
Abstract
We conducted an experiment with two agricultural soils with acidic and alkaline pH levels to assess the effects of zinc oxide nanoparticles (nZnO) on the bacterial community. The effect of the nZnO concentrations (0, 0.1, 1, 10, 100, 1000 mg Zn/kg soil) and contact time between nanoparticles and soil (180 days) was considered. We measured the microbial respiration rate, nitrogen transformation, enzymatic activities (dehydrogenase (DH), acidic phosphatase (ACP), and alkaline phosphatase (ALP)), and the community-level physiological profile (CLPP) soil parameters. Respiration potential and nitrogen transformation were negatively affected only at the highest nZnO concentration. The changes in enzymatic activities were very variable with time and between both soils. A stimulating effect of the nanoparticles on microbial activity was clearly shown at 30 days after the nZnO application in both soils, except for the 1000 mg/kg in calcareous soil, after which time in the latter, the functional richness of the bacterial community was reduced to virtually zero. However, values of the enzymatic activities demonstrated that there was self-adaptation of microbial communities over the study period (180 days). The nZnO 1000 mg/kg dose produced an increase in bacterial growth in the acidic soil, without apparent changes in their metabolic profiles over time. A good correlation was found between microbial respiration rates (calcareous and acidic soils) and microbial metabolic activity (acidic soil) based on extracted Zn concentrations. Our findings suggest the necessity of additional studies to examine the effects of nZnO in natural microorganism populations in soil with different pH levels for extended periods of time.
Similar content being viewed by others

Explore related subjects
Discover the latest articles, news and stories from top researchers in related subjects.Avoid common mistakes on your manuscript.
Introduction
Zinc oxide nanoparticles (nZnO) have among the highest production volumes of nanoparticles, and therefore, their environmental levels are expected to rise constantly given the extensive application of this material (Piccinno et al. 2012). nZnO concentrations in the environment range from less than 100 μg/L in water to a few milligrams per kilogram in soil (Boxall et al. 2007). Harmful effects on soil microorganisms have been demonstrated, at very low concentrations (< 1 mg/kg) (Simonin and Richaume 2015). Single-species studies have highlighted varying sensitivities of bacteria to nZnO (Brayner et al. 2006; Jones et al. 2008; Negi et al. 2012; Simon-Deckers et al. 2009). Even although single-species in vitro studies are highly informative (Jiang et al. 2009; Roesch et al. 2007; Simon-Deckers et al. 2009), they lack the complexity of natural populations involving hundreds to thousands of interacting species. In consequence, there is a necessity to evaluate the risks related to nanoparticles in agricultural soils so as to preserve ecosystems.
Soil is a nonrenewable resource increasingly under environmental stress; for preserving soil characteristics, it is necessary to know the reaction of ecosystems to disruption and the resistance of soil microbial populations. Soil microbial communities play an important role in preserving ecosystem functions, soil health, and production. Slight changes in microbial diversity can affect environmental quality (Holden et al. 2014). A microorganism ecotoxicological approach, to estimate the effect of chemicals on natural associations of populations, provides a faithful evaluation of the response of microorganisms to contamination and leads to a more suitable environmental risk assessment. Changes in the functioning of soil microbial communities can be monitored by measuring enzyme activities, soil respiration potential, and nitrification activity, which have been linked to modifications in soil quality (Jimenez et al. 2002; Schloter et al. 2003).
Community-level physiological profiling based on carbon substrate utilization approaches has been widely used to characterize microbial communities of different soils. The calculated numbers of soil microorganisms are in the extent of 109 cells/g soil, with a variety of 104 species (Christensen et al. 1999; Curtis et al. 2002). The rapid development of microorganisms makes it possible to distinguish changes in microbial communities by measuring their metabolic responses to stressors (Garland and Mills 1991). However, little research has been conducted on the interaction between nanoparticles and agricultural soil microbial communities in vivo that would mimic the natural environment. Researching changes in soil microbial functioning is useful to understand how agricultural soils are impacted by nanoparticles.
The influence of soil characteristics on the nanoparticle toxicity is starting to be studied (Langdon et al. 2014; Read et al. 2016; Schlich and Hund-Rinke 2015; Simonin and Richaume 2015; Simonin et al. 2016). The description of soil parameters governing the bioavailability and toxicity of nanoparticles is decisive an environmental risk assessment. Soil pH, ionic strength, and the availability of natural organic matter seem to be important in metal nanoparticles mobility, dissolution, and bioavailability to soil microorganisms (Cornelis et al. 2014; Simonin and Richaume 2015; Smolders et al. 2004). pH affects the availability of nutrients, modifies the balance of the solid phase, and controls the association and diversity of the microbial community.
Risk assessment recommendations take into account studying nanoparticle in natural environments to know in what way nanoparticles relate with agricultural soil components and the capacity of microorganisms to be resistant to short- and long-term nanoparticle disruption. The aim of this work was to study the role that pH plays in mediating the dose–response effect of nZnO on agricultural soil bacterial communities, using concentrations ranging from 0.1 to 1000 mg/kg dry soil and long-term monitoring, to imitate realistic exposure of soils to nZnO.
Materials and methods
Characterization of nZnO and soils
Zinc nanopowder (< 100 nm) was obtained from Sigma-Aldrich (Germany), the characteristics of the constituent nZnO were determined previously (Fernandez et al. 2013). The size and shape of the nanoparticles were determined with a transmission electron microscopy (TEM). The particle size distribution was determined with a Nano-Zetasizer (1000 HS, Malvern Instrument Ltd., UK), which uses the dynamic light scattering (DLS) technique.
Two different soils with different pH levels were selected, an acidic soil (pH 5.4) and a calcareous soil (pH 8.5), and were collected from the surface layer (0–20 cm) of two agricultural fields located in Madrid (Spain). The soils were air-dried and sieved (2 mm mesh) prior to characterization. Their chemical properties were determined by standard methods, and the analytical results are shown in Table 1.
Sample preparation
The soils were spiked with 0.1, 1, 10, 100, and 1000 mg Zn/kg soil as nZnO. The Zn concentrations were calculated on a soil dry weight (DW) basis. Untreated soils were used as controls. The nZnO powders were added directly to the soil, following method in previous studies (Garcia-Gomez et al. 2015), and hand blended.
Samples (soil with and without nZnO treatments) were split into two groups with three replicates per group and treatment. The first group was used to assess the effects of nZnO on respiration potential, enzymatic activities, and carbon source utilization by soil microbial community. The second group was used to determine the nitrification activity and received an organic nitrogen source (5 g Lucerne/kg soil). Polyethylene containers (each one with a capacity of 10 L, a mean internal diameter of 24 cm, and a height of 24 cm) were filled with 600 g (DW) of control and treatment soils. The containers were incubated for 180 days (20 ± 2 °C, dark, 40–60% water holding capacity). The moisture of the samples was controlled by reference to weight. Subsamples were taken at 1, 30, 90, and 180 days to analyze the microbial parameters and soil zinc availability.
Soil chemical analyses
The total Zn in the soil samples was determined after treating 1 g of dried sample with an acid mixture of HNO3:HF:double deionized water (1:1:1) followed by digestion in Teflon bombs in a microwave oven (CEM Corporation, model-Mars, Matthews, NC, USA). Two certified reference soils provided by the Institute for Reference Materials and Measurements of the European Commission (ERM-CC141 and BCR-143R) were used to verify the quality of the results for total Zn contents.
The actual available fraction of Zn applied to soil was estimated with the CaCl2 extraction (Zn–CaCl2), sometimes referred as the effective bioavailable metal fraction. The air-dried soil samples were extracted for 2 h with a 0.01-M CaCl2 solution (pH 5.7, without buffer) at 20 °C in a 1:10 extraction ratio (w:v) (Houba et al. 2000).
Microbial parameters analyses
Microbial respiration
Effects on carbon transformation activity of soil microorganisms were determined following the principles of standardized methods (OECD 2000a, 217). Soil subsamples (5 g) used for determining microbial respiration rates induced by glucose were amended with 4 mg glucose/g soil (DW), and the carbon dioxide released was measured using a BacTrac 4300 SY-Lab (Microbiological Analyzers).
Nitrogen transformation test
The effects on nitrogen transformation activity of soil microorganisms were determined using the standardized method of the OECD (2000b, 216). Nitrate was extracted from the soil samples (10 g) by shaking with 50 ml of 0.1 M potassium chloride solution for 1 h. Then, the mixture was centrifuged at 3000 rpm, 5 min, and filtered. Nitrates were determined using the NitriVer® 3 and NitraVer® 5 reagents in a Beckman DU530 spectrophotometer at 507 and 400 nm, respectively.
Enzymatic activities
Dehydrogenase activity (DH) was measured according to the method of Rogers and Li (1985). Samples of alfalfa-enriched soils (1 g) were mixed with 0.2 ml of 3% (w/v) of 2,3,5-triphenyltetrazolium chloride (TTC) and 0.5 ml of glucose 0.5% (w/v) both dissolved in deionized water (calcareous soil) or in tris(hydroxymethyl)-aminomethane (TRIS) buffer pH 7.4 (acidic soil). The test tubes were capped and incubated at 27 °C in the dark for 24 h. After incubation, 2.5 ml of methanol was added to extract the reaction product (triphenylformazan) and the mixture was stirred and centrifuged at 3500 rpm, 4 min. The absorbance was measured at 490 nm in a GENios microplate spectrofluorometer (Tecan, Switzerland) using a standard curve of reagent grade triphenylformazan (TPF). The dehydrogenase activity was calculated as micrograms of the TPF formed per gram of soil during 24 h.
Phosphatase (acidic and alkaline) activities were measured according to the method of Freeman et al. (1995). Briefly, 1 g of soil sample was preincubated at 27 °C for 30 min with 3.2 ml of buffer (50 ml of NaOH 1 M, 1.21 g of TRIS, 1.16 g of maleic acid, 1.4 g of citric acid, and 0.63 g of boric acid in 500 ml of water) adjusted at pH 6.5 or pH 10 for the acidic or alkaline phosphatase, respectively. Then, 80 μl of a solution 0.01 M of methylumbelliferyl phosphate (MUP) in buffer is added, shaken, and incubated at 27 °C in the dark for 2 h. The mixture was centrifuged at 3000 rpm, 4 min and the fluorescence intensity of 4-methylumbelliferone (MU) formed is measured at the excitation wavelengths of 320 and 360 nm for acidic and alkaline phosphatase, respectively, and emission wavelength of 465 nm, in a GENios microplate spectrofluorometer (Tecan, Switzerland) using a standard curve of regent grade MU. The activities were calculated as micrograms of MU formed per gram of soil.
Microbial CLPP analysis
CLPP were obtained using Biolog® EcoPlates established methods (Ge et al. 2011). Five-gram portions of sample were suspended in a sterile sodium chloride solution (0.9% w/v). The cellular suspension was diluted (1:100) in the sterile sodium chloride solution and subsequently dispensed (125 μl) into each well of Biolog® EcoPlates and incubated at 25 °C in the dark. Data were recorded with a plate reader SPARK 10 M (Tecan, Switzerland) at 590 nm every 24 h until 144 h.
Three indices were determined. Firstly, average well color development AWCD = ∑ODi/95, where ODi is the absorbance value from each well, corrected by subtracting the blank well values from each plate well, and 95 is the number of total substrates. Secondly, the substrate richness (S) was calculated as the number of wells with a corrected absorbance greater than 0.20. Thirdly, the Shannon index (H) or “diversity” showing the substrate utilization model was calculated as follows: H′ = − ∑pi ln (pi), where pi is the ratio of the activity on each substrate (ODi) to the sum of activities on all substrates ∑ODi.
Statistical analysis
Mean values were taken from each triplicate data set. To clarify the presentation of the results, the enzymatic activity data of treated soil samples were expressed as the percentage of the enzymatic activity compared to the control sample. Statistically, significant differences between individual means for chemical and toxicological data were identified by analysis of variance (ANOVA) with Fisher’s least significant difference procedure (LSD, P < 0.05). Pearson’s correlation coefficients (n = 3) were calculated to relate enzymatic activity, microbial activity, and functional microbial diversity to CaCl2 extractable concentrations of zinc in calcareous and acidic soils. The data were analyzed statistically using the STATGRAPHICS software (version 5.0).
Results and discussion
There have been very few studies conducted to determine the soil pH effect on the toxicity of zinc oxide nanoparticles with the soil microbial community. To the best of our knowledge, this is the first study to evaluate nZnO toxicity on agricultural soil bacterial communities, using concentrations that range from 0.1 to 1000 mg/kg dry soil and long-term monitoring (180 days), to reproduce realistic soil exposure to nZnO.
Soil pH seems to be the principal factor that governs concentrations of soluble metals. Zn solubility correlates negatively with pH (Sauve et al. 2000). Moreover, clay content in calcareous soil was higher than in acidic soil, which also helps increase Zn retention in soil and, hence, decrease Zn availability to microorganisms, as clays are believed to adsorb zinc ions across both ion exchange and specific adsorption (Farrah and Pickering 1997). Zinc availability has also been connected to soil organic matter content. In this study, the soil organic matter content effect of content on Zn availability to microorganisms should be considered insignificant because values were very low and similar in both soils (< 2%).
Soil chemical analysis
Since zinc is both an essential trace element and possible toxicant, the effects of zinc fixation in soil are of possible significance. The available Zn fraction was remarkably higher in acidic than in calcareous soil (Table 2). Significant differences (p < 0.001) in CaCl2 extractable Zn were observed between acidic and calcareous soils at 30, 90, and 180 days after the nZnO application (DAA). Zinc sorption to the soil and ZnO solubility are negatively correlated with pH, which is well defined by the competitive adsorption model (Sauve et al. 2000). In addition, the clay content was higher in the calcareous soil than in acidic soil, which increased the retention of Zn in soil. At the nZnO loading rate of 1000 mg/kg, the available Zn fraction in calcareous soil decreased with the incubation time, with values of 0.12, 0.08, 0.05, and 0.05% of total added Zn, after 1, 30, 90, and 180 DAA, respectively. As time passed, some of the adsorbed Zn may disperse into soil aggregate particles and thus lose its extractability. The results agreed with previous work by Donner et al. (2010) that Zn availability in soil was affected with time. The available Zn fractions from the acidic soil were 52, 45, and 29% with the loading rates of 10, 100, and 1000 mg/kg, respectively, and those percentages were consistent across time (Table 2). At low pH, there is a high Zn concentration in solution (as Zn2+ and Zn(OH)+), because of the increased Zn dissolution and the pH influences on Zn solubility. Tipping et al. (2003) found similar correlation with higher Zn solution concentrations in lower pH soils than in more neutral soils.
The pH analysis of soils showed no change between 1 and 180 DAA. With the increasing nZnO concentration in the acidic soil, an increase of around 1 pH unit in relation to the control took place. The dissolution of Zn2+ ions from nanoparticles reduced protons and raised soil pH. Tourinho et al. (2013) and Waalewijn-Kool et al. (2013) have described a similar pH increment after nZnO additions.
Microbial parameter analysis
Microbial respiration
The effects on carbon transformation activity of soil microorganisms were measured in calcareous and acidic soils to evaluate the repercussions of different doses of nZnO on overall microbial activity. Altogether, the change in microbial respiration potential in nZnO-contaminated calcareous and acidic soils was small (Fig. 1). Soil respiration potential decreased at 10, 100, and 1000 mg/kg nZnO loading rates in calcareous soil, and at 100 and 1000 mg/kg in acidic soil, and in both soils, the differences were less than 10% and were consistent across time. Soil pH seems to be directly or indirectly one of the main factors affecting the behavior of nanoparticles in respiration potential of soil organisms (Heggelund et al. 2014; Waalewijn-Kool et al. 2013). In this work, the pH of calcareous and acidic soils did not change the soil glucose-induced respiration rate substantially (Fig. 1), the sole exception being the 1000 mg/kg treatment in which the respiration rates were significantly reduced in both calcareous (p < 0.001) and acidic (p < 0.002) soils. Inhibition in soil substrate-induced respiration has already been reported at high dose of nZnO by other authors (Ge et al. 2011). The answer of soil microbial communities to the administration of nZnO has been shown to be highly diverse and dependent upon the characteristics of the systems and the form of zinc applied (Collins et al. 2012; Ge et al. 2011, 2012; Rousk et al. 2012). Other researches described that substrate-induced microbial respiration was not sensitive in assessing the effect of metals in soil (Hund-Rinke and Simon 2008), possibly because the modified community may incorporate taxa that are functionally compensating with the taxa in the original group.

Changes in respiration over time due to nZnO doses in soil at the nominal concentrations are shown (0.1–1000 mg/kg soil) (error bars = ± SE, n = 3). All results are described as the percent of activity relative to the average activity of unexposed controls. Within each treatment group, means sharing the same letter are not significantly different (LSD test, p < 0.05)
Nitrogen transformation test
Soil inorganic-N availability is one of the most sensitive microbial processes that can be affected by contamination (Dalzell et al. 2002). The nitrogen transformation test was carried out to investigate the effects of nZnO on the nitrogen transformation activity of soil microorganisms. Figure 2 shows the changes in nitrate concentrations over time in calcareous and acidic nZnO-contaminated soils. The nitrification rates increased with increasing soil pH. The nitrification in the acidic soil was significantly lower than that in the calcareous soil at 90 and 180 DAA (p < 0.01). The addition of 1000 mg/kg of nZnO caused inhibition of nitrification at 1, 30, 90, and 180 DAA in the acidic treated soil, with inhibition rates of 36, 16, 17, and 14%, respectively. The lower values of nitrification in acidic treated soil may be related to the low immobilization and high bioavailability of Zn, which determines their negative effect on the nitrifying microorganism populations. At 1 and 30 DAA, the inhibition rates of nitrification in the 1000 mg/kg nZnO-treated calcareous soil were 13 and 22%, respectively (Fig. 2). Over time, the inhibition decreased and the nitrate concentrations recovered, with acidic soil values around 100% (except the 1000 mg/kg) and calcareous soil values around 125% at 180 DAA. The effect of nanoparticles addition to calcareous soil on nitrate concentrations indicated that there were significantly (p < 0.001) higher values at 180 DAA compared with 1 DAA. These results may be related to the adaptation of soil microorganism to the metals (Bruins et al. 2000). Also, toxicity of nZnO in the soil may decrease with time due to aging or equilibration processes (Waalewijn-Kool et al. 2013).

Changes in nitrification over time due to nZnO doses in soil at the nominal concentrations are shown (0.1–1000 mg/kg soil) (error bars = ± SE, n = 3). All results are described as the percent of activity relative to the average activity of unexposed controls. Within each treatment group, means sharing the same letter are not significantly different (LSD test, p < 0.05)
Enzymatic activities
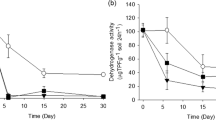
The toxicity originates from the heavy metals affecting the metabolism, morphology, and growth of soil microorganisms. The effects of nZnO on soil microorganisms were evaluated through dehydrogenase (DH), acidic phosphatase (ACP), and alkaline phosphatase (ALP) activities. DH activity shows the respiratory activity of soil populations through a reduction reaction (Voet et al. 2006). ACP and ALP phosphatases participate in the phosphatase cycle, catalyzing the hydrolysis of glycosidic bonds and the transport of phosphate groups to other compounds (Cox et al. 2000). In the calcareous soil, the nZnO negatively affected the electron transfer by inhibiting the dehydrogenase activity as soon as the nanoparticles were applied (Fig. 3a). DH activity was significantly (p < 0.05) reduced (nZnO-spiked vs. un-spiked soil samples) at 1 (0.1–1000 mg/kg), 30 (100 and 1000 mg/kg), and 90 (0.1–1000 mg/kg) DAA with a range of variation between 2 and 28%. At 180 DAA, the DH activity increased in the 0.1–1000 mg/kg treatments, with percentages between 10 and 80%. Other authors associated the reduction of nanoparticles toxicity at the intermediate and high concentrations to the aggregation incidents (Baalousha 2009; Simonin et al. 2017). Furthermore, nanoparticle exposures may lead to resistant microbial populations over time (Simonin and Richaume 2015). In the acidic soil, the DH activity decreased significantly (p < 0.001) at 30 DAA, with inhibition percentages of between 22% (100 mg/kg) and 89% (1000 mg/kg). Significant (p < 0.01) reductions of 72% (1 DAA), 67% (90 DAA), and 73% (180 DAA) were observed at the highest (1000 mg/kg) nanoparticle dose. The DH activity increased in the 0.1–100 mg/kg nZnO-treated acidic soil, with induction percentages between 1–6, 20–26, and 6–12% at 1, 90, and 180 DAA, respectively (Fig. 3a). The nZnO negatively affected the electron transfer in acidic soil only by suppressing DH activity at the highest nanoparticles dose. In particular, more than 67% of the activity was inhibited from 1 to 180 DAA. This inhibition has already been described at this dose of nZnO in medium and low pH soils (Garcia-Gomez et al. 2015; Kim et al. 2013). DH activity value was significantly (p < 0.001) different (calcareous vs. acidic soil samples) at 1, 30, 90, and 180 DAA in the 1000 mg/kg treatments. The DH activity was more inhibited in the acidic soil than in the calcareous soil by nZnO. The greater inhibition of DH activity in the acidic treated soil may be related to the low immobilization and high bioavailability of Zn, which determines their negative effect on the bacterial community. The calcareous soil also had a higher clay content, which may have attenuated the effect on DH activity by nZnO stabilization (Frenk et al. 2013).

Differences in specific activity of a dehydrogenase, b acidic phosphatase, and c alkaline phosphatase over time due to nZnO doses in soil at the nominal concentrations are shown (0.1–1000 mg/kg soil) (error bars = ± SE, n = 3). All results are described as the percent of activity relative to the average activity of unexposed controls. Within each treatment group, means sharing the same letter are not significantly different (LSD test, p < 0.05)
In calcareous soil, the nZnO 1000 mg/kg dose significantly inhibited the activity of ACP at 30 (10%, p < 0.01), 90 (55%, p < 0.001), and 180 (40%, p < 0.001) DAA (Fig. 3b), while on the other hand, nZnO doses lower than 1000 mg/kg caused stimulation of the ACP activity at 1 (20–107%), 30 (9–27%), and 180 (10–16%) DAA. nZnO inhibited the ACP activity in acidic soil (Fig. 3b) with percentages of 0.3–23 and 19–28% at 1 and 180 DAA, respectively, depending on the concentration of nanoparticles, while in the 1000 mg/kg treatments, the inhibition amounted to 37% (p < 0.05), 20% (p < 0.05), 56% (p < 0.01), and 56% (p < 0.01) at 1, 30, 90, and 180 DAA, respectively.
In the case of ALP (Fig. 3c), inhibition was observed at 30 (6%), 90 (34%), and 180 (1%) DAA in calcareous soil, but only at the highest concentrations (1000 mg/kg). In contrast to calcareous soil, in acidic soil, nZnO caused statistically significant (p < 0.001) dose effect stimulation of alkaline phosphatase (the sole exception was 90 DAA at concentrations of 0.1 and 1 mg/kg) (Fig. 3c). In calcareous soil, only a general light stimulation was noted, which did not exceed 20% (1 DAA at concentration of 10 mg/kg). The ALP activity was the most different parameter between the two soils, with values very close to those of the controls in calcareous soils, and dose–response values in acidic soil that increased with time. At 180 DAA, ACP activity (Fig. 3b) was found predominantly in calcareous soil and ALP activity (Fig. 3c) in acidic soil, which is in disagreement with the findings of other authors (Dick and Tabatabai 1984; Eivazi and Tabatabai 1977). Soil enzyme assays incorporate physical, mineralogical, and chemical parameters to show a single response. Only chemical parameters associated to nZnO administration could be related to clarify the ALP activity dose–response effect in acidic soil. Uncommon dose–response relationships have been reported after testing the effect of nanoparticles on soil microbial populations and communities (Bernhardt et al. 2010; Ge et al. 2011; Rahmatpour et al. 2017; Simonin and Richaume 2015). For instance, Simonin et al. (2017) attributed the observed decrease of TiO2 nanoparticle toxicity at intermediate and high concentrations to nanoparticle aggregation. In other studies, low concentrations of nanoparticles were more toxic than high ones (Menard et al. 2011; Simon-Deckers et al. 2009; Simonin et al. 2017). Nannipieri et al. (2011) concluded that the analysis of the effect of heavy metals on soil phosphatase activities is difficult, because environmental factors (pH, redox conditions, and surface charges) are determinant for the activity of phosphatases. Consequently, small changes in soil conditions could have uncertain results.
In the present study, the induction and inhibition of enzymatic activities were very variable with time and between the calcareous and acidic soils. However, we detected a slight decrease in toxicity over time in the calcareous (DH, ACP, and ALP activities) and acidic (DH and ALP activities) soils at lower nZnO concentrations than 1000 mg/kg, which can be attributed to changes in the characteristics and reactivity of nanoparticles (aging) (Mudunkotuwa et al. 2012) and/or to adaptation to the metal-resistance of soil microorganisms (Speir et al. 1995). This suggests that the changes in enzymatic activities may not have been driven by changes in microbial abundance but by the adaptation of soil microbial populations to the nanoparticles (Bruins et al. 2000). Other authors have shown notable variations in soil microbial enzymatic activities associated with their susceptibility to heavy metal toxicity (Giller et al. 1998). These differences are based on several factors, including mainly procedure and soil environmental aspects (pH, organic matter, and content of minerals) (Aon and Colaneri 2001; Fernandez-Calvino et al. 2011; Huang et al. 2005; Peyrot et al. 2014). On the other hand, we did not detect the remarkable decrease in activity of dehydrogenase and phosphatases described by other authors at low nZnO concentrations (Josko et al. 2014; Kim et al. 2011; Simonin and Richaume 2015).
Microbial CLPP analysis
The color development in Biolog® EcoPlates was recorded at 24 h intervals until maximum absorbance values at 144 h, which is the time necessary to better identify the differences in microbial community function. The response includes the microbial activity in each microplate (average well color development, AWCD), the Shannon–Weaver index (H) (i.e., the functional diversity and evenness of the species present), and the microbial community functional richness index (R) (number of oxidized C substrates) using an OD of 0.20 as the start for positive response (Garland 1996). In the present study, EcoPlate data depicted marked differences in AWCD, R, and H values related to the soil pH (Fig. 4). Thirty days after nZnO application, the stimulating effect on the microbial activity was clearly shown based on the higher AWCD, Shannon–Weaver index, and Richness compared to the control (1 DAA), in the calcareous (except for 1000 mg/kg) and acidic soils. However, after that, the nZnO disturbed the calcareous soil microbial structure, with AWCD, R, and H index values lower than 0.01, 1, and 0, respectively. At 90 and 180 DAA, specific groups of bacteria were not adapted to the environmental stresses, and values of AWCD, R, and H index were reduced to virtually zero (data not shown). One possible mechanism may be the direct toxicity of nZnO on soil bacterial communities through membrane cell damage or released Zn ions. It has been revealed that mannitol addition has a suppressive effect on AWCD value (Yu et al. 2016). In other studies that worked with weak acidic (pH 6.0) soils, a strong effect of nZnO was observed over time, normally during periods that did not last more than 60 days (Ge et al. 2011). Collins et al. (2012) defined the reduction of microbial diversity due to the effect of nZnO contamination. These authors detected a dissolution/modification of the nZnO in a soil with an alkaline pH over a 30-day period that changed the microbial community structure. Substrate-induced respiration represents the entire spectrum of the catabolic functions of the microbial communities, while CLPP is confined to only a subpopulation of cultivable aerobic microorganisms. This possibly explains the differences between the microbial parameter analysis and microbial physiological profile results. Other authors have described how AWCD is not always associated with microbial respiration potential, because color development reflects the response of the bacterial community to substrates (Yao et al. 2000). Many bacteria produce extracellular substances that have been shown to alter nanoparticle behavior. Also, the aging of nanoparticles in the soil can affect their toxic potential due to adsorption and dissolution phenomena, and this applies to nZnO between 1 h in acidic soil and 30 days in calcareous soil (Wang et al. 2013). All together, these could contribute to explain the calcareous soil results after 30 DAA in this work. Except for the 1000 mg/kg dose, the AWCD, R, and H acidic soil values were lower than that of the calcareous soil (30 DAA), probably due to the high bioavailability of Zn relative to acidic soil. Soil pH has been established as connecting bacterial community composition and variety, with a relationship between pH and diversity identified in natural soils (Fierer and Jackson 2006; Read et al. 2016). Even though the precise mechanism is not known, it has been theorized as being due to bacterial taxa having moderately close growth sensitivities to pH (Rousk et al. 2010). Read et al. (2016) described a change in zinc nanoparticles exposed communities at a pH below 6, with a strong effect on the bacterial community composition with increasing Zn concentrations. Under high pH soil conditions, microbial populations were scarcely sensitive to zinc addition, which disagrees with our findings in calcareous soil, where the data of AWCD, Shannon–Weaver Index, and richness significantly reduced at 30 (only in the 1000 mg/kg treatment), 90, and 180 DAA. The divergence between calcareous and acidic soils with the 1000 mg/kg treatment could be explained by the higher nanoparticle dissolution and availability in acidic soil (Table 2). Similar differences in effects have been previously observed in a single soil at varying pH levels (Read et al. 2016). However, in our study, the bacterial communities in the acidic soil displayed a wide diversity of physiological potentials, as showed by the Shannon–Weaver and R Indices. One explanation is that the resistance of specific low pH bacterial communities may vary to zinc and could become less susceptible to Zn exposure than high pH communities, because the species that lack tolerance to imposed stress are eliminated and also due to the improvement of other species populations that prosper under acidic pH conditions. Another explanation may be the capability of bacteria to secrete the extracellular substances that protect microorganisms against stress. Extracellular polysaccharides and proteins protect the Escherichia coli from nZnO (Wang et al. 2016).

Changes in a average well-color development (AWCD), b richness, and c Shannon–Weaver index over time due to nZnO doses in soil at the nominal concentrations shown (0–1000 mg/kg soil) (error bars = ± SE, n = 3). Within each treatment group, means sharing the same letter are not significantly different (LSD test, p < 0.05)
The variation of the metabolic behavior of the microbial community should reveal a change in its composition. In order to determine whether the variations detected in the whole substrate utilization could be assigned to changes in the capacity of microorganism populations to degrade some specific carbon source, we estimated the ability of each soil to oxidize different classes of carbon sources incorporated in EcoPlates (i.e., amino acids, carboxylic acids, carbohydrates, polymers, and miscellaneous (phosphorylated amines and phenols)). Substantial differences were found in the categorized substrate utilization patterns of soil microbial communities (Fig. 5) after 30-day treatment. At 30 DAA, the carboxylic acids and polymers were the most intensively metabolized in calcareous soil, while the acidic soil showed increased utilization of carboxylic acids, carbohydrates, and polymers, over the study period of 6 months. The nZnO 1000 mg/kg dose produced an increase of bacterial growth in acidic soil, without changes in their metabolic profiles over time (Fig. 5). This finding indicates that the microbial populations were stable in the acidic soil. In contrast to our results, other authors have reported that both high Zn concentrations and decreased pH values had important consequences on the EcoPlate pattern of soil microbial populations (Knight et al. 1997). This divergence may be explained by the different characteristics of the soils studied in this work.

Categorized substrate utilization patterns of soils by the microbial communities from calcareous and acidic soils exposed to different nZnO treatments over time for amino acids (AA), carboxylic acids (CA), carbohydrates (CH), polymers (Pol), and miscellaneous (phosphorylated, amines, and phenols) (Mis). Error bars represent the means ± SE (n = 3). Overall utilization of the six categories of nZnO concentrations is reported above bars
Pearson’s correlation coefficients (r) were calculated to relate enzymatic activity, microbial catabolic activity (AWCD and Shannon index for the different groups of carbon compounds in Biolog® EcoPlates), and functional microbial diversity to CaCl2 extractable concentrations of zinc in calcareous and acidic soils. Significant degrees of correlation were found in the calcareous soil between DH (180 DAA), ACP (180 DAA), ALP (30 DAA) activities and microbial respiration potential (30, 90, and 180 DAA) and extractable Zn (Table 3). However, a good correlation between microbial catabolic activity and extracted Zn concentrations was not observed in calcareous soil at 30 DAA. In the acidic soil, a high number of significant correlations were obtained among the extractable Zn, microbial activity, and biochemical parameters at each sampling time. A good inverse correlation coefficient between the total Zn amount and the microbial respiration potential was found in both the calcareous and acidic soils over time, regardless of the pH value (Table 3). The results are analogous to correlations reported between extracted Zn concentrations and the respiration potential and enzymatic activities (Liao and Xiao 2007; Strachel et al. 2017). In our study, the AWCD values correlated with nZnO concentrations, but only in acidic soil over time. These differences can be attributed to low immobilization and high Zn bioavailability.
Conclusions
The study of nanotoxicity in soils is difficult and complex as a result of the many sorts of relations between nanoparticles and microorganisms and nanoparticles and their environment, all of which have been shown to modify toxicity in model systems. Our work was designed to investigate the different interactions between nZnO and calcareous and acidic agricultural soils. As expected, soil microbial enzyme activities were adversely affected by nZnO in acidic soil more than in calcareous soil. However, soil microbial catabolic activities were negatively affected in calcareous soil more than in acidic soil, despite their high metal sorption capability. This study clearly demonstrates that soil microbial parameters and microbial functional diversity present different resistances to different soil pH. Our findings underline the need for additional acute and chronic toxicity studies examining natural microbial communities of soil with large ranges in pH for extended periods of time to unravel the mechanisms for nZnO toxicity.
References
Aon MA, Colaneri AC (2001) II. Temporal and spatial evolution of enzymatic activities and physico-chemical properties in an agricultural soil. Appl Soil Ecol 18:255–270. https://doi.org/10.1016/S0929-1393(01)00161-5
Baalousha M (2009) Aggregation and disaggregation of iron oxide nanoparticles: influence of particle concentration, pH and natural organic matter. Sci Total Environ 407:2093–2101. https://doi.org/10.1016/j.scitotenv.2008.11.022
Bernhardt ES, Colman BP, Hochella MF, Cardinale BJ, Nisbet RM, Richardson CJ, Yin L (2010) An ecological perspective on nanomaterial impacts in the environment. J Environ Qual 39:1954–1965. https://doi.org/10.2134/jeq2009.0479
Boxall A, Chaudhry Q, Sinclair C, et al (2007) Current and future predicted environmental exposure to engineered nanoparticles. Technical report. Central Science Laboratory. York. http://randd.defra.gov.uk/Document.aspx?Document=CB01098_6270_FRP
Brayner R, Ferrari-Iliou R, Brivois N, Djediat S et al (2006) Toxicological impact studies based on Escherichia coli bacteria in ultrafine ZnO nanoparticles colloidal medium. Nano Lett 6:866–870. https://doi.org/10.1021/nl052326h
Bruins MR, Kapil S, Oehme FW (2000) Microbial resistance to metals in the environment. Ecotoxicol Environ Saf 45:198–207. https://doi.org/10.1006/eesa.1999.1860
Christensen H, Hansen M, Sørensen J (1999) Counting and size classification of active soil bacteria by fluorescence in situ hybridization with an rRNA oligonucleotide probe. Appl Environ Microbiol 65:1753–1761
Collins D, Luxton T, Kumar N, Shah S, Walker VK, Shah V (2012) Assessing the impact of copper and zinc oxide nanoparticles on soil: a field study. PLoS One 7:e42663. https://doi.org/10.1371/journal.pone.0042663
Cornelis G, Hund-Rinke K, Kuhlbusch T, Van den Brink N, Nickel C (2014) Bioavailability of engineered nanoparticles in soils: a review. Crit Rev Environ Sci Technol 44:2720–2764. https://doi.org/10.1080/10643389.2013.829767
Cox M, Lehninger AL, Nelson DR (2000) Lehninger principles of biochemistry. Worth Publishers, New York
Curtis TP, Sloan WT, Scannel JW (2002) Estimating prokaryotic diversity and its limits. Proc Natl Acad Sci U S A 99:10494–10499. https://doi.org/10.1073/pnas.142680199
Dalzell DJB, Alte S, Aspichueta E, de la Sota A, Etxebarria J, Gutierrez M, Hoffmann CC, Sales D, Obst U, Christofi N (2002) A comparison of five rapid direct toxicity assessment methods to determine toxicity of pollutants to activated sludge. Chemosphere 47:535–545. https://doi.org/10.1016/S0045-6535(01)00331-9
Dick WA, Tabatabai MA (1984) Kinetic parameters of phosphatases in soils and organic waste materials. Soil Sci 137:7–15. https://doi.org/10.1097/00010694-198401000-00002
Donner E, Broos K, Heemsbergen D, Warne MSJ, McLaughlin MJ, Hodson ME, Nortcliff S (2010) Biological and chemical assessments of zinc ageing in field soils. Environ Pollut 158:339–345. https://doi.org/10.1016/j.envpol.2009.06.034
Eivazi F, Tabatabai MA (1977) Phosphatases in soils. Soil Biol Biochem 9:167–172. https://doi.org/10.1016/0038-0717(77)90070-0
Farrah H, Pickering WF (1997) Influence of clay-solute interactions on aqueous heavy metal ion levels. Water Air Soil Pollut 8:189–197. https://doi.org/10.1007/BF002940422
Fernandez D, Garcia-Gomez C, Babin M (2013) In vitro evaluation of cellular responses induced by ZnO nanoparticles, zinc ions and bulk ZnO in fish cells. Sci Total Environ 452-453:262–274. https://doi.org/10.1016/j.scitotenv.2013.02.079
Fernandez-Calvino D, Rousk J, Brookes PC, Baath E (2011) Bacterial pH-optima for growth track soil pH, but are higher than expected at low pH. Soil Biol Biochem 43:1569–1575. https://doi.org/10.1016/j.soilbio.2011.04.007
Fierer N, Jackson RB (2006) The diversity and biogeography of soil bacterial communities. Proc Natl Acad Sci U S A 103:626–631. https://doi.org/10.1073/pnas.0507535103
Freeman C, Liska G, Ostle NJ, Jones SE, Lock MA (1995) The use of fluorogenic substrates for measuring enzyme-activity in peatlands. Plant Soil 175:147–152. https://doi.org/10.1007/BF02413020
Frenk S, Ben-Moshe T, Dror I, Berkowitz B, Minz D (2013) Effect of metal oxide nanoparticles on microbial community structure and function in two different soil types. PLoS One 8:e84441. https://doi.org/10.1371/journal.pone.0084441
Garcia-Gomez C, Babin M, Obrador A, Alvarez JM et al (2015) Integrating ecotoxicity and chemical approaches to compare the effects of ZnO nanoparticles, ZnO bulk, and ZnCl on plants and microorganisms in a natural soil. Environ Sci Pollut Res Int 22:16803–16813. https://doi.org/10.1007/s11356-015-4867-y
Garland J (1996) Analytical approaches to the characterization of samples of microbial communities using patterns of potential c source utilization. Soil Biol Biochem 28:213–221. https://doi.org/10.1016/0038-0717(95)00112-3
Garland JL, Mills AL (1991) Classification and characterization of heterotrophic microbial communities on the basis of patterns of community-level-sole-carbon-source-utilization. Appl Environ Microbiol 57:2351–2359
Ge Y, Schimel JP, Holden PA (2011) Evidence for negative effects of TiO2 and ZnO nanoparticles on soil bacterial communities. Environ Sci Technol 45:1659–1664. https://doi.org/10.1021/es103040t
Ge Y, Schimel JP, Holden PA (2012) Identification of soil bacteria susceptible to TiO2 and ZnO nanoparticles. Appl Environ Microbiol 78:6749–6758. https://doi.org/10.1128/AEM.00941-12
Giller KE, Witter E, McGrath SP (1998) Toxicity of heavy metals to microorganisms and microbial processes in agricultural soils: a review. Soil Biol Biochem 30:1389–1414. https://doi.org/10.1016/S0038-0717(97)00270-8
Heggelund LR, Diez-Ortiz M, Lofts S, Lahive E, Jurkschat K, Wojnarowicz J, Cedergreen N, Spurgeon D, Svendsen C (2014) Soil pH effects on the comparative toxicity of dissolved zinc, non-nano and nano ZnO to the earthworm Eisenia fetida. Nanotoxicology 8:559–572. https://doi.org/10.3109/17435390.2013.809808
Holden PA, Schimel JP, Godwin HA (2014) Five reasons to use bacteria when assessing manufactured nanomaterial environmental hazards and fates. Curr Opin Biotechnol 27:73–78. https://doi.org/10.1016/j.copbio.2013.11.008
Houba VJG, Temminghoff EJM, Gaikhorst GA, Vark W (2000) Soil analysis procedures using 0.01 M calcium chloride as extraction reagent. Commun Soil Sci Plant Anal 31:1299–1396. https://doi.org/10.1080/00103620009370514
Huang PM, Wang MK, Chiu CY (2005) Soil mineral–organic matter–microbe interactions: impacts on biogeochemical processes and biodiversity in soils. Pedobiologia 49:609–635. https://doi.org/10.1016/j.pedobi.2005.06.006
Hund-Rinke K, Simon M (2008) Bioavailability assessment of contaminants in soils via respiration and nitrification tests. Environ Pollut 153:468–475. https://doi.org/10.1016/j.envpol.2007.08.003
Jiang W, Mashayekhi H, Xing B (2009) Bacterial toxicity comparison between nano-and micro-scaled oxide particles. Environ Pollut 157:1619–1625. https://doi.org/10.1016/j.envpol.2008.12.025
Jimenez MP, Horra A, Pruzzo L, Palma MR (2002) Soil quality: a new index based on microbiological and biochemical parameters. Biol Fertil Soils 35:302–306. https://doi.org/10.1007/s00374-002-0450-z
Jones N, Ray B, Ranjit KT, Manna AC (2008) Antibacterial activity of ZnO nanoparticle suspensions on a broad spectrum of microorganisms. FEMS Microbiol Lett 279:71–76. https://doi.org/10.1111/j.1574-6968.2007.01012.x
Josko I, Oleszczu P, Futa B (2014) The effect of inorganic nanoparticles (ZnO, Cr2O3, CuO and Ni) and their bulk counterparts on enzyme activities in different soils. Geoderma 232-234:528–537. https://doi.org/10.1016/j.geoderma.2014.06.012
Kim S, Kim J, Lee I (2011) Effects of Zn and ZnO nanoparticles and Zn2+ on soil enzyme activity and bioaccumulation of Zn in Cucumis sativus. Chem Ecol 27:49–55. https://doi.org/10.1080/02757540.2010.529074
Kim S, Sin H, Lee S, Lee I (2013) Influence of metal oxide particles on soil enzyme activity and bioaccumulation of two plants. J Microbiol Biotechnol 23:1279–1286. https://doi.org/10.4014/jmb.1304.04084
Knight B, McGrath SP, Chaudri AM (1997) Biomass carbon measurements and substrate utilization patterns of microbial populations from soils amended with cadmium, copper, or zinc. Appl Environ Microbiol 63:39–43
Langdon KA, McLaughlin MJ, Kirby JK, Merrington G (2014) The effect of soil properties on the toxicity of silver to the soil nitrification process. Environ Toxicol Chem 33:1170–1178. https://doi.org/10.1002/etc.2543
Liao M, Xiao XM (2007) Effect of heavy metals on substrate utilization pattern, biomass, and activity of microbial communities in a reclaimed mining wasteland of red soil area. Ecotoxicol Environ Saf 66:217–223. https://doi.org/10.1016/j.ecoenv.2005.12.013
Menard A, Drobne D, Jemec A (2011) Ecotoxicity of nanosized TiO2. Review of in vivo data. Environ Pollut 159:677–684. https://doi.org/10.1016/j.envpol.2010.11.027
Mudunkotuwa IA, Pettibone JM, Grassian VH (2012) Environmental implications of nanoparticle aging in the processing and fate of copper-based nanomaterials. Environ Sci Technol 46:7001–7010. https://doi.org/10.1021/es203851d
Nannipieri P, Giagnoni L, Landi L, Renella G (2011) Role of phosphatase enzymes in soil. Phosphorus in action. Soil biology. Bünemann, Oberson, Frossard, Berlin. https://doi.org/10.1007/978-3-642-15271-9_9
Negi H, Agarwal T, Zaidi MGH, Goel R (2012) Comparative antibacterial efficacy of metal oxide nanoparticles against Gram negative bacteria. Ann Microbiol 62:765–772. https://doi.org/10.1007/s13213-011-0317-3
Organization for Economic Cooperation and Development (OECD) (2000a) Guidelines for testing of chemicals. Soil microorganisms, carbon transformation test. Test guideline N° 217. Paris, France. https://doi.org/10.1787/20745761
Organization for Economic Cooperation and Development (OECD) (2000b) Guidelines for testing of chemicals. Soil microorganisms, nitrogen transformation test. Test guideline N° 216. Paris, France. https://doi.org/10.1002/etc.2398
Peyrot C, Wilkinson KJ, Desrosiers M, Sauve S (2014) Effects of silver nanoparticles on soil enzyme activities with and without added organic matter. Environ Toxicol Chem 33:115–125. https://doi.org/10.1002/etc.2398
Piccinno F, Gottschalk F, Seeger S, Nowack B (2012) Industrial production quantities and uses of ten engineered nanomaterials in Europe and the world. J Nanopart Res 14:1109. https://doi.org/10.1007/s11051-012-1109-9
Rahmatpour S, Shirvani M, Mosaddeghi MR, Nourbakhsh F, Bazarganipour M (2017) Dose–response effects of silver nano- particles and silver nitrate on microbial and enzyme activities in calcareous soils. Geoderma 285:313–322. https://doi.org/10.1016/j.geoderma.2016.10.006
Read DS, Matzke M, Gweon HS, Newbold LK, Heggelund L, Ortiz MD, Lahive E, Spurgeon D, Svendsen C (2016) Soil pH effects on the interactions between dissolved zinc, non-nano- and nano-ZnO with soil bacterial communities. Environ Sci Pollut Res 23:4120–4128. https://doi.org/10.1007/s11356-015-4538-z
Roesch LF, Fulthorpe RR, Riva A, Casella G et al (2007) Pyrosequencing enumerates and contrasts soil microbial diversity. ISME J 1:283–290. https://doi.org/10.1002/etc.2398
Rogers JE, Li SW (1985) Effect of metals and other inorganic-ions on soil microbial activity: soil dehydrogenase assay as a simple toxicity test. Bull Environ Contam Toxicol 34:858–865. https://doi.org/10.1007/BF01609817
Rousk J, Baath E, Brookes PC, Lauber CL et al (2010) Soil bacterial and fungal communities across a pH gradient in an arable soil. ISME J 4:1340–1351. https://doi.org/10.1038/ismej.2010.58
Rousk J, Ackermann K, Curling SF, Jones DL (2012) Comparative toxicity of nanoparticulate CuO and ZnO to soil bacterial communities. PLoS One 7:e34197. https://doi.org/10.1371/journal.pone.0034197
Sauve S, Hendershot W, Allen HE (2000) Solid-solution partitioning of metals in contaminated soils: dependence on pH, total metal burden, and organic matter. Environ Sci Technol 34:1125–1131. https://doi.org/10.1021/es9907764
Schlich K, Hund-Rinke K (2015) Influence of soil properties on the effect of silver nanomaterials on microbial activity in five soils. Environ Pollut 196:321–330. https://doi.org/10.1016/j.envpol.2014.10.021
Schloter M, Dilly O, Munch JC (2003) Indicators for evaluating soil quality. Agric Ecosyst Environ 98:255–262. https://doi.org/10.1016/S0167-8809(03)00085-9
Simon-Deckers A, Loo S, Mayne-L’hermite M, Herlin-Boime N et al (2009) Size-, composition-and shape- dependent toxicological impact of metal oxide nanoparticles and carbon nanotubes toward bacteria. Environ Sci Technol 43:8423–8429. https://doi.org/10.1021/es9016975
Simonin M, Richaume A (2015) Impact of engineered nanoparticles on the activity, abundance, and diversity of soil microbial communities: a review. Environ Sci Pollut Res Int 22:13710–13723. https://doi.org/10.1007/s11356-015-4171-x
Simonin M, Richaume A, Guyonnet JP, Dubost A, Martins JMF, Pommier T (2016) Titanium dioxide nanoparticles strongly impact soil microbial function by affecting archaeal nitrifiers. Sci Rep 6:33643. https://doi.org/10.1038/srep33643
Simonin M, Martins JM, Le Roux X, Uzu G et al (2017) Toxicity of TiO2 nanoparticles on soil nitrification at environmentally relevant concentrations: lack of classical dose–response relationships. Nanotoxicology 11:247–255. https://doi.org/10.1080/17435390.2017.1290845
Smolders E, Buekers J, Oliver I, McLaughlin MJ (2004) Soil properties affecting toxicity of zinc to soil microbial properties in laboratory-spiked and field-contaminated soils. Environ Toxicol Chem 23:2633–2640. https://doi.org/10.1897/04-27
Speir TW, Kettles HA, Parshotam A, Searle PL, Vlaar LNC (1995) A simple kinetic approach to derive the ecological dose value, ED50, for the assessment of Cr(VI) toxicity to soil biological properties. Soil Biol Biochem 27:801–810. https://doi.org/10.1016/0038-0717(94)00231-O
Strachel R, Wyszkowska J, Bacmaga M (2017) The influence of nitrogen on the biological properties of soil contaminated with zinc. Bull Environ Contam Toxicol 98:426–432. https://doi.org/10.1007/s00128-016-1977-2
Tipping E, Rieuwerts J, Pan G, Ashmorec MR et al (2003) The solid–solution partitioning of heavy metals (Cu, Zn, Cd, Pb) in upland soils of England and Wales. Environ Pollut 125:213–225. https://doi.org/10.1016/S0269-7491(03)00058-7
Tourinho PS, van Gestel CA, Lofts S, Soares AM, Loureiro S (2013) Influence of soil pH on the toxicity of zinc oxide nanoparticles to the terrestrial isopod Porcellionides pruinosus. Environ Toxicol Chem 32:2808–2815. https://doi.org/10.1002/etc.2369
Voet D, Voet JG, Pratt CW (2006) Fundamentals of biochemistry: life at the molecular level. Wiley, New York
Waalewijn-Kool PL, Ortiz MD, Lofts S, van Gestel CAM (2013) The effect of pH on the toxicity of zinc oxide nanoparticles to Folsomia candida in amended field soil. Environ Toxicol Chem 32:2349–2355. https://doi.org/10.1002/etc.2302
Wang P, Menzies NW, Lombi E, McKenna BA et al (2013) Fate of ZnO nanoparticles in soils and cowpea (Vigna unguiculata). Environ Sci Technol 47:13822–13830. https://doi.org/10.1021/es403466p
Wang Q, Kang F, Gao Y, Mao X, Hu X (2016) Sequestration of nanoparticles by an EPS matrix reduces the particle-specific bactericidal activity. Sci Rep 6:21379–21389. https://doi.org/10.1038/srep21379
Yao H, He Z, Wilson MJ, Campbell CD (2000) Microbial biomass and community structure in a sequence of soils with increasing fertility and changing land use. Microb Ecol 40:223–237. https://doi.org/10.1007/s002480000053
Yu H, Si P, Shao W, Qiao X, Yang X, Gao D, Wang Z (2016) Response of enzyme activities and microbial communities to soil amendment with sugar alcohols. Microbiologyopen 5:604–615. https://doi.org/10.1002/mbo3.355
Acknowledgements
The authors are grateful to Carmen del Rio for the technical assistance provided.
Funding information
This work was financed by the Spanish projects RTA2013-00091-C02-01 and RTA2013-00091-C02-02.
Author information
Authors and Affiliations
Corresponding author
Additional information
Responsible editor: Robert Duran
Rights and permissions
About this article

Cite this article
García-Gómez, C., Fernández, M.D., García, S. et al. Soil pH effects on the toxicity of zinc oxide nanoparticles to soil microbial community. Environ Sci Pollut Res 25, 28140–28152 (2018). https://doi.org/10.1007/s11356-018-2833-1
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s11356-018-2833-1