentration range, but not at low or higher concentrations. Substances occurring in high concentrations in the sediment might show an effect in YES, but not in the reproduction test. Furthermore, the coexistence of androgenic, anti-estrogenic, or (reproductive) toxic compounds might mask estrogenic effects in the Potamopyrgus test.
4.4 Comparison of biotest results and analytical data
For half of the 50 study sites analytical data for PCB concentrations in the suspended particulate matter are available. A conducted Spearman correlation did not show a significant relationship between the concentrations of these chemicals with known endocrine activity and embryo numbers. Nevertheless, it is noticeable that the sediment with the highest PCB contamination (exceeding quality limit values by up to 6.4-fold for four of seven congeners; site 39) caused the highest production of unshelled embryos. Significant correlations between the endpoints evaluated in the Potamopyrgus test (embryo number or mortality) as well as EEQs/TEQs and the contamination with metals, PCBs, PAHs, or OTCs were not found. In other studies, attempts to correlate bioassay effects with chemical data also failed (Vahl et al. 1997; Lyytikäinen et al. 2001; Oetken et al. 2005). Possible causes are the limited number of analyzed chemicals, effects of chemical mixtures (e.g. additive or antagonistic) as well as the differing bioavailability.
It is assumed that alkylphenols as well as natural and synthetic estrogens are important contributors to estrogenicity of environmental samples. However, these analytes were not included in the chemical analytical program provided by HLUG. The total estrogenicity of most sediment extracts of French Rivers studied by Fenet et al. (2003) could be explained mainly by the presence of nonylphenol (NP) and 4 t-octylphenol (OP). Cargouët et al. (2004) determined a contribution of 17α-ethinylestradiol (EE2) to the estimated estrogenic activity in French rivers of 35–50%. 17ß-estradiol (E2) and estrone (E1) were identified as the main contributors to the estrogenic activity of a Dutch harbor sediment (Houtman et al. 2006). In sediment collected near sewage treatment plants in Tokyo Bay, Japan, 34% of the observed estrogenicity of the midpolar fraction, which was the one with the highest estrogenic activity, could be explained by NP and E1 (Hashimoto et al. 2005). Analyses by Viganò et al. (2008) showed that E1, E3 (estriol), and NP were the main estrogenic chemicals in a sediment from an Italian river. However, the authors point out that unidentified substances contribute to the estrogenic potency. Also in other studies, the substances which are responsible for large parts of estrogenicity of environmental samples remain unknown. Only 6% of the estrogenic activity of most sediments investigated in a study by Grund et al. (2011) could be explained by the analyzed compounds. The xenoestrogens NP and BPA as well as E1 were detected in all samples, while E2 and EE2 were below the limit of quantification. In another study, only 3% of wastewater effluent estrogenicity could be explained by the analyzed substances BPA, NP, and OP, while E2, E1, and EE2 could not be determined (Sun et al. 2008). These results indicate that further investigations are necessary in order to identify the compounds the partially high estrogenic potency of environmental samples can be ascribed to. Additionally, chemicals present in sediments in lower concentrations than the detection limit of the chemical analyses, but with high estrogenic potencies (e.g. E2, EE2), potentially contribute to the observed effects (Hollert et al. 2005). In the study by Grund et al. (2011), if it is assumed that the non-detectable estrogens E2 and EE2 were present in sediment extracts in concentrations just below the detection limits, up to 59% of the biological effectiveness could be explained. The development of more sensitive analytical approaches, especially for natural and synthetic hormones, would help to reduce this problem.
For the identification of so-far-unknown chemicals in environmental samples which are responsible for the induction of specific effects, sophisticated techniques such as effect directed analyses (EDA; Brack et al. 2007) or toxicity identification evaluation (TIE; Mehler et al. 2010) are available.
5 Conclusions
This assessment revealed an endocrine activity for the majority of the 50 studied sediments, evident from an increasing reproduction rate in vivo and an estrogenic and androgenic activity in vitro. Seventy-eight percent of the sediment samples caused a significant increased reproductive output in the Potamopyrgus test, 66% showed an estrogenic activity in YES (maximum: 40.9 ng EEQ kg−1 sediment), and 68% showed an androgenic activity in YAS (maximum: 93.4 ng TEQ kg−1). More than half of the sediments had a promoting effect on snail reproduction and also showed an estrogenic activity in YES. Despite these coincidences, no significant correlations could be established between embryo numbers and EEQs. Possible reasons are the detection of different substances due to different modes of action in both test systems and potentially occurring interactions of toxic and/or endocrine compounds. Since P. antipodarum is tolerant to natural stressors and different environmental conditions, but sensitive to pollutants, it seems to be a suitable test organism for the ecotoxicological assessment of field sediments. So far, biological effects monitoring in smaller rivers and creeks is not performed on a routine basis. Also, the assessment of endocrine activities by means of specific bioassays and/or in vivo tests is rather a matter of scientific research than of standardized monitoring programs. The results of the present study show that endocrine disrupting compounds play an important role in aquatic ecosystems and are ubiquitously distributed in surface waters in Hesse, indicating the need to implement effect measures of endocrine activities in monitoring programs.
References
Alonso A, Castro-Díez P (2008) What explains the invading success of the aquatic mud snail Potamopyrgus antipodarum (Hydrobiidae, Mollusca)? Hydrobiologia 614:107–116
An JP, Tzagarakis-Foster C, Scharschmidt TC, Lomri N, Leitman DC (2001) Estrogen receptor β-selective transcriptional activity and recruitment of coregulators by phytoestrogens. J Biol Chem 276:17808–17814
Ankley GT, Benoit DA, Balogh JC, Reynoldson TB, Day KE, Hoke RA (1994) Evaluation of potential confounding factors in sediment toxicity tests with three freshwater benthic invertebrates. Environ Toxicol Chem 13:627–635
Bortone SA, Cody RP (1999) Morphological masculinization in poeciliid females from a paper mill effluent receiving tributary of the St. Johns River, Florida, USA. B Environ Contam Tox 63:150–156
Brack W, Klamer HJC, López de Alda M, Barceló D (2007) Effect-directed analysis of key toxicants in European river basins—a review. Environ Sci Pollut Res 14:30–38
Cargouët M, Perdiz D, Mouatassim-Souali A, Tamisier-Karolak S, Levi Y (2004) Assessment of river contamination by estrogenic compounds in Paris area (France). Sci Total Environ 324:55–66
Chapman PM (1990) The sediment quality triad approach to determining pollution-induced degradation. Sci Total Environ 97(98):815–825
DIN 18123–5 (1996) Bestimmung der Korngrößenverteilung durch Siebung. DIN Deutsches Institut für Normung eV
DIN 38414-S20 (1996) Schlamm und Sedimente. Bestimmung von 6 polychlorierten Biphenylen (PCB). DIN Deutsches Institut für Normung eV
DIN 38414-S3 (1985) Schlamm und Sedimente. Bestimmung des Glührückstandes und des Glühverlustes der Trockenmasse eines Schlammes. DIN Deutsches Institut für Normung eV
DIN EN ISO 17294–2 (2005) Wasserbeschaffenheit—Anwendung der induktiv gekoppelten Plasma-Massenspektrometrie (ICP-MS). Bestimmung von 62 Elementen. DIN Deutsches Institut für Normung eV
Duft M, Schulte-Oehlmann U, Weltje L, Tillmann M, Oehlmann J (2003a) Stimulated embryo production as a parameter of estrogenic exposure via sediments in the freshwater mudsnail Potamopyrgus antipodarum. Aquat Toxicol 64:437–449
Duft M, Schulte-Oehlmann U, Tillmann M, Markert B, Oehlmann J (2003b) Toxicity of triphenyltin and tributyltin to the freshwater mudsnail Potamopyrgus antipodarum in a new sediment biotest. Environ Toxicol Chem 22:145–152
Duft M, Schmitt C, Bachmann J, Brandelik C, Schulte-Oehlmann U, Oehlmann J (2007) Prosobranch snails as test organisms for the assessment of endocrine active chemicals—an overview and a guideline proposal for a reproduction test with the freshwater mudsnail Potamopyrgus antipodarum. Ecotoxicology 16:169–182
EP and EC (2008) Guideline 2008/105/EG of the European Parliament and the Council, 16-Dec-2008, concerning quality limit values in the field of water policy and for amending and subsequently repealing of the Council Directives 82/176/EWG, 83/513/EWG, 84/156/EWG, 84/491/EWG and 86/280/EWG as well as for amending the Directive 2000/60/EG
Fenet H, Gomez E, Pillon A, Rosain D, Nicolas J-C, Casellas C, Balaguer P (2003) Estrogenic activity in water and sediments of a French river: contribution of alkylphenols. Arch Environ Cont Toxicol 44:1–6
Galante-Oliveira S, Oliveira I, Ferreira N, Santos JA, Pacheco M, Barroso C (2011) Nucella lapillus L. imposex levels after legislation prohibiting TBT antifoulants: temporal trends from 2003 to 2008 along the Portuguese coast. J Environ Monit 13:304–312
Giudice BD, Young TM (2010) The antimicrobial triclocarban stimulates embryo production in the freshwater mudsnail Potamopyrgus antipodarum. Environ Toxicol Chem 29:966–970
Grund S, Higley E, Schönenberger R, Suter MJF, Giesy JP, Braunbeck T, Hecker M, Hollert H (2011) The endocrine disrupting potential of sediments from the Upper Danube River (Germany) as revealed by in vitro bioassays and chemical analysis. Environ Sci Pollut Res 18:446–460
Guillette LJ, Gross TS, Masson GR, Matter JM, Percival HF, Woodward AR (1994) Developmental abnormalities of the gonad and abnormal sex hormone concentrations in juvenile alligators from contaminated and control lakes in Florida. Environ Health Pers 102:680–688
Hashimoto S, Horiuchi A, Yoshimoto T, Nakao M, Omura H, Kato Y, Tanaka H, Kannan K, Giesy JP (2005) Horizontal and vertical distribution of estrogenic activities in sediments and waters from Tokyo Bay, Japan. Arch Environ Cont Toxicol 48:209–216
HMUELV (2009) Hessisches Ministerium für Umwelt, Energie, Landwirtschaft und Verbraucherschutz. Kartenservice zur Umsetzung der Wasserrahmenrichtlinie in Hessen. Available via http://wrrl.hessen.de/viewer.htm. Accessed 15 Jan 2011
Hollert H, Dürr M, Holtey-Weber R, Islinger M, Brack W, Färber H, Erdinger L, Braunbeck T (2005) Endocrine disruption of water and sediment extracts in a non-radioactive dot blot/RNAse protection-assay using isolated hepatocytes of rainbow trout—deficiencies between bioanalytical effectiveness and chemically determined concentrations and how to explain them. Environ Sci Pollut Res 12:347–360
Houtman CJ, Cenijn PH, Hamers T, Lamoree MH, Legler J, Murk AJ, Brouwer A (2004) Toxicological profiling of sediments using in vitro bioassays, with emphasis on endocrine disruption. Environ Toxicol Chem 23:32–40
Houtman CJ, Booij P, Jover E, del Rio DP, Swart K, van Velzen M, Vreuls R, Legler J, Brouwer A, Lamoree MH (2006) Estrogenic and dioxin-like compounds in sediment from Zierikzee harbour identified with CALUX assay-directed fractionation combined with one and two dimensional gas chromatography analyses. Chemosphere 65:2244–2252
Howell WM, Black DA, Bortone SA (1980) Abnormal expression of secondary sex characters in a population of mosquitofish, Gambusia affinis holbrooki—evidence for environmentally induced masculinization. Copeia 4:676–681
Jobling S, Casey D, Rogers-Gray T, Oehlmann J, Schulte-Oehlmann U, Pawlowski S, Baunbeck T, Turner AP, Tyler CR (2004) Comparative responses of molluscs and fish to environmental estrogens and an estrogenic effluent. Aquat Toxicol 66:207–222
Ketata I, Denier X, Hamza-Chaffai A, Minier C (2008) Endocrine-related reproductive effects in molluscs. Comp Biochem Phys C 147:261–270
Kirk LA, Tyler CR, Lye CM, Sumpter JP (2002) Changes in estrogenic and androgenic activities at different stages of treatment in wastewater treatment works. Environ Toxicol Chem 21:972–979
Kohno H, Kouda K, Tokunaga R, Sonoda Y (2007) Detection of estrogenic activity in herbal teas by in vitro reporter assays. Eur Food Res Technol 225:913–920
Kuiper GGJM, Lemmen JG, Carlsson B, Corton JC, Safe SH, van der Saag PT, van der Burg P, Gustafsson JA (1998) Interaction of estrogenic chemicals and phytoestrogens with estrogen receptor β. Endocrinology 139:4252–4263
Lafont R, Mathieu M (2007) Steroids in aquatic invertebrates. Ecotoxicology 16:109–130
Larsson DGJ, Hällman H, Förlin L (2000) More male fish embryos near a pulp mill. Environ Toxicol Chem 19:2911–2917
LAWA (1998) Länderarbeitsgemeinschaft Wasser (Regional Working Group on Water): quality limit values for the protection of inland surface waters, volume II: Derivation and testing of quality limit values for the protection of inland surface waters for the heavy metals lead, cadmium, chromium, copper, nickel, mercury, and zinc. Germany, Berlin
LeBlanc GA (2007) Crustacean endocrine toxicology: a review. Ecotoxicology 16:61–81
Legler J, Dennekamp M, Vethaak AD, Brouwer A, Koeman JH, van der Burg B, Murk AJ (2002) Detection of estrogenic activity in sediment-associated compounds using in vitro reporter gene assays. Sci Total Environ 293:69–83
Legler J, Leonards P, Spenkelink A, Murk AJ (2003) In vitro biomonitoring in polar extracts of solid phase matrices reveals the presence of unknown compounds with estrogenic activity. Ecotoxicology 12:239–249
Liang K, Yang L, Xiao Z, Huang J (2009) A bipartite recombinant yeast system for the identification of subtype-selective estrogen receptor ligands. Mol Biotechnol 41:53–62
Lyytikäinen M, Sormunen A, Ristola T, Juvonen R, Kukkonen JVK (2001) Toxicity of freshwater sediments in the vicinity of an old sawmill: application of three bioassays. Arch Environ Cont Toxicol 40:318–326
Matthiessen P (2008) An assessment of endocrine disruption in mollusks and the potential for developing internationally standardized mollusk life cycle test guidelines. Integr Environ Assess Manag 4:274–284
Mehler WT, Maul JD, You J, Lydy MJ (2010) Identifying the causes of sediment-associated contamination in the Illinois River (USA) using a whole-sediment toxicity identification evaluation. Environ Toxicol Chem 29:158–167
Murkies AL, Wilcox G, Davis SR (1998) Clinical review 92—phytoestrogens. J Clin Endocr Metab 83:297–303
Oehlmann J, Bauer B, Minchin D, Schulte-Oehlmann U, Fioroni P, Markert B (1998) Imposex in Nucella lapillus and intersex in Littorina littorea: interspecific comparison of two TBT-induced effects and their geographical uniformity. Hydrobiologia 378:199–213
Oehlmann J, Schulte-Oehlmann U, Tillmann M, Markert B (2000) Effects of endocrine disruptors on prosobranch snails (Mollusca: Gastropoda) in the laboratory. Part I: Bisphenol A and octylphenol as xeno-estrogens. Ecotoxicology 9:383–397
Oehlmann J, Di Benedetto P, Tillmann M, Duft M, Oetken M, Schulte-Oehlmann U (2007) Endocrine disruption in prosobranch molluscs: evidence and ecological relevance. Ecotoxicology 16:29–43
Oetken M, Stachel B, Pfenninger M, Oehlmann J (2005) Impact of a flood disaster on sediment toxicity in a major river system—the Elbe flood 2002 as a case study. Environ Pollut 134:87–95
Peck M, Gibson RW, Kortenkamp A, Hill EM (2004) Sediments are major sinks of steroidal estrogens in two United Kingdom rivers. Environ Toxicol Chem 23:945–952
Routledge EJ, Sumpter JP (1996) Estrogenic activity of surfactants and some of their degradation products assessed using a recombinant yeast screen. Environ Toxicol Chem 15:241–248
Rubinos D, Iglesias L, Devesa-Rey R, Díaz-Fierros F, Barral MT (2010) Arsenic release from river sediments in a gold-mining area (Anllóns River basin, Spain): effect of time, pH and phosphorus concentration. Eur J Mineral 22:665–678
Rutishauser BV, Pesonen M, Escher BI, Ackermann GE, Aerni HR, Suter MJF, Eggen RIL (2004) Comparative analysis of estrogenic activity in sewage treatment plant effluents involving three in vitro assays and chemical analysis of steroids. Environ Toxicol Chem 23:857–864
Santos MM, Castro LFC, Vieira MN, Micael J, Morabito R, Massanisso P, Reis-Henriques MA (2005) New insights into the mechanism of imposex induction in the dogwhelk Nucella lapillus. Comp Biochem Phys C 141:101–109
Schmitt C, Oetken M, Dittberner O, Wagner M, Oehlmann J (2008) Endocrine modulation and toxic effects of two commonly used UV screens on the aquatic invertebrates Potamopyrgus antipodarum and Lumbriculus variegatus. Environ Pollut 152:322–329
Schmitt C, Balaam J, Leonards P, Brix R, Streck G, Tuikka A, Bervoets L, Brack W, van Hattum B, Meire P, de Deckere E (2010) Characterizing field sediments from three European river basins with special emphasis on endocrine effects—a recommendation for Potamopyrgus antipodarum as test organism. Chemosphere 80:13–19
Sohoni P, Sumpter JP (1998) Several environmental oestrogens are also anti-androgens. J Endocrinol 158:327–339
Sumpter JP (2005) Endocrine disrupters in the aquatic environment: an overview. Acta Hydroch Hydrob 33:9–16
Sun Q, Deng S, Huang J, Shen G, Yu G (2008) Contributors to estrogenic activity in wastewater from a large wastewater treatment plant in Beijing, China. Eviron Toxicol Phar 25(1):20–26
Tan BLL, Hawker DW, Müller JF, Leusch FDL, Tremblay LA, Chapman HF (2007) Comprehensive study of endocrine disrupting compounds using grab and passive sampling at selected wastewater treatment plants in South East Queensland, Australia. Environ Int 33:654–669
Thomas KV, Hurst MR, Matthiessen P, McHugh M, Smith A, Waldock MJ (2002) An assessment of in vitro androgenic activity and the identification of environmental androgens in United Kingdom estuaries. Environ Toxicol Chem 21:1456–1461
Thomas KV, Balaam J, Hurst M, Nedyalkova Z, Mekenyan O (2004) Potency and characterization of estrogen-receptor agonists in United Kingdom estuarine sediments. Environ Toxicol Chem 23:471–479
Vahl HH, Karbe L, Westendorf J (1997) Genotoxicity assessment of suspended particulate matter in the Elbe river: comparison of Salmonella microsome test, arabinose resistance test, and umu-test. Mutat Res-Gen Tox En 394:81–93
Vasconcelos P, Moura P, Barroso CM, Gaspar MB (2011) Size matters: importance of penis length variation on reproduction studies and imposex monitoring in Bolinus brandaris (Gastropoda: Muricidae). Hydrobiologia 661:363–375
Viganò L, Benfenati E, van Cauwenberge A, Eidem JK, Erratico C, Goksøyr A, Kloas W, Maggioni S, Mandich A, Urbatzka R (2008) Estrogenicity profile and estrogenic compounds determined in river sediments by chemical analysis, ELISA and yeast assays. Chemosphere 73:1078–1089
VO-WRRL (2010) Verordnung zur Umsetzung der Wasserrahmenrichtlinie—Hessen (Regulation for the implementation of the Water Framework Directive—Hesse), 17-May-2005, amended by the regulation from the 12-July-2010 (Law and Ordinance Gazette I p. 225), Annex 4 (to section 6 (1), sentence 2 and section 6 (2))
Wagner M, Oehlmann J (2009) Endocrine disruptors in bottled mineral water: total estrogenic burden and migration from plastic bottles. Environ Sci Pollut Res 16:278–286
Wirzinger G, Vogt C, Bachmann J, Hasenbank M, Liers C, Stark C, Ziebart S, Oehlmann J (2007) Imposex of the netted whelk Nassarius reticulatus (Prosobranchia) in Brittany along a transect from a point source. Cah Biol Mar 48:85–94
Acknowledgements
The authors would like to thank the Hessian Agency for the Environment and Geology (HLUG) for funding. HLUG staff, in particular Dr. Peter Seel and Silvia Fengler, are acknowledged for the cooperation and for providing chemical analytical data. Furthermore, we thank Martin Wagner for providing knowledge and support in realizing the yeast screens and Dr. Matthias Oetken for reviewing the manuscript.
Author information
Authors and Affiliations
Corresponding author
Additional information
Responsible editor: Henner Hollert
An erratum to this article can be found at http://dx.doi.org/10.1007/s11368-011-0465-x.
Electronic supplementary material
Below is the link to the electronic supplementary material.
ESM 1
Overview of the sampling sites (DOC 153 kb)
Rights and permissions
About this article
Cite this article
Galluba, S., Oehlmann, J. Widespread endocrine activity in river sediments in Hesse, Germany, assessed by a combination of in vitro and in vivo bioassays. J Soils Sediments 12, 252–264 (2012). https://doi.org/10.1007/s11368-011-0454-0
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s11368-011-0454-0