
Abstract
Purpose
The relationship between plant absorption and accumulation of heavy metals and the effect of iron plaque on roots of wetland plants are unknown, especially for plants grown in heavy metal-contaminated soil. This experiment was designed to study the effects of iron addition on the formation of iron plaque in the rhizosphere of the wetland plant species Iris pseudacorus L. in artificial Pb-contaminated soil and the effects of iron plaque on Pb accumulation by plants.
Materials and methods
Soil was collected from 0- to 30-cm depth of a clayey illitic thermic typic epiaqualfs in an abandoned paddy field in Jiaxing of Zhejiang Province, China. The seeds of yellow flag (I. pseudacorus L.) were germinated in soil mixed with vermiculite and grown for 30 days. Three treatments of iron (0, 100, and 500 mg Fe kg−1 as FeSO4) were added when uniform seedlings were transplanted into plastic pots filled with 1.0-kg soil, which had been previously treated with four treatments of Pb [0, 100, 500, and 1,000 mg Pb kg−1 as Pb(NO3)2] and waterlogged for 72 days. The yellow flags were grown in a greenhouse for 60 days. The plaque on roots was extracted using the cold DCB (dithionite–citrate–bicarbonate) technique. The plant samples were digested with 10 mL concentrated nitric acid by microwave digestion. The concentration of Fe and Pb were determined using atomic spectrophotometry (NovAA300, Germany).
Results and discussion
The amount of iron plaque on the roots of yellow flag (I. pseudacorus L.) was markedly affected by Fe addition but not by Pb addition. The total Pb accumulated in plants was increased initially and then decreased with the addition of Fe in three lower Pb treatments (0, 100, and 500 mg Pb kg−1) but decreased with more Fe addition in the 1,000 mg Pb kg−1 treatment. The pH value was decreased with the addition of more iron, which increases Pb availability, and heavy doses of iron caused iron toxicity. Plant growth decreased due to iron toxicity at higher iron additions (500 mg kg−1), while Pb accumulation decreased. Intermediate levels of iron supply (100 mg kg−1) enhanced Pb absorption in roots, which improved phytoremediation effectiveness.
Conclusions
Iron supply enhanced the amount of iron plaque and increased both Pb adsorbed on the roots and Pb uptake by plants. However, plant growth was inhibited by iron toxicity at high iron dose (500 mg kg−1) and biomass decreased. The intermediate iron dose (100 mg kg−1) generally enhanced Pb absorption and accumulation, which enhances the ability of yellow flag to remove Pb from Pb-contaminated soils.
Similar content being viewed by others
Explore related subjects
Discover the latest articles, news and stories from top researchers in related subjects.Avoid common mistakes on your manuscript.
1 Introduction
A number of air, water, and soil processes and pathways are involved in the emission of heavy metals into the environment (Järup 2003). Major sources of lead (Pb) in the environment are oil or wastes, plants used as solid fuel, metal smelting, and burning of coal (Mihai et al. 2007). Pb toxicity inhibits root elongation (Chao et al. 2007). Peroxidase activity and the malondialdehyde induced at high Pb concentration provoke ultrastructural damage to the leaf (Hu et al. 2007). Heavy metal contamination is a serious threat for the safety of aquatic ecosystems, agro-ecosystem, and human health (Järup 2003; Farmer et al. 2006; Overesch et al. 2007). Constructed wetlands are widely used for removal of heavy metals in treating wastewater (Manios et al. 2003; Deng et al. 2006; Yang et al. 2006). Sedimentation, sorption, co-precipitation, cation exchange, phytoaccumulation, and microbial activity are the main processes for the removal of heavy metals (Sheoran and Sheoran 2006). Wetland plants develop aerenchyma cells to transfer O2 from the aerial parts to the roots, inducing the precipitation of iron oxides or hydroxides on the root surfaces (Chen et al. 1980; Armstrong et al. 1994).
Iron oxide coating (iron plaque) has been found on the roots of many wetland plants (Otte et al. 1989; Ye et al. 1997; Sundby et al. 1998; Hansel et al. 2001). The presence and degree of iron plaque formation are controlled by both abiotic and biotic factors (Mendelssohn et al. 1995). Several possible mechanisms for oxidation of Fe (II) have been proposed with O2 diffused from wetland plant roots considered the best mechanism (Sundby et al. 1998). In this mechanism, iron-oxidizing bacteria play a role in Fe plaque formation (Emerson et al. 1999; Weiss et al. 2005). Producing oxidase in the rhizosphere may participate in the oxidation of Fe (II) to form iron plaque (Mitsui et al. 1962). The iron plaque occurs primarily as fillings or casts in exposed cavities of epidermal cells (Chen et al. 1980). Iron (hydr)oxide precipitates (or plaques) occurring on the surface of roots may consist of ferrihydrite, goethite, and siderite (St-Cyr and Crowder 1989; Hansel et al. 2001). Due to the high capacity of functional groups on iron (hydr)oxides to sequester metals by adsorption and/or co-precipitation, researchers have paid considerable attention to iron plaque (Liu et al. 2004; Chen et al. 2006). Recently, the influence of iron plaque on the growth of wetland plant roots and on heavy metal absorption and accumulation by plants has been studied in laboratory and field experiments (Otte et al. 1989; Ye et al. 1997, 1998). However, the relationship between the amount of heavy metal absorption and accumulation and iron plaque on roots of wetland plants is still unclear.
Iris pseudacorus L. is an erect glabrous perennial wetland plant (Kim 2008) with an exceptionally high ability to absorb heavy metals even under unfavorable conditions. As a result, it has been used as an efficient economical approach for removing heavy metals from wastewater (Piccardi and Clauser 1983; Barbolani et al. 1986; Zhang et al. 2007). In this study, we hypothesized that iron addition could benefit the formation of iron plaque on the roots of plants, and the amount of iron plaque would significantly affect Pb absorption and accumulation on plants growing in Pb-contaminated soil. The objective of the present study was to determine the optimum amount of iron addition for improving Pb accumulation in the wetland plant species I. pseudacorus L.
2 Materials and methods
2.1 Soil
The test soil was collected from 0- to 30-cm depth in an abandoned paddy field in Jiaxing of Zhejiang province, China. The soil was classified as a clayey illitic thermic typic epiaqualfs and was previously under long-term agricultural use. The soil was sieved through a 2-mm sieve after being air-dried. It had a pH of 5.61 (soil:water = 1:2.5), organic matter content of 3.62%, and total phosphorus, nitrogen, iron, and lead contents of 1.03 g kg−1, 2.32 g kg−1, 2.95%, and 45.58 mg kg−1, respectively.
2.2 Greenhouse experiment
The seeds of yellow flag (I. pseudacorus L.) collected at a pond site in Huajiachi campus of Zhejiang University, Hangzhou were germinated in soil mixed with vermiculite and grown for 30 days. Three treatments of iron (0, 100, and 500 mg Fe kg−1 as FeSO4) were added when uniform seedlings were transplanted into plastic pots filled with 1.0 kg soil. The soil had been previously treated with four treatments of Pb [0, 100, 500, and 1,000 mg Pb kg−1 as Pb(NO3)2] and waterlogged for 72 days. Each treatment was replicated three times. The pots were arranged randomly, and their position was rotated regularly to ensure uniform conditions in the greenhouse. Water was kept about 0.5 cm above the soil surface for 60 days.
2.3 Analytical methods
At harvest, plant samples were separated into shoots and roots. Fresh roots were washed with tap water, then rinsed in deionized water, and dried on filter paper. The plaque on fresh roots was then extracted using the cold DCB (dithionite–citrate–bicarbonate) technique (Taylor and Crowder 1983). The plant samples including roots and shoots were dried at 105 °C for 15 min, and then dried at 70 °C for 24 h and dry weights recorded. Ground samples (about 0.2–0.5 g weight) were digested in 10 mL concentrated nitric acid by microwave digestion. A three-step digestion procedure was used: initial rise to 80 °C for 2 min, then to 120 °C for 5 min, and a final digestion at 185 °C for 20 min. The digests were transferred to 100-mL volumetric flasks with deionized water and filtered into plastic bottles. The concentrations of Fe and Pb were determined by atomic spectrophotometry (NovAA300, Germany).
2.4 Calculation and statistical analysis
Mean values were compared using analysis of variance with post hoc multiple comparisons using the least significant difference (LSD) test at the 5% level. Correlation analysis was carried out by bivariate correlation (two-tailed) and test of significance in SPSS 11.5. Some data processing was carried out using Micro Excel 2003.
3 Results
3.1 Plant biomass as affected by Pb2+ and Fe2+ treatments
The dry weight of I. pseudacorus L. roots was dramatically affected by both Pb addition (F = 14.3, p < 0.001) and Fe supply (F = 70.6, p < 0.01), and the interaction of Pb and Fe was significant (F = 8.4, p < 0.01) (Table 1). The dry weight of roots increased with increasing Pb dose in the absence of iron addition. However, it decreased in the presence of 100 mg Fe kg−1 (see Table 1). The dry weight of roots treated with 500 mg Fe kg−1 was the lowest among the three Fe treatments (0, 100, and 500 mg Fe kg−1). Fe (F = 20.6, p < 0.01) had a significant influence on the dry weight of shoots as compare to Pb (F = 1.9, p > 0.05). The dry weight of shoots treated with 500 mg Fe kg−1 was the lowest among the three iron additions in four Pb treatments (see Table 1).
3.2 Iron plaque formation and lead adsorption on the roots
The iron plaque covering the roots of yellow flag was very thick and could be mechanically separated from the roots. The roots covered with heavy iron plaque became very brittle. The amount of iron plaque on the roots was markedly affected by Fe addition (F = 7.0, p < 0.05) but not by Pb additions. Iron plaque formation (F = 3.0, p < 0.05) was also affected by an interaction between Pb and Fe. Thus, the addition of Fe was found to strongly affect the amount of plaque formation (Table 2).
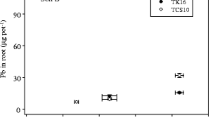
The concentration of Pb in DCB extracts was significantly affected by the addition of Pb (F = 28.0, p < 0.01) and Fe (F = 13.9, p < 0.01), and the interaction between Fe and Pb also had a significant influence on the content of Pb in DCB extracts (F = 3.9, p < 0.01). Similarly, the addition of Fe increased the content of Pb adsorbed by iron plaque (Fig. 1). The pattern of the content of Pb on roots with Pb addition in the 0 and 100 mg Fe kg−1 additions initially increased and then decreased. However, the content of Pb increased again in the 500 mg Fe kg−1 addition. The correlation coefficient of Pb content adsorbed by iron plaque and amount of iron plaque on the roots was 0.61 (n = 36, p < 0.01).

Pb adsorbed by iron plaque on the roots in different treatments of Pb and Fe2+
3.3 Lead in roots and shoots
The content of Pb in the roots after DCB extraction was increased with increasing Pb supply in the three Fe treatments, and Pb content with Fe additions was generally higher than the control (no Fe addition) (Fig. 2a). Moreover, the difference of Pb content between treatments with high Fe (500 mg Fe kg−1) and low Fe addition (0 and 100 mg Fe kg−1) was significant in the higher Pb additions (500 and 1,000 mg Pb kg−1). The Pb content was significantly affected by both Pb (F = 585.6, p < 0.01) and Fe (F = 224.1, p = 0.01) addition, and the interaction of Pb and Fe also had a significant influence on Pb content in the roots after DCB extraction (F = 78.1, p < 0.01). The content of Pb in shoots was increased with increasing Pb addition in the three Fe treatments (F = 77.2, p < 0.01) (Fig. 2b). However, Pb content in shoots was not affected by Fe addition or a Pb and Fe interaction. Pb accumulated in roots was from 50.4% to 78.3% of total Pb accumulated by plants (Fig. 3). The total Pb accumulated in plants was initially increased then decreased with increasing Fe addition for the three Pb additions (0, 100, and 500 mg Pb kg−1), and Pb accumulation decreased with Fe addition in the 1,000 mg Pb kg−1 treatment.

The concentration of Pb in roots after DCB extraction (a) and in shoots (b)

Accumulation of Pb in roots and shoots in different treatments
4 Discussion
Lead is not an essential element for plant growth and is considered as a toxic element for plants when it occurs at high concentration. Lead has been shown to inhibit photosynthesis and transpiration and decrease the content of chlorophyll (Parys et al. 1998). In the present study, plant growth was significantly affected by both Fe and Pb additions (see Table 1). When iron addition was 500 mg kg−1, the dry weight of root and shoots was generally far lower than that of lower iron levels for a given Pb treatment. These results suggest that iron toxicity induced by the higher iron addition had a negative influence on yellow flag growth. In order to prevent excessive Fe uptake, wetland plants have evolved a mechanism to oxidize iron from its ferrous to ferric form resulting in iron plaque formation in the rhizosphere. However, higher Fe (II) concentration in waterlogged soil could induce iron toxicity and inhibit plant growth (Batty and Younger 2003). It was found that the pH of the surface water decreased to 3.95 from 4.22 in the 500 mg Fe kg−1 treatment compared to the range of 5.50 to 7.00 for the lower Fe treatments (data not shown). With increasing Fe addition, more Pb was adsorbed on roots for a given Pb addition level (see Fig. 1). The reason may be due to the small biomass of roots at higher Fe levels. However, Wang and Zhou (2003) reported that the decrease of pH should enhance Pb availability, which could occur in our study because the higher iron resulted in lower pH values. Both excessive Fe and Pb had a great influence on root metabolism, and the higher Pb concentration inhibited photosynthesis (Parys et al. 1998), which resulted in decreased plant growth.
Heavy iron plaque was observed on the roots of yellow flag, and more precipitation generally occurred on the roots with higher iron additions (see Table 2). However, there was no significant positive correlation between the amount of iron plaque and Fe addition rates. The presence and degree of iron plaque formation are controlled by both abiotic and biotic factors, the most important abiotic factor is Fe availability in soil (Mendelssohn et al. 1995), and the biotic factor is generally believed to be the degree of radial oxygen loss (ROL) from the roots (Jespersen et al. 1998). Several possible mechanisms for Fe (II) oxidation have been proposed, and O2 diffusion from wetland plant roots is considered an important mechanism (Sundby et al. 1998). The mechanism of forming rhizoconcretions of Fe (hydr)oxide occurs as the redox boundary moves radially outward from the root surface such that precipitation takes place progressively farther away from the root surface (Sundby et al. 1998). However, Armstrong (1967) calculated that radial O2 loss could account for only 1/9 of the total oxidizing capacity of rice roots. The ROL of yellow flag is low, and the highest oxygen release rate observed for I. pseudacorus was 0.34 mg h−1 plant−1 in the Eh range of –250 to –150 mV (Wieβner et al. 2002). Some researchers found that other sources of oxygen might contribute to rhizosphere oxidation of iron, for example glycolic acid oxidase (Mitsui et al. 1962), and excessive Fe2+ uptake induced production of superoxide radial, which leads to H2O2 formation. The formation of hydroxyl radical is the result of H2O2 accumulation, which reacts with iron by the Fenton reaction and causes lipid peroxidation, inactivation of membrane proteins, loss of cellular integrity, and ultimately death (Hendry and Brocklebank 1985). Root enzymatic oxidation and bacteria in the rhizosphere may play an important role in contributing to Fe (II) oxidation in the rhizosphere (Mendelssohn et al. 1995). The non-protein amino acid 3-(3-carboxyphenyl) alanine is excreted in considerable amounts by the roots and is microbially degraded at a slower rate than protein amino acids (Cardenas and Kichuth 1978). Thus, root enzymatic oxidation in the rhizosphere might be one of the causes for heavy formation of iron plaque on the roots of I. pseudacorus L. in this study.
Pb in iron plaque was positively correlated with the amount of iron plaque (r = 0.61, p < 0.01) (Fig. 4), this implied that Pb was adsorbed or co-precipitated with iron (hydr)oxide plaque on the roots; resulting in enhanced accumulation of Pb on the roots. In the present study, the pH in the soil was below 7.0, and the reported point of zero charge (PZC) of most Fe (hydr)oxides usually range from 7.0 to 9.5 (Violante et al. 2003). However, Fe (hydr)oxides selectively adsorb divalent cations even at solution pH values lower than the PZC of metal oxides. Metal ions associated with hydrous (hydr)oxide surfaces raise the PZC value of (hydr)oxides (Violante et al. 2003), and the following metal affinity series for freshly precipitated Fe has been reported: Pb > Cu > Zn > Ni > Cd > Co > Sr > Mg (Jackson 1998). There was a significant positive correlation between Pb in root and Pb on the root (r = 0.95; p < 0.01) (Fig. 5), and the correlation coefficient between Pb in shoots and Pb in roots was 0.79 (p < 0.01) (Fig. 6). Based on the correlation analysis of Pb in different parts of the plant, formation of iron plaque on the roots of yellow flag improved Pb uptake. Similar results were obtained by Liu et al. (2007), who found that iron plaque enhanced Pb absorption in rice (Oryza sativa L.), and Ye et al. (1998), who found that iron plaque was not a barrier to Pb uptake and translocation in Typha latifolia. However, some researchers observed that the presence of iron plaque reduced the uptake of other metals by the plants. Zinc uptake of Aster tripolium is reduced when plaque was presented (Otte et al. 1989), and manganese uptake by Phragmites australis was decreased (Batty et al. 2000). The discrepancies in effects of plaque on metal uptake in different plants and experimental conditions indicate that further broad studies are needed for various plants and metals.

Relationship between the concentration of Pb (g kg−1 root dry weight) and Fe (g kg−1 root dry weight) in iron plaque of yellow flag

Relationship between the concentration of Pb (mg kg−1 root dry weight ± standard deviation) in roots after DCB extraction and Pb in iron plaque (g kg−1 root dry weight ± standard deviation) of yellow flag

Relationship between the concentration of Pb in shoots (mg kg−1 root dry weight ± standard deviation) and Pb in the roots of yellow flag after DCB extraction
The total accumulation of Pb in the roots was 1.01 to 3.6 times that in shoots (see Fig. 3), which indicates that Pb mainly accumulated in roots. Fitzgerald et al. (2003) found that Pb accumulated mainly in the roots of monocots but in the shoots of dicots, and T. latifolia was found to accumulate Pb mostly in the roots (Ye et al. 1997). Compared with the no iron treatment, Pb accumulation in shoots of the 100 mg Fe kg−1 treatment was slightly enhanced; however, treatments with 500 mg Fe kg−1 decreased Pb accumulation. Although more iron addition resulted in more iron plaque formation and more Pb adsorbed, iron toxicity induced by higher iron addition decreased plant growth (see Table 1) and Pb accumulation also decreased (see Fig. 3). Thus, intermediate iron additions (100 mg kg−1) could enhance Pb absorption in roots resulting in improved phytoremediation. If the roots are never removed from the soil, Pb would return to the soil after plant death and decay. Weis and Weis (2004) pointed out that studies were needed regarding the turnover of nutritive roots and the potential release of metals from these decomposing roots.
5 Conclusions
Our results indicated that iron addition can enhance the amount of iron plaque on the roots of yellow flag planted in artificially Pb-contaminated soils and improved Pb absorption by plants. However, iron toxicity at higher iron addition (500 mg kg−1) inhibited plant growth and resulted in a decrease of biomass. Thus, intermediate rates of iron addition (100 mg kg−1) generally enhanced Pb accumulation and Pb removal by yellow flag.
References
Armstrong W (1967) The oxidizing activity of roots in waterlogged soils. Physiol Plant 20:920–926
Armstrong W, Strange ME, Cringle S, Beckett PM (1994) Microelectrode and modeling study of oxygen distribution in roots. Ann Bot 74:287–299
Barbolani E, Clauser M, Pantani F, Gellini R (1986) Residual heavy metal (Cu and Cd) removal by Iris pseudacorus. Water Air Soil Pollut 28:277–282
Batty LC, Younger PL (2003) Effects of external iron concentration upon seedling growth and uptake of Fe and phosphate by the common reed, Phragmites australis (Cav.) Trin ex. Steudel. Ann Bot 92:801–805
Batty LC, Baker AJM, Wheeler BD, Curtis CD (2000) The effect of pH and plaque on the uptake of Cu and Mn in Phragmites australis (Cav.) Trin ex. Steudel. Ann Bot 86:647–653
Cardenas ML, Kickuth R (1978) Secondary metabolites from macrophytes and their ecochemical function. Part 3. 3-3 carboxyphenyl alanine as constituent and excretion product of Iris pseudacorus. Angew Bot 52:203–214
Chao L, Zhou QX, Chen S (2007) Single and joint stress of acetochlor and Pb on three agricultural crops in northeast China. J Environ Sci-China 19:719–724
Chen CC, Dixon JB, Turner FT (1980) Iron coatings on rice roots: morphology and models of development. Soil Sci Soc Am J 44:1113–1119
Chen SB, Zhu YG, Ma YB (2006) Effects of phosphate amendments on Pb extractability and movement of phosphorus in contaminated soil (in Chinese). Acta Scientiae Circumstantiae 26:1140–1144
Deng H, Ye ZH, Wong MH (2006) Lead and zinc accumulation and tolerance in populations of six wetland plants. Environ Pollut 141:69–80
Emerson D, Weiss JV, Megonigal JP (1999) Iron-oxidizing bacteria are associated with ferric hydroxide precipitates (Fe-plaque) on the roots of wetland plants. Appl Environ Microbiol 65:2758–2761
Farmer JG, MacKenzie AB, Moody GH (2006) Human teeth as historical biomonitors of environmental and dietary lead: some lessons from isotopic studies of 19th and 20th century archival material. Environ Geochem Health 28:421–430
Fitzgerald EJ, Caffrey JM, Nesaratnam ST, McLoughlin P (2003) Copper and lead concentrations in salt marsh plants on the Suir Estuary, Ireland. Environ Pollut 123:67–74
Hansel CM, Fendorf S, Sutton S, Newville M (2001) Characterization of Fe plaque and associated metals on the roots of mine-waste impacted aquatic plants. Environ Sci Technol 35:3863–3868
Hendry GAF, Brocklebank JD (1985) Iron-induced oxygen radical damage in waterlogged plants. New Phytol 101:199–206
Hu JZ, Shi GX, Xu QS, Wang X, Yuan QH, Du KH (2007) Effects of Pb2+ on the active oxygen-scavenging enzyme activities and ultrastructure in Potamogeton crispus leaves. Russ J Plant Physl 54:414–419
Jackson TA (1998) The biogeochemical and ecological significance of interactions between colloidal minerals and trace elements. In: Parker A, Rae JE (eds) Environmental interactions of clays. Springer-Verlag, Berlin, pp 9–205
Järup L (2003) Hazards of heavy metal contamination. Brit Med Bull 68:167–182
Jespersen DN, Sorrell BK, Brix H (1998) Growth and root oxygen release by Typha latifolia and its effects on sediment methanogenesis. Aquat Bot 61:165–180
Kim YS (2008) Evaluation of the water purification capacity in Iris pseudacorus and Acorus calamu. Korean J Hortic Sci Technol 26:172–176
Liu WJ, Zhu YG, Smith FA, Smith SE (2004) Do iron plaque and genotypes affect arsenate uptake and translocation by rice seedlings (Oryza sativa L.) grown in solution culture? J Exp Bot 55:1707–1713
Liu YJ, Zhu YG, Ding H, Guo W, Chen Z, Liu WJ (2007) The effect of root surface iron plaque on Pb uptake by rice (Oryza sativa L.) roots (in Chinese). Environ Chem 26:328–331
Manios T, Stentiford EI, Millner P (2003) Removal of heavy metals from a metaliferous water solution by Typha latifolia plants and sewage sludge compost. Chemosphere 53:487–494
Mendelssohn IA, Kleiss BA, Wakeley JS (1995) Factors controlling the formation of oxidized root channels: a review. Wetlands 15:37–46
Mihai M, Lacatusu I, Murariu A, Meghea A (2007) Quantification of lead pollution in Bucharest area and potential risks for human health. Environ Eng Manage J 6:473–478
Mitsui S, Kumazawa K, Yazaki J, Hirata H, Ishizuka K (1962) Dynamic aspects of N, P, K uptake and O2 secretion in relation to metabolic pathways within the plant roots. Soil Sci Plant Nutr 8:25–30
Otte ML, Rozena J, Koster L, Haarsma MS, Broekma RA (1989) Iron plaque on the roots of Aster tripolium L.: interaction with zinc uptake. New Phytol 111:309–316
Overesch M, Rinklebe J, Broll G, Neue HU (2007) Metals and arsenic in soils and corresponding vegetation at Central Elbe river floodplains (Germany). Environ Pollut 145:800–812
Parys E, Romanowska E, Siedlecka M, Poskuta JW (1998) The effect of lead on photosynthesis and respiration in detached leaves and in mesophyll protoplasts of Pisum sativum. Acta Physiol Plant 20:313–322
Piccardi EB, Clauser M (1983) Absorption of copper by Iris pseudacorus. Water Air Soil Pollut 19:185–192
Sheoran AS, Sheoran V (2006) Heavy metal removal mechanism of acid mine drainage in wetlands: a critical review. Miner Eng 19:105–116
St-Cyr L, Crowder AA (1989) Factors affecting iron plaque on the roots of Phragmites australis (Cav.) Trin ex Steudel. Plant Soil 116:85–93
Sundby B, Vale C, Cacador I, Catarino F, Madureira MJ, Caetano M (1998) Metal-rich concretions on the roots of salt marsh plants: mechanism and rate of formation. Limnol Oceanogr 43:245–252
Taylor GJ, Crowder AA (1983) Use of the DCB technique for extraction of hydrous iron oxides from roots of wetland plants. Am J Bot 70:1254–1257
Violante A, Barberis E, Pigna M, Boero V (2003) Factors affecting the formation, nature, and properties of iron precipitation products at the soil-root interface. J Plant Nutr 26:1889–1908
Wang X, Zhou QX (2003) Distribution of forms for cadmium, lead, copper and zinc in soil and its influences by modifier (In Chinese). J Agro-Environ Sci 22:541–545
Weis JS, Weis P (2004) Metal uptake, transport and release by wetland plants: implications for phytoremediation and restoration. Environ Int 30:685–700
Weiss JV, Emerson D, Megonigal JP (2005) Rhizosphere iron(III) deposition and reduction in a Juncus effusus L.-dominated wetland. Soil Sci Soc Am J 69:1861–1870
Wieβer A, Kuschk P, Kätner M, Stottmeister U (2002) Abilities of helophyte species to release oxygen into rhizospheres with varying redox conditions in laboratory-scale hydroponic systems. Int J Phytorem 4:1–15
Yang B, Lan CY, Yang CS, Liao WB, Chang H, Shu WS (2006) Long-term efficiency and stability of wetlands for treating wastewater of a lead/zinc mine and the concurrent ecosystem development. Environ Pollut 143:499–512
Ye ZH, Baker AJM, Wong MH, Willis AJ (1997) Zinc, lead and cadmium tolerance, uptake and accumulation by Typha latifolia L. New Phytol 136:469–480
Ye ZH, Wong MH, Baker AJ, Willis AJ (1998) Zinc, lead and cadmium accumulation and tolerance in Typha latifolia as affected by iron plaque on the root surface. Aquat Bot 61:55–67
Zhang XB, Liu P, Yang YS (2007) Phytoremediation of urban wastewater by model wetlands with ornamental hydrophytes. J Environ Sci 19:902–909
Acknowledgments
We thank Prof. Randy Dahlgren from the University of California, Davis for helping to carefully revise this manuscript. This work was jointly supported by the Science and Technology Project of Zhejiang Province (2005E10047, 2005C13003), the National Nature Science Foundation for Distinguished Young Scholars of China (40425007), Aid Program for Science and Technology Innovative Research Team in Higher Educational Institutions of Hunan Province, and the Science Foundation of Hengyang Normal University, China.
Author information
Authors and Affiliations
Corresponding authors
Additional information
Responsible editor: Ji-Zheng He
Rights and permissions
About this article
Cite this article
Zhong, S., Shi, J. & Xu, J. Influence of iron plaque on accumulation of lead by yellow flag (Iris pseudacorus L.) grown in artificial Pb-contaminated soil. J Soils Sediments 10, 964–970 (2010). https://doi.org/10.1007/s11368-010-0213-7
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s11368-010-0213-7