
Abstract
Purpose
Life Cycle Impact Assessment methodology is still lacking a procedure that relates phosphorus emission to ecological damage in freshwater ecosystems. The aim of this study is to apply new insights in the characterization of aqueous eutrophication at the end-point level. Characterization factors for freshwater eutrophication in European waters caused by emissions of phosphorus to agricultural soils and freshwater were developed. The characterization factors are representative for emissions to the 101 most important European river catchments west of the Ural Mountains.
Methods
We combined site-generic fate factors of total phosphorus, calculated by means of the integrated assessment model CARMEN, with damage factors based on a concentration–response relationship between the concentration of total phosphorus and occurrence of macrofauna species in freshwaters. Environmental fate processes, such as surface run off, groundwater drainage, and hydrological freshwater residence times, are included in the fate factor which relates emission of phosphorus from wastewater treatment plants and due to agricultural supply of manure and fertilizer, to concentrations in freshwater.
Results and discussion
The product of fate factor and damage factor constitutes the characterization factor at the endpoint level with the following results: 1.1·103, 1.2·103, and 2.1·104 disappeared fraction of species·m3·day/kg phosphorus emission for manure, fertilizer, and sewage treatment plants, respectively. Normalization factors are based on the emission of total phosphorus in Europe resulting in 60.1 disappeared fraction of species·m3/person with a relative contribution of 16% by manure application, 18% by fertilizer application, and 66% by sewage treatment plant emissions.
Conclusions
From intervention (P emission) to ecological damage of inland waters, most relevant site-specific processes are included to derive a characterization factor at the damage level. Although the characterisation factor for P due to agricultural application is a factor of 20 lower compared to emissions to freshwater, the nutrient enrichment of European freshwaters is still for one third attributed to agricultural application of phosphorus.
Similar content being viewed by others

Explore related subjects
Discover the latest articles, news and stories from top researchers in related subjects.Avoid common mistakes on your manuscript.
1 Introduction
Eutrophication in inland waters, caused by human activities, is one of the major factors that determine its ecological quality. Nutrient enrichment of freshwaters accounts for about half of the impaired lake area and 60% of the impaired river reaches in the USA (Carpenter et al. 1998). Phosphorus enrichment in the Dutch situation ranks higher as cause of ecological damage in surface waters than exposure to toxic substances (RIVM 2000). Uncontrolled algae bloom is often the result of too high input of nutrients so ultimately hindering other requirements for aquatic life such as light and oxygen. This may cause a chain of adverse ecological effects. Both inland and marine waters are subject to this form of water pollution due to different sources and substances. Algal growth is controlled by many factors and has seasonal dimensions. In northern Europe, the growth is greatly reduced or negligible during the winter period of low light and temperature. The availability of phosphorus or nitrogen merely regulates algae growth. For inland waters in temperate zones, phosphor is most often the limiting nutrient whereas in marine waters it is nitrogen (Crouzet et al. 1999). The natural P cycle is the predominant source of phosphorus hence growth of phytoplankton depends on its availability. In densely populated regions, however industrial and agricultural sources exceed natural inputs by far (Kristensen and Hansen 1994). As a result, an additional amount may lead to undesirable growth of phytoplankton.
Characterization of aquatic eutrophication in life cycle impact assessment (LCIA) typically only takes into account those nutrients that are limiting the growth of aquatic biomass. Using the Redfield ratio (Redfield et al. 1963), Heijungs et al. (1992) have proposed characterization factors for aquatic eutrophication. They ignored however the ultimate fate of phosphorus. Most often only a small fraction of eutrophying emissions will reach the aquatic ecosystem where it has an adverse effect. Huijbregts and Seppälä (2001) introduced fate analysis in the derivation of aquatic eutrophication potentials. Seppälä et al. (2004) emphasized the importance of a site-specific approach for aquatic eutrophication. They distinguished several nutrient forms in various sectors in Finland that determine the choice of a characterization factor. Potting et al. (2005) considerably improved the methodology by using an integrated assessment model (Klepper et al. 1995; Beusen et al. 1995) to estimate the fate of P (and N) emitted from each region or country in Europe separately. Fractions of the P emission flux that actually reach freshwater were calculated. For phosphorus, this fraction is approximately 0.05 and varies between 0.02 and 0.15, depending on the emission country. It has not been feasible in Life Cycle Assessment until now to relate the phosphorus concentration to effects on the biodiversity in inland waters. An ecological damage function for nutrient enrichment in terms of “disappeared fraction” (DF) of species as a function of the P concentration has not been formulated yet for LCIA.
The goal of this study is to develop characterization factors for phosphorus emissions to agricultural soils and directly to freshwater causing freshwater eutrophication. First, we relate nutrient emission—through a fate factor—to an increased concentration in receiving water bodies with help of the CAuse effect Relation Model to support Environmental Negotiations (CARMEN) model. Second, we introduce the damage factor derived from the disappeared fraction of macro-invertebrates that depends on the total phosphorus concentration, according to Struijs et al. (2010). The volume-weighted averaged product of fate factor and damage factor constitutes the characterization factor at the endpoint level.
2 Methodology
2.1 Framework
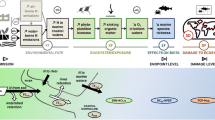
There are various routes how phosphorus compounds can enter the water compartment. Communal and—of minor importance—industrial wastewater treatment plants are the main point sources of phosphorus directly to the receiving water bodies. Emission through nutrient supply on agricultural land is a second (diffuse) source. Different fractions of P become available for uptake by crops and transport after supply of manure or fertilizer to agricultural soils. P-containing nutrients enter water bodies by surface run-off and erosion and by leaching from soils and subsequent transport to surface water through groundwater drainage (see Fig. 1). Direct deposition of airborne phosphorus on soil and on inland surface waters is negligible. Airborne phosphorus contributes less than 3% to waterborne emission (Berdowski and Jonker 1994).

CARMEN model: main sources for P emission to soil and surface waters; transport routes to the aquatic environment
The fate factor (FFk,j) represents the volume-weighted change in exposure concentration in river j due to a marginal change in emission rate by source k of phosphorus in all emission regions/countries simultaneously. Potting et al. (2005) have showed that the variability was relatively low in the environmental fate factors of phosphorous. It appeared less than a factor of three for communal sewage treatment plants and less than a factor of seven for agricultural emissions. Instead of deriving country-specific fate factors or characterization factors, we calculated a site-generic, European-average endpoint characterization factor for three emission routes, i.e., supply of manure and fertilizer to agricultural soil and direct emission of phosphorus from communal sewage treatment plants (STP). The dimensions of FFk,j is time (in days) and represents the effective residence time in freshwater j of emitted phosphorous.
The fate factor is multiplied by an ecological damage factor (EDFj) which is specific for freshwater j. EDFj is the marginal shift in the disappeared fraction of species in river j (with total phosphorus concentration C P,j) due to a marginal change in C P,j. The ecological damage factor, EDFj, has units of disappeared fraction·m3/kg.
The characterization factor of phosphorous emissions of source type k (manure, fertilizer, or sewage treatment plant) at the endpoint level (CFk) for freshwater eutrophication is the summation of the group FFk,j × EDFj over all European rivers j Eq. 1:
The characterization factor is the quotient of the (volume weighted) marginal increment of the fraction of disappeared macro-invertebrates in river j and a small increase in emission rate simultaneously in all countries (for source category k), summed over all rivers j. Therefore, the dimension of CF is disappeared fraction·m3·day/kg phosphorus emission.
2.2 Fate factor
CARMEN is a geographic information system (GIS)-based integrated assessment model, applied to assess the transnational implications of nutrient fluxes in eutrophication management. It calculates the change in nutrient loads in ground water, inland waters and coastal seas from changes in nutrient emissions (Beusen 2005; Beusen et al. 1995; Klepper et al. 1995). It evaluates the transport of nutrients from agricultural supply (or due to atmospheric deposition) through groundwater drainage and surface runoff spatially resolved over 124,320 grid elements of 1/6 × 1/6°. It also models straightforwardly the transport of nutrients emitted by wastewater treatment plants. In CARMEN, Europe includes 32 countries (Annex A) where emission takes place. The area subdivides into 101 river catchments (Annex B). The emissions are treated grid-wise in CARMEN which especially for application of manure and fertilizer has the advantage that all location specific circumstances are taken into account, such as characteristics of the soil, the slope and precipitation.
In the computation with the model CARMEN, Europe west of the Ural in 1995 was chosen as a reference system. Using CARMEN in the derivation of fate factors has the advantage that it is both accurate and practical as the model deals with gross supply of manure and fertilizer while accounting for widely varying agricultural fields in Europe. The major phosphorus sources that enrich surface water (Fig. 1) are agricultural supply of manure and fertilizer (diffuse sources) and effluent from wastewater treatment plants (point sources). Here we applied CARMEN to calculate the concentration increment in inland waters due to a small increase of gross supply of manure or fertilizer or due to a small increase of discharge of effluent by STP.
Separately for each source (manure, fertilizer, or STPs), denoted as intervention k, the consequences of an increase in emission is evaluated with respect to the reference scenario. The fate factor is the ratio of a volume weighted marginal increment in phosphorus concentration in a river and a marginal emission increase or application of phosphorus across whole Europe.
The fate factor FFk,j (days) with respect to river catchment j and for source category k is here defined as the marginal increase in concentration in inland water of river j (ΔC P,j kg/m3), multiplied with the volume of river j (m3), due to a marginal (1%) increase of emission ΔMk (kg/day) in all 32 European countries Eq. 2:
2.3 Ecological damage factor
The ecological damage factor, EDFj, follows from an empirical relationship between the disappeared fraction of macro-invertebrates and the aqueous phosphorus concentration. This relationship proposed by Struijs et al. (2010) rests on field data from 1980 until 2005 stored in the Limnodata Neerlandica. They used these field data to analyze the occurrence of macro-invertebrates species for varying total P concentration under different circumstances. Their analysis showed that there is a concentration (0.1 mg/l) with an optimum number of occurring macro-invertebrate genera (Nmax). Below or above 0.1 mg/l the number of occurring macro-invertebrate genera (N) is lower than Nmax. Therefore at the phosphorus concentration of 0.1 mg/l, the DF of macro-invertebrate genera was set to zero by Struijs et al. (2010) according to their definition of DF (= 1–N/Nmax.). They have shown that above 0.1 mg/l DF increases with increasing C P according to a sigmoid function (Fig. 2). According to Smith et al. (2003) phosphorus concentrations higher than 0.1 mg/L are usually of anthropogenic origin and from a generic point of view concentrations above 0.3 mg/L are considered a potential cause of encroachment of aquatic life due to nutrient enrichment. Fitting a logistic equation through the points that represent 1-N/Nmax above 0.3 mg/l (circles in Fig. 2) gives Eq. 3:

The disappeared fraction of macro-invertebrate genera versus the logarithm of the phosphorus concentration (modified from Struijs et al. (2010))
Struijs et al. (2010) derived Eq. 3 from data on the potential occurrence of macro-invertebrate genera at phosphorus concentrations above 0.3 mg/l. This is half an order of magnitude above 0.1 mg/l, the concentration above which an anthropogenic contribution is very likely (Smith et al. 2003). Here we apply the DF function also between 0.1 and 0.3 mg/l and consider Eq. 3 valid for concentrations above 0.1 mg/l. Although water bodies with lower concentrations than 0.1 mg/l may have undergone some anthropogenic influences with respect to phosphorus input, the disappeared fraction due to nutrient enrichment by P was set to zero.
In deriving a damage factor, we focused on macro-invertebrates only, neglecting the potentially adverse effect of phosphorus concentrations on other taxonomic groups in fresh waters. However, numerous genera of macro-invertebrates occur in many types of freshwater systems, each having its own specific environmental requirements and sensitivity to stressors (Van den Brink and Ter Braak 1998; Dyer and Belanger 1999). Macro-invertebrate diversity is therefore technically suitable to indicate deterioration of the aquatic ecosystem. Struijs et al. (2010) combined a set of 1,056,194 records with 2,790 different macrofauna taxa with a set of 265,959 records of total phosphorus concentrations occurring in the Limnodata Neerlandica. In the analysis, they deliberately took as many as conditions with respect to, e.g., weather and river flow rate into account. The purpose was to derive a generic damage function that is based on macro-invertebrate occurrence and that is generally applicable to all freshwater systems in Europe. Struijs et al. (2010) have cautioned not to apply the P concentration—macro-invertebrate occurrence function to a water body of specific size and with a specific hydraulic retention time. Equation 3 is only valid for C P above 0.1 mg/l, although at lower concentrations the diversity of macro-invertebrates also declines (see Fig. 2) this is not a matter of eutrophication. We assume that the disappeared fraction of macrofauna species in fresh water is a generic indicator for ecological damage due to a eutrophying phosphorus concentration above 0.1 mg/l. The EDFj for river j was obtained by differentiation of Eq. 3. It is river specific because it depends on C P,j in river j (Annex B) according to Eq. 4:
EDF (Annex C) has dimensions of reciprocal concentration (e.g. m3/kg) The ecological damage factor, EDF, was set to zero for P concentrations below 0.1 mg/l. This implies that we do not account for a potential overall increase in species richness due to an increase in P for oligotrophic situations, as shown in Fig. 2. Figure 3 shows EDF as a function of C P above 0.1 mg/l.

EDFP, the ecological damage factor as function of the phosphorus concentration (C P > 0.1 mg/l); the insert shows EDFP in the relevant range of C P (<2 mg/l)
3 Results and discussion
Site-generic endpoint characterization factors were calculated for the three emission sources according to Eq. 1 by summing up the products of fate factor and damage factor listed for each river in the table of Annex C. The results are given in Table 1. In Annex B characteristics of 101 rivers included in CARMEN are given. Annex C contains for each river fate factors for phosphorus emission resulting from each emission source (k). Annex C also contains the ecological damage factor (EDFj) for each river j calculated from the total phosphorus concentration in each river (see Annex B). The relationship given by Eq. 3 and Fig. 2, on which EDFj and the characterization factor are based, is however only applicable in a generic way.
Dividing the total P emission in 1995 per source category by the number of Europeans provides the P emission per person (second column of Table 1). The product of these emission data and the characterization factor yields the normalization factor. As can be inferred from the last column of Table 1, the contribution of agricultural P emissions to nutrient enrichment of freshwater is approximately one third.
There is a remarkable difference between emission to soil and emission to freshwater. The first is the result of supply of nutrients to agricultural soil. Most of it is taken up by crops or sorbed in the soil. Fate factors regarding aquatic eutrophication due to supply of manure or fertilizer are related to gross supply of phosphorus. Net emission of phosphorus to soil takes only the share into account that is available to enrich surface water after uptake by plants and crops in the topsoil. This is approximately 5% (depending of the country or region) and therefore the gross fate factors reported here are far below the fate factor for emission source category STP (Annex C). Emission from this source is almost entirely available to increase the amount of phosphorus in freshwater.
Usually, in life cycle assessment, both the agricultural topsoil and wastewater treatment plants are considered part of the technosphere (Potting et al. 2005). As a consequence, inventory data for P are deemed proper if they include emission rates from the technosphere into the receiving environment. For point sources such as communal wastewater treatment plants, such net emission data are readily available and suitable in environmental fate analysis. For P supply to agricultural soil, however, this can be troublesome as most often only gross supply in terms of kg P/ha/year is known, i.e., emission to the topsoil (i.e., to the technosphere). Conversion from gross emission to agricultural soil into emission from the topsoil includes removal processes such as uptake by plants, binding to topsoil and erosion. The factor with which the characterization factor for gross P emission to soil should be multiplied is on average 19.33 if only net P emission rates are available. This is close to a simpler alternative: applying the characterization factor for STPs directly. The fate processes, however, vary highly with the texture of the soil, slope, and land use. Therefore fate factors, spatially resolved by means of the GIS based CARMEN model, is more accurate when dealing with gross supply of manure and fertilizer per European country.
One percent emission changed was chosen as an optimal marginal increment in emission rate of P from source k. This perturbation with respect to the reference scenario of 1995 caused a linear response. An increase in P emission below 1% yielded very low increments in concentration of P because the calculation procedure is prone to threshold effects.
The damage factor was derived from an empirical relationship between the abundance/occurrence of macro-invertebrate genera and total phosphorus concentrations, observed in Dutch inland waters. The diversity of macro invertebrates is often considered to be a suitable and sensitive indicator for describing the ecological condition of freshwater because these organisms are relatively abundant and diverse and often display a high but species-dependent sensitivity to changing environmental conditions (e.g., Metcalfe 1989). We have applied this damage function for 101 catchments across whole Europe. This is justified because in temperate zones (Europe) phosphorus in inland water bodies is the limiting nutrient for algae and a too high algae growth due to P enrichment above 0.3 mg/l is a stress factor for macro invertebrates. In some oligotrophic rivers in Europe, the optimum total P concentration with respect to the highest macro-invertebrate diversity may be below 0.1 mg/l. For such oligotrophic waters, we may ask ourselves whether the highest diversity corresponds to a pristine aquatic ecosystem. Most often such waters have low volumes. Therefore, in a site-generic approach which covers 101 river catchments of which many have high volumes and are heavily loaded, this will not seriously affect the final result, being a site-generic characterization factor for phosphorus enrichment of European inland waters.
The question rises how these characterization factors compare to earlier reported work. Potting et al. (2005) found equivalency factors that were approximately a factor of seven higher for STPs than for agricultural emissions. In our work, the difference is almost a factor of 18 which is in accordance with the work from Huijbregts and Seppälä (2001) who found a difference of a factor of 20 in the fate factor of freshwater versus agricultural emissions of phosphorus.
4 Conclusions
Characterization factors representing the impact of phosphorous emissions on biodiversity of macrofauna of freshwaters in Europe were developed. These characterization factors account for spatial differences with respect to three emission sources of which two are related to gross emission to agricultural soil, i.e., through application of manure or fertilizer. We have evaluated emission and fate of these P inputs by means of the GIS-based model CARMEN that accounts for spatially differentiated emission rates, soil conditions, slope, precipitation, and land use. The third emission source stems from European households, either directly to water or via communal sewage treatment plants. We have computed a fate factor with respect to each river that accounts for different circumstances. The ecological damage function takes these differences in consideration as well. From intervention (P emission) to ecological damage of inland waters, most relevant site-specific processes are included.
The normalization factors indicate that ecological damage caused by anthropogenic emission of phosphorus in Europe is for one third attributed to agricultural production and for two third to discharge of communal wastewater.
References
Berdowski JJM, Jonker WJ (1994) Emission in the Netherlands. Industrial sectors, regions and individual substances (1992 and estimates for 1993; Publication series emission registration no. 21; in Dutch). Ministry of Housing, Spatial Planning and Environment, The Hague
Beusen A (2005) User manual of CARMEN1. National Institute of Public Health and Environmental Protection (RIVM), Bilthoven, Manuscript, not published
Beusen AHW, Klepper O, Meinardi CR (1995) Modelling the flow of nitrogen and phosphorus in Europe: from loads to coastal seas. Water Sci Technol 31(8):141–145
Carpenter SR, Caraco NF, Correll DL, Howarthet RW, Sharpley AN, Smith VH (1998) Nonpoint pollution of surface waters with phosphorus and nitrogen. Ecol Appl 8(3):559–568
Crouzet P, Leonard J, Nixon S, Rees Y, Parr W, Laffon, L, Bogestrand J, Kristensen P, Lallana C, Izzo G, Bokn T, Bak J, Lack TJ, Thyssen N (ed) (1999) Nutrients in European ecosystems. European Environment Agency, Copenhagen, Environmental assessment report, no 4
Dyer SD, Belanger SE (1999) Determination of the sensitivity of macroinvertebrates in stream mesocosms through field-derived assessments. Environ Tox Chem 18:2903–2907.
Heijungs R, Guinée JB, Huppes G, Lankreijer RM, Udo de Haes HA, Wegener Sleeswijk A, Ansems AMM, Eggels AMM, Van Duin R, De Goede HP (1992) Environmental life cycle assessment of products. Guidelines and backgrounds. Centre of Environmental Sciences, Leiden
Huijbregts MAJ, Seppälä J (2001) Life cycle impact assessment of pollutants causing aquatic eutrophication. Int J LCA 6(6):339–344
Klepper O, Beusen AHW, Bollen MSJ, Meinardi CR (1995) Modelling the flow of nitrogen and phosphor in Europe: from loads to coastal seas (RIVM report 461501004). National Institute of Public Health and Environmental Protection (RIVM), Bilthoven
Kristensen P, Hansen HO (1994) European rivers and lakes. Assessment of their environmental state. European Environmental Agency, Copenhagen, EEA environmental monographs 1
Metcalfe JL (1989) Biological water quality assessment of running waters based on macroinvertebrate communities: history and present status in Europe. Environ Pollut 60:101–139
Potting J, Beusen A, Øllgaard H, Hansen OC, De Haan B, Hauschild M (2005) Aquatic eutrophication. In: Potting J, Hauschild M (eds) Technical background for spatial differentiation in life cycle impact assessment. Danish Environmental Protection Agency, Copenhagen
Redfield AC, Ketchum BH, Richards FA (1963) The influence of organisms on the composition of sea water. In: Hill MN (ed) The sea, vol 2. Interscience, New York, pp 26–27
RIVM (2000) Environmental Outlook 2000–2030. Samson H.D. Tjeenk Willink bv, Alphen aan den Rijn. (in Dutch)
Seppälä J, Knuuttila S, Silvo K (2004) Eutrophication of aquatic ecosystems. A new Method for calculating the potential contributions of nitrogen and phosphorus. Int J LCA 9(2):90–100
Smith RA, Alexander RB, Schwarz GE (2003) Natural background concentrations of nutrients in streams and rivers of the conterminous United States. Environ Sci Technol 34:3039–3048
Struijs J, De Zwart D, Leuven, RSEW, Huijbregts MAJ (2010) Field sensitivity distribution of macroinvertebrates for phosphorus in inland waters. Accepted for publication in Integr Environ Assess Manage
United Nations Department of Economic and Social Affairs/Population Division (2004) World population to 2300. United Nations, New York
Van den Brink PJ, Ter Braak CJF (1998) Multivariate analysis of stress in experimental ecosystems by Principal Response Curves and similarity analysis. Aquat Ecol 32:163–178
Acknowledgements
The authors wish to thank Dr. H.J. van Wijnen (Laboratory for Ecological Risk Assessment, National Institute for Public Health and the Environment) for advising on demography.
Author information
Authors and Affiliations
Corresponding author
Electronic supplementary materials
Below is the link to the electronic supplementary material.
ESM 1
(PDF 255 kb)
Rights and permissions
About this article
Cite this article
Struijs, J., Beusen, A., de Zwart, D. et al. Characterization factors for inland water eutrophication at the damage level in life cycle impact assessment. Int J Life Cycle Assess 16, 59–64 (2011). https://doi.org/10.1007/s11367-010-0232-z
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s11367-010-0232-z