
Abstract
Data regarding NaCl impact on halophyte plant species exposed to a polymetallic contamination remain scarce. Seedlings of the salt marsh species Kosteletzkya pentacarpos were simultaneously exposed to cadmium (10 μM) and zinc (100 μM) in the absence or presence of 50 mM NaCl. Heavy metal exposure reduced plant growth and increased Cd and Zn concentrations in all organs. Cd and Zn accumulation reduced net photosynthesis in relation to stomatal closure, decreased in chlorophyll concentration and alteration in chlorophyll fluorescence-related parameters. Salinity reduced Cd and Zn bioaccumulation and translocation, with a higher impact on Cd than Zn. It mitigated the deleterious impact of heavy metals on photosynthetic parameters. NaCl reduced the heavy metal-induced oxidative stress assessed by malondialdehyde, carbonyl, and H2O2 concentration. Subcellular distribution revealed that Cd mainly accumulated in the cell walls, but NaCl increased it in the cytosol fraction in the leaf and in the metal-rich granule fraction in the roots. It had no impact on Zn subcellular distribution. The additional NaCl contributed to a higher sequestration of Cd on phytochelatins and stimulated glutathione synthesis. The positive impact of NaCl on K. pentacarpos response to polymetallic pollution made this species a promising candidate for revegetation of heavy metal-contaminated salt areas.
Similar content being viewed by others

Explore related subjects
Discover the latest articles, news and stories from top researchers in related subjects.Avoid common mistakes on your manuscript.
Introduction
The increasing anthropogenic activities, such as mining, agriculture, metallurgy, combustion of fossil fuels, and military operations, have led to widespread contamination of the environment (Ogundele et al. 2017; Różański et al. 2017). In addition to organic contamination, heavy metals such as cadmium, zinc, lead, copper, mercury, and nickel constitute the most hazardous soil pollutants. Although some of them (Cu, Zn) are essential elements for living organisms, their presence in the soil at high concentrations are deleterious to plant growth and survival. Heavy metals impair plant photosynthesis, induce oxidative stress, affect plant mineral nutrition, and compromise the plant water and hormonal status (Tattibayeva et al. 2016; Wang et al. 2015; Shahid et al. 2014).
The majority of studies devoted to heavy metal impact on plant behavior consider one single heavy metal. However, in real metal-polluted environmental settings, several heavy metals often coexist which interact with each other in a complex way. This is especially the case for Cd and Zn, which are frequently simultaneously present on polluted soils (Mani et al. 2015). Although these elements share several physicochemical properties, they have quite different impact on plant physiology since Zn is an essential element while Cd has no recognized biological functions. Cadmium and zinc may have different distributions in heavy metal-treated plants (Lefèvre et al. 2014) and may bind to different complexing compounds (Lefèvre et al. 2016). However, their putative interaction in the case of polymetallic contamination remains poorly documented. It might be argued that cadmium and zinc interact, to some extent, not only in terms of absorption but also in terms of plant response to accumulated ions (Qiu et al. 2011). Moreover, the overall plant response is directly influenced by ion distribution and the presence of NaCl may somewhat affect both processes in a complex way (Lutts and Lefèvre 2015).
Heavy metal pollution is frequently reported in coastal areas, leading to imbalance of soil ecological system and serious threat to food safety (El Nemr and El-Said 2017). Hence, it is of crucial importance to study the behavior of plants simultaneously exposed to salinity and heavy metal conditions. Halophyte species have been considered as a promising material for phytoremediation of heavy metal-contaminated sites, and moderate doses of salt are thought to play a role in the protection against heavy metal toxicity in these species (Lutts and Lefèvre 2015). Once again, such a protective effect has often been tested on plants exposed to one single heavy metal stress (Nawaz et al. 2017; Zhang et al. 2016). For instance, salinity improved zinc tolerance by the halophyte Spartina densiflora in relation to maintenance of photosynthetic apparatus and mineral nutrition (Redondo-Gomez et al. 2011). Salinity also improved Cd tolerance in Sesuvium portulacastrum, which could be explained by an increase in glutathione (GSH), proline concentration, maintenance of the redox balance, and photosynthesis (Wali et al. 2015, 2016), although it has been recently demonstrated by Wali et al. (2017) that Cd hampered salt tolerance in this species.
Kosteletzkya pentacarpos (L.) Presl. (formerly designed as Kosteletzkya virginica) is a perennial dicot halophyte species of Malvaceae family, and it is recommended as a potential resource for food, feed, biodiesel, as well as health care (Halchak et al. 2011; Qin et al. 2015; Vaughn et al. 2013). Kosteletzkya pentacarpos is able to cope with a high level of salinity in its natural environment (up to 420 mM NaCl), exhibiting a high selectivity for K over Na (Blits and Gallagher 1990). Ghanem et al. (2010) demonstrated that mucilage produced by the plant may be involved in Na fixation in stem, preventing accumulation of this toxic element in photosynthetic leaves.
Kosteletzkya pentacarpos is also, to some extent, able to cope with heavy metal pollution in salt marsh conditions, and it could therefore be recommended as an interesting tool for phytomanagement of polluted coastal areas (Han et al. 2012). Han et al. (2013a) demonstrated that NaCl differently interfered with Cd and Zn toxicities in this wetland species. Cadmium increased the leaf K concentration while Zn had an opposite effect. Salinity reduced Cd accumulation to a higher extent than Zn accumulation. Distribution of heavy metals among plant organs also appeared differently affected by salinity since Cd was reduced mainly in the leaves while Zn was reduced in the roots. Management of heavy metal oxidative stress by K. pentacarpos appeared as a crucial component of resistance to Cd (Han et al. 2013b) or Zn (Han et al. 2013a). These data, however, were obtained for plants exposed to one single pollutant (Cd or Zn), but this species was never tested in the simultaneous presence of the two heavy metals.
The present work was undertaken in order to answer the following questions: (1) How does the addition of NaCl influence the growth response of K. pentacarpos simultaneously exposed to cadmium and zinc stresses? (2) What is the effect of NaCl on Cd and Zn absorption and distribution in the case of polymetallic contamination? (3) What is the impact of a mixed pollution on the management of antioxidative status by the plant and NaCl influence on this property?
Materials and methods
Plant material and culture condition
Seeds of Kosteletzkya pentacarpos were harvested from the Jinhai Agricultural Experimental Farm of Yancheng (Jiangsu Province) and kindly provided by Prof. P. Qin, University of Nanjing (People’s Republic of China). Germination was performed in trays filled with a perlite and vermiculite mix (1:3 v/v) and moistened regularly with a half-strength, modified Hoagland nutrient solution. Seedlings were grown in a phytotron under a 12-h photoperiod [mean light intensity (photosynthetically active radiation, PAR) = 150 μmol m−2 s−1 provided by Osram Sylvania (Danvers, MA) fluorescent tubes (F36W/133-T8/CW) with 25/23 °C day/night temperature and 70/50% atmospheric humidity]. Fifteen days after sowing, seedlings were fixed on polyvinylchloride plates floating on an aerated half-strength, modified Hoagland nutrient solution and transferred in 50-L tanks into a greenhouse. The nutrient solution contained the following chemicals (in mM): 2.0 KNO3, 1.7 Ca(NO3)2, 1.0 KH2PO4, 0.5 NH4NO3, and 0.5 MgSO4 and (in μM) 17.8 Na2SO4, 11.3 H3BO3, 1.6 MnSO4, 1 ZnSO4, 0.3 CuSO4, 0.03 (NH4)6Mo7O24, and 14.5 Fe-EDDHA. Minimum temperatures were 16–18 °C, and daily maxima were 24–28 °C. Natural light was supplemented by Philips lamps (Philips Lighting S.A., Brussels, Belgium) (HPLR 400 W) in order to maintain a light irradiance of 300 μmol m−2 s−1 (PAR) at the top of the canopy.
After 10 days of acclimation in the absence of stress (25 days after sowing), NaCl, CdCl2, and ZnCl2 were added to containers in order to create four treatments: (1) control, (2) 50 mM NaCl, (3) 10 μM CdCl2 + 100 μM ZnCl2 (heavy metals (HMs)), and (4) 10 μM CdCl2 + 100 μM ZnCl2 + 50 mM NaCl (HMs + Na). Solutions were readjusted every 2 days and renewed every week. The pH of solutions was set to 5.7 ± 0.02 with KOH. Three replications with 12 plants per replication and per treatment were used for the measurement of different parameters.
Growth assessment
After 2 weeks of treatment, stem height, the number of lateral branches (LBs), the number of leaves (LN) on the main stem, the number of leaves on LBs, and total length of LBs were recorded. Plants were then harvested. Roots were quickly rinsed in sterile deionized water for 30 s to remove ions from the free space and gently blotted dry with a paper towel. Roots, stems, and leaves of each plant were separated and weighed for fresh weight determination. Before harvest, total leaf area of each plant was measured with a leaf area meter (AM300 leaf area meter; ADC BioScientific, Ltd., Hoddesdon, UK). Material was then incubated in an oven for 72 h at 70 °C for dry weight determination.
Evaluation of ion concentration
Dried samples were ground to a fine powder using a porcelain mortar and a pestle, digested in 35% HNO3, and evaporated to dryness on a sand bath at 80 °C. The minerals were incubated with a mix of 37% HCl and 68% HNO3 (3:1), and the mixture was slightly evaporated. Minerals were dissolved in 0.1 N HCl. Ion concentrations were determined by Solaar S4 atomic absorption spectrometry (Thermo Scientific, Cambridge, UK). For each treatment, three separated plants were considered and each analysis was performed on technical triplicates.
Translocation factor (TF) is representative of the plant’s ability to translocate pollutant from the root to the shoot system. In the present study, it was estimated for Cd and Zn on the basis of concentration (TFc) and on the basis of the total amount (TFa) of translocated pollutants according to the following:
- TFc:
Concentration in the shoot (mg g−1 dry weight (DW)) / Concentration in the roots (mg g−1 DW)
- TFa:
Total amount in the shoot (mg) / Total amount in the roots (mg)
The bioaccumulation factor (BF) is an indicator of the plant’s ability to accumulate the heavy metal in harvestable organs compared to its mean concentration in the environment. Since a nutrient solution was used in the present experiment, we expressed shoot Cd and Zn concentration on a tissue water content basis for BF calculation according to the following:
- BF:
Shoot concentration of heavy metal (μM) / Concentration in the nutrient solution (μM)
Subcellular distribution of Cd and Zn
The subcellular distribution of Cd and Zn in K. pentacarpos roots and shoots was performed at 4 °C according to Weigel and Jager (1980) with some modification (Li et al. 2011): 0.2 g fresh material was ground with liquid nitrogen and homogenized in the 5.0 mL buffer solution (0.25 M sucrose, 1.0 mM dithioerythritol, and 50 mM Tris-HCl (pH 7.5)). The homogenate was centrifuged at 2500g for 20 min. The supernatant of this first step was regarded as a cytosol fraction. After adding 2.0 mL ultra-pure water, the pellet was heated at 100 °C for 2 min, followed by adding 2.0 mL NaOH (1 M) and heated again at 70 °C for 1 h. Samples were then centrifuged at 10,000g for 15 min. The supernatant and pellet were designated as a cell debris fraction (containing mainly cell walls) and metal-rich granule fraction (granules containing heavy metals bound to sulfur, iron, calcium, and carbonates), respectively. The three factions were separately analyzed for ion content by spectrometry as described above.
Photosynthesis-related parameters
Photosynthesis-related parameters were determined on leaves located at the middle portion of the main stem. A portable pulse-modulated chlorophyll fluorimeter (FMS2; Hansatech, King’s Lynn, UK) was used to determine the chlorophyll fluorescence. All measurements were performed in the middle part of the abaxial side of the leaves. Leaf portions were acclimated to darkness for 30 min. The minimal fluorescence level (F0) was measured by estimating the modulated light (0.1 μmol m−2 s−1). The maximal fluorescence level (Fm) with all photosystem II (PSII) reaction centers closed was determined by a 0.8-s saturating pulse at 18,000 μmol m−2 s−1 in dark-adapted leaves. Leaf was then continuously illuminated with white actinic light (600 mmol m−2 s−1) for 3 min. The steady-state value of fluorescence (Fs) was recorded, and a second saturating pulse at 18,000 μmol m−2 s−1 was imposed to determine maximal fluorescence level in the light-adapted state (F′m). The actinic light was removed, and the minimal fluorescence level in the light-adapted state (F′0) was determined by illuminating the leaf with a 3-s pulse of far-red. Using both light and dark fluorescence parameters, the maximal efficiency of PSII photochemistry in the dark-adapted state (Fv/Fm), the photochemical quenching coefficient (qP), the non-photochemical quenching (NPQ), and the actual PSII efficiency (ФPSII) were calculated according to Maxwell and Johnson (2000).
Chlorophyll (Chl a and Chl b) and total carotenoid (xanthophylls and β-carotene) concentrations were quantified on the whole leaves from seven plants per treatment after acetone extraction as previously described (Han et al. 2012). Net photosynthesis (A) was recorded with an infrared gas analyzer (LCA4 8.7; ADC BioScientific, Ltd., Hoddesdon, Hertfordshire, UK) using a Parkinson leaf cuvette (PLC) on intact leaves for 1 min (20 records min−1) with an air flow rate of 300 mL min−1. Air taken in the greenhouses was sent to a chamber into which a leaf portion of 3.5 cm2 was introduced. The net CO2 assimilation rate and instantaneous transpiration rate (E) were estimated on leaves located at the middle part of the main stem. Leaf stomatal conductance (gs) was measured using a diffusion porometer (AP4; Delta-T Devices, Ltd., Cambridge, UK). Five plants were measured for each treatment, and all measurements were performed around at midday (between 12:30 a.m. and 2:30 p.m.).
Oxidative stress parameters and non-enzymatic antioxidants
The level of lipid peroxidation was measured as 2-thiobarbituric acid-reactive substances, mainly malondialdehyde (MDA) (Heath and Packer 1968). The concentration of MDA was calculated using an extinction coefficient of 155 mM−1 cm−1. Carbonyl assay was performed using the Reznick and Packer (1994) spectrophotometric method in detecting the product of the reaction of dinitrophenylhydrazine (DNPH) with protein carbonyls to form protein hydrazones.
For hydrogen peroxide quantification, samples (0.5 g fresh weight (FW)) were ground to powder in the presence of 5 mL of 5% TCA. The mixture was centrifuged at 10,000g for 20 min at 4 °C. The supernatant was adjusted to pH 8.4 with 17 M ammonia solution and then filtered. The filtrate was divided into aliquots of 1 mL. To one of these (the blank), 8 μg of catalase (CAT) (10,000 U mg−1) was added and samples were then kept at room temperatures for 10 min. To both aliquots (with and without CAT), 1 mL of colorimetric reagent was added. The reaction solution was incubated for 10 min at 30 °C. Absorbance at 505 nm was determined. The colorimetric reagent contained 10 mg of 4-aminoantipyrine, 10 mg of phenol, and 5 mg of peroxidase (150 U mg−1) dissolved in 50 ml of 100 mM acetic buffer (pH 5.6) (Zhou et al. 2006).
For ascorbate extraction, frozen tissues were homogenized in ice-cold 5% metaphosphoric acid solution (1:5, w/v) and then centrifuged at 20,000g and 4 °C for 10 min. Total ascorbate (AsA) + dehydroascorbate (DHA) contents were determined according to Wang et al. (1991) on the basis of Fe3+-Fe2+ reduction by ascorbate in acid solution. Fe2+ forms a red chelate with bathophenanthroline absorbing at 534 nm. The ascorbate (reduced form) assay mixture contained 0.1 mL of the extract, 0.5 mL of absolute ethanol, 0.6 M trichloroacetic acid, 3 mM bathophenanthroline, 8 mM H3PO4, and 0.17 mM FeCl3. The final total volume was 1.5 mL, and the mixture was allowed to stand at 30 °C for 90 min. The absorbance of the colored solution was read at 534 nm. The total ascorbate assay mixture contained 0.1 mL of the sample, 0.15 mL of 3.89 mM dithiothreitol and 0.35 mL of absolute ethanol in a total volume of 0.6 mL. Then, the reaction mixture was left standing at room temperature for 10 min. After reduction of dehydroascorbate to ascorbate, 0.15 mL of 20% trichloroacetic acid was added and the color was developed by adding 0.15 mL of 0.4% (v/v) H3PO4-ethanol, 0.3 mL of 0.5% (w/v) bathophenanthroline-ethanol, and 0.15 mL of 0.03% (w/v) FeCl3-ethanol. DHA concentrations were estimated from the difference of total ascorbate and ascorbate concentration. Standard curve in the range 0–10 μmol ascorbate was used.
For reduced GSH and total glutathione (GSHt) quantification, 200 mg of frozen samples were extracted and derivatized by ortho-phthalaldehyde according to Cereser et al. (2001). GSHt was quantified after a reduction step of oxidized glutathione (GSSG) by dithiothreitol. Extracts were filtered through 0.45-μm microfilters (Chromafil PES-45/15, Macherey-Nagel) prior to injection, and OPA derivatives were separated on a reversed-phase HPLC column with an acetonitrile-sodium acetate gradient system and detected fluorimetrically. Five microliters of the sample was injected into a Shimadzu HPLC system (Shimadzu, ‘s-Hertogenbosch, The Netherlands) equipped with a Nucleodur C18 Pyramid column (125 × 4.6 mm internal diameter; 5 μm particle size) (Macherey-Nagel, Düren, Germany). Derivatives were eluted in acetonitrile gradient in a 50 mM sodium acetate buffer (pH 6.2) at 30 °C at a flow rate of 0.7 mL min−1. Fluorimetric detection was performed with a spectrum system Shimadzu RF-20A fluorescence detector at 420 nm after excitation at 340 nm. GSH was quantified using nine-point calibration curves with custom-made external standard solutions ranging from 0.0625 to 50 μM, and in every ten injections, a check standard solution was used to confirm the calibration of the system. The recovery was determined using GSH as an internal standard.
To estimate the total global antioxidant activity, ferric reducing ability of plasma (FRAP) was assayed according to Benzie and Strain (1996), considering the ability of plant extract to reduce ferric to ferrous ion at low pH and to produce a colored ferrous-tripyridyltriazine complex which was spectrophotometrically detected at 593 nm. A second assay was performed using the 2,2′-azinobis(3-ethylbenzothiazoline-6-sulfonic acid) (ABTS) decolorization procedure according to Pellegrini et al. (1999) assays. Results are expressed in μM Trolox equivalents (TE)/g fresh mass.
Evaluation of total non-protein thiol and phytochelatin content
The total non-protein thiol (NPT) concentration was determined according to De Vos et al. (1992): 200 mg fresh weight of tissue was ground in 2 mL of 5% (w/v) sulfosalicylic acid plus 6.3 mM diethylenetriaminepentaacetic acid (pH < 1) at 0 °C with quartz sand in a mortar. The homogenate was centrifuged at 10,000g for 10 min at 4 °C. The supernatants were collected and used for the determination of thiols using Ellman’s reagent. Three hundred microliters of the supernatant was mixed with 630 μL of 0.5 M KH2PO4 and 25 μL of 10 mM 5,5-dithiobis(2-nitrobenzoic acid) (final pH 7.0). The absorbance at 412 nm was recorded after 2 min, and the NPT concentration was estimated using an extinction coefficient of 13,600 M−1 cm−1. Phytochelatin content was evaluated as the difference between NPT and GSH levels (Schäfer et al. 1997).
Statistical analysis
For each treatment, three 50-L tanks containing 12 plants each were used in a randomized complete block design. Tissue material from five individual plants were dried in a 70 °C oven, which were used to analyze for growth, water status, and ionic determination, and seven remaining individual plants were frozen in liquid nitrogen immediately and analyzed for pigment content, HM subcellular distribution, MDA, carbonyl, H2O2, antioxidant (GSH, GSSH, AsA, and DHA), NPT, phytochelatins (PCs), as well as FRAP and ABTS. Each analysis was performed on technical triplicates. All of parameter data were subjected to an analysis of variance, one-way ANOVA, using SPSS software, with the treatment considered as the main factor. The statistical significance of the results was analyzed by Tukey’s test at 5% level (P < 0.05).
Results
Plant growth and water status
All plants remained alive until the end of the experiment. However, after 5–7 days of treatment, chlorosis and necrosis were observed on HM-treated plants only. Salinity had no significant impact on morphological properties (Table 1). In contrast, HMs in the absence of NaCl reduced the LN on the main stem, the number of LBs, and the number of leaves on lateral branches. The main stem length, the total branch length, and the total leaf area were also clearly affected by HM treatment. The addition of NaCl partly alleviated the toxicity of HMs on all morphological parameters, except on LN.
Heavy metals had a detrimental impact on root and leaf dry weight (Fig. 1) which were indeed reduced by 63 and 44%, respectively. The addition of NaCl significantly improved the root dry weight by 51%, when compared to HMs alone, but it had no impact on the leaf dry weight. As shown in Fig. 1c, d, salinity did not affect water content in roots and leaves. In contrast, HMs led to significant water loss. The presence of NaCl was, however, unable to improve this parameter.

a Root and b leaf dry weight and c root and d leaf water content in the seedlings of Kosteletzkya pentacarpos exposed during 2 weeks to heavy metals (HMs; 10 μM CdCl2 + 100 μM ZnCl2) in the presence or in the absence of 50 mM NaCl. Each value is the mean of five replicates, and vertical bars are SE. Values exhibiting different letters are significantly different at P < 0.05 according to Tukey’s test
Accumulation and subcellular distribution of Cd and Zn
As expected, Cd was not detected in control or NaCl-treated plants. After 2 weeks of HM treatment, the total concentration of Cd was higher in the roots than in the leaves (Table 2). The presence of 50 mM NaCl, however, reduced Cd concentration in all organs but to a higher extent in the leaves (74%) than in the roots (37%). In roots and leaves, the majority of accumulated Cd was detected in the cell debris fraction while less than 25% was present in the cytoplasm. NaCl had a contrasting impact on Cd accumulation in the cytoplasm depending on the considered organ since it decreased it in the roots while it increased it in the leaves. The presence of 50 mM NaCl increased Cd recorded in the root metal-rich granule (MRG) fraction and in the leaf cell debris fraction.
The additional NaCl did not affect Zn concentration in plants exposed to non-contaminated solution. In the presence of heavy metals, zinc accumulated to a higher extent in the roots than in the leaves. NaCl only slightly reduced Zn concentration in the roots (12%) while it reduced Zn accumulation in the shoots by more than 44%. In the roots of control plants, Zn was present in similar proportion in the cytoplasm and cell debris fraction and only a minor proportion was recorded in the root MRG fractions. In contrast, MRG fraction contained more or less 20% of leaf Zn and the presence of this element in the cytoplasm was clearly lower than that in roots. It is noteworthy that NaCl had no obvious impact on Zn distribution in roots and leaves.
NaCl also reduced Cd and Zn accumulation in the stems (from 318 to 109 μg g−1 DW for Cd in HM and HM + NaCl-treated plants and from 841 to 388 μg g−1 DW for Zn; detailed data not shown). The bioaccumulation factor recorded in HM-treated plants (Table 3) was higher for Cd than for Zn; in both cases, the presence of NaCl reduced BF values, but its impact was higher for Cd than for Zn. For both elements, translocation factor estimated on a total amount basis (TFa) was higher than translocation factor estimated on a concentration basis (TFc). As far as Zn is concerned, TFa was slightly higher in HM-treated plants than in controls. NaCl reduced Cd TFc by 47% and TFa by more than 60%. In contrast, NaCl had no significant impact on the Zn TFc values, although it reduced TFa by 50%.
Sodium concentration increased from 0.52 ± 0.03 to 12.9 ± 2.0 mg g−1 DW in the roots of control and NaCl-treated plants, respectively. The presence of heavy metals in the solution significantly decreased the root Na concentration to 5.6 ± 0.04 in HM + Na-treated plants. Heavy metals in the solution, however, did not reduce the leaf Na which was 13.2 ± 2.9 mg g−1 DW in NaCl-treated plants and 14.0 ± 0.2 mg g−1 DW in HM + NaCl-treated ones (detailed data not shown).
Photosynthesis-related parameters
In the absence of HM, the addition of NaCl had no impact on net CO2 assimilation rate, instantaneous transpiration, and stomatal conductance (Fig. 2a–c). However, after 2 weeks of exposure to heavy metal stress, A, E, and gs values decreased sharply. The addition of NaCl to HM treatment significantly mitigated the deleterious effects of pollutant on these parameters.

Net photosynthesis (A; a), instantaneous transpiration (E; b), stomatal conductance (gs; c), maximal efficiency of PSII photochemistry in the dark-adapted state (Fv/Fm; d), photochemical quenching (qP; e), non-photochemical quenching (NPQ; f), actual PSII efficiency (ФPSII; g), total chlorophyll (Chl a + Chl b; h), and total carotenoid (i) in the seedlings of Kosteletzkya pentacarpos exposed during 2 weeks to heavy metals (HMs; 10 μM CdCl2 + 100 μM ZnCl2) in the presence or in the absence of 50 mM NaCl. Value of pigment content is the mean of three replicates, and others are the mean of five replicates. Vertical bars are SE. Values exhibiting different letters are significantly different at P < 0.05 according to Tukey’s test
Heavy metals also had a direct impact on fluorescence-related parameters. The maximal efficiency of PSII photochemistry in the dark-adapted state (Fv/Fm) was significantly reduced when K. pentacarpos was exposed to HMs (Fig. 2d), and a similar observation was made for qP (Fig. 2e) and ФPSII (Fig. 2g). The presence of NaCl in HM-contaminated solutions slightly increased Fv/Fm, qP, and ФPSII comparatively to values recorded in the absence of salt. The presence of NaCl in the absence of HM slightly increased NPQ values (Fig. 2f). The recorded increase in NPQ was, however, higher in plants exposed to HM, and the addition of NaCl to polluted solution had no impact on this parameter.
Plants exposed to 50 mM NaCl had a higher concentration of total chlorophyll and total carotenoid comparatively to controls (Fig. 2h, i). In contrast, HMs induced a sharp decrease in total chlorophyll and carotenoid concentration. The addition of NaCl to HM-containing solution reduced the negative impact of Cd and Zn on these photosynthetic pigment concentrations, which, however, remained lower than those in controls.
Oxidative stress parameters and non-enzymatic antioxidants
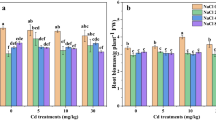
As indicated in Fig. 3, the additional NaCl alone did not induce MDA or carbonyl increase neither in the roots (Fig. 3a, b) nor in the leaves (Fig. 3d, e), suggesting that no oxidative stress occurred in response to NaCl. In contrast, both parameters increased in HM-treated plants and such an increase was associated to an increase in H2O2 in roots (Fig. 3c) and leaves (Fig. 3f). The addition of NaCl to contaminated solution reduced MDA, carbonyl, and H2O2 in roots as well as carbonyl and H2O2 in leaves. The additional NaCl alone had no impact on the total antioxidant activity estimated by FRAP (Fig. 4a, b) or ABTS (Fig. 4c, d) while HM strongly increased this total antioxidant activity. In all cases, the total antioxidant activity of HM-treated plants was reduced by NaCl addition.

Concentration of malondialdehyde (root, a; leaf, d), carbonyl (root, b; leaf, e), and hydrogen peroxide (root, c; leaf, f) in the seedlings of Kosteletzkya pentacarpos exposed during 2 weeks to heavy metals (HMs; 10 μM CdCl2 + 100 μM ZnCl2) in the presence or in the absence of 50 mM NaCl. Each value is the mean of three replicates, and vertical bars are SE. Values exhibiting different letters are significantly different at P < 0.05 according to Tukey’s test

Total antioxidant (estimated in terms of Trolox equivalent per g fresh weight) quantified by FRAP (Fig. 5a, b) and ABTS (Fig. 5c, d) methods in the seedlings of Kosteletzkya pentacarpos exposed during 2 weeks to heavy metals (HMs; 10 μM CdCl2 + 100 μM ZnCl2) in the presence or in the absence of 50 mM NaCl. Values are given separately for roots (a, c) and leaves (b, d). Each value is the mean of three replicates, and vertical bars are SE. Values exhibiting different letters are significantly different at P < 0.05 according to Tukey’s test
In roots, AsA and DHA, on the one hand (Fig. 5a, c), and GSH and GSSG, on the other hand (Fig. 5e, g), were strongly increased by HM treatment. NaCl added to HM-containing solution did not affect root AsA concentration but reduced root DHA to values similar to control plants. It also reduced GSH and GSSG, but in this case, values remained higher than those in control or NaCl-treated plants. As far as leaves were concerned, HM also increased AsA and DHA values (Fig. 5b, d) and the addition of NaCl mitigated such a HM-induced increase. NaCl significantly increased the leaf GSH concentration (Fig. 5f) while HM treatment reduced it. The mixed treatment (NaCl + HM) increased leaf GSH concentration to values similar to those recorded for NaCl-treated plants. The leaf GSSG concentration remained unaffected by the treatments (Fig. 5h).

Concentration of ascorbic acid (AsA; root, a; leaf, b), dehydroascorbate (DHA; root, c; leaf, d), reduced glutathione (GSH; root, e; leaf, f), and oxidized glutathione (GSSG; root, g; leaf, h) in the seedlings of Kosteletzkya pentacarpos exposed during 2 weeks to heavy metals (HMs; 10 μM CdCl2 + 100 μM ZnCl2) in the presence or in the absence of 50 mM NaCl. Each value is the mean of three replicates, and vertical bars are SE. Values exhibiting different letters are significantly different at P < 0.05 according to Tukey’s test
Evaluation of total NPT and PC content
Non-protein thiols were lower in the roots than in the leaves (Table 4), whatever the considered treatment. NaCl in the absence of heavy metals had no impact on the NPT concentrations. Exposure to HMs induced an increase in NPT values. PC constituted 90 and 92% of NPT in roots and leaves, respectively. It is noteworthy that in K. pentacarpos exposed to Cd + Zn solutions, PC concentration was higher in the leaves than in the roots. The addition of NaCl to HM-containing solution reduced both NPT and PC values.
Discussion
In coastal areas, heavy metal excess is often associated with high levels of salinity and contaminated areas are frequently characterized by multiple pollution. Zinc commonly contains 0.5–5% of Cd, and both elements are simultaneously present in polluted places. Although salinity impact on heavy metal absorption by plants has been studied in the case of monometallic exposure (Han et al. 2013a, 2013b; Wali et al. 2015; Ghabriche et al. 2017), information regarding salt impacts on plants facing a polymetallic situation remains scanty.
The present study demonstrates that 50 mM NaCl clearly mitigated the deleterious impact of Cd + Zn on the halophyte Kosteletzkya pentacarpos. Heavy metals indeed affected all morphological parameters. Han et al. (2012) reported that Cd clearly impacted plant ramification in this species while Zn did not. In the present study, a drastic effect on lateral branches growth was observed, suggesting that the presence of Zn in excess did not reduce Cd toxicity. Hormone synthesis and distribution within organs may be modified by heavy metals and have a strong impact on plant architecture (Li et al. 2009; Yan et al. 2016; Sofo et al. 2017). Han et al. (2013b) reported that in K. virginica, salinity reduced the synthesis of 1-aminocyclopropane-1-carboxylic acid (precursor of ethylene) and abscisic acid, which are both acting as senescing hormones. We may thus hypothesize that such a salt-induced modification may also help improve the growth of plants exposed to a mixed toxicity and delayed senescence may explain a higher concentration in photosynthetic pigments.
NaCl directly interferes with heavy metal absorption. Cadmium concentration was clearly lower in roots and leaves of plants in the presence of NaCl than in plants exposed to heavy metals in the absence of NaCl. From a relative point of view, the recorded decrease was higher for Cd than for Zn. Such a decrease could not be attributed to a decrease in transpiration rate since E and gs both increased in salt-treated plants exposed to heavy metals. Competition may occur between Cd and Zn, and the interactive pattern could be an antagonist in some cases and an additive or synergistic in other cases depending on the tested species, the external doses, and the environmental conditions (Sun et al. 2005; Qiu et al. 2011; Cherif et al. 2012; Tkalec et al. 2014). All these studies, however, focused on Cd and Zn interaction in the absence of salt. In our study performed under salt conditions, it is noteworthy that the recorded NaCl-induced decrease in Cd accumulation was higher than the salt-induced decrease in Cd accumulation when plants were, under similar experimental conditions, exposed to Cd alone as we previously reported (Han et al. 2012, 2013b): NaCl reduced leaf Cd concentration by 74% when Zn excess was simultaneously present, while in the case of exposure to Cd alone, an NaCl-induced decrease in leaf Cd was not higher than 44% (Han et al. 2012).
The overall consequence of these processes was that the bioaccumulation factor was reduced for both Cd and Zn in the presence of NaCl. Hence, if plants are used for phytoextraction purposes aiming at removing Cd and Zn from contaminated soils, salinity does not appear as a positive environmental factor. Some new strategies to recover and recycle Cd and Zn from harvested biomass start to be available but are more efficient when a small amount of highly contaminated biomass has to be manipulated (Hazotte et al. 2017). In our study, NaCl reduced the total amount of Cd and Zn in the shoot by 69.3 and 43.1%, respectively, confirming that NaCl-induced growth stimulation did not compensate the salt-induced decrease in heavy metal accumulation for pollutant removal.
Heavy metal translocation from roots to shoots was also strongly inhibited by NaCl. As a consequence, the total amount of heavy metals retained by the roots increased in NaCl-treated plants and to a higher extent for Zn (78%) than for Cd (25%). It has been recently demonstrated that NaCl increased the heavy metal (especially Zn) biosorption efficiency of roots from K. pentacarpos in relation to a salt-induced increase in mucilage pectic compounds and hemicellulose involved in heavy metal binding (Lutts et al. 2016). Although the mucilage in K. pentacarpos is mainly present in intercellular spaces (Ghanem et al. 2010) and hemicellulose is a major component of the cell wall, our data suggested that NaCl did not strongly increase the proportion of root Cd sequestered in cell walls which are recovered in the cell debris fraction. At the leaf level, Cd proportion in the cell wall was even lower in NaCl-treated plants than in those facing pollution in the absence of salt.
Beside cell wall sequestration, heavy metal complexation to cysteine-rich phytochelatin contributes to heavy metal resistance, especially for Cd (Sun et al. 2005; Lefèvre et al. 2016). In our study, heavy metal exposure increased PC concentration in leaves and roots, confirming that K. pentacarpos is able to trigger this protective mechanism. The addition of NaCl to a polluted medium reduced PC concentration, but the recorded decrease (20 and 50% in roots and leaves, respectively) was lower than the recorded decrease in Cd (38 and 73% in roots and leaves, respectively) which leads us to hypothesize that the proportion of Cd sequestered in PC increased in NaCl-treated plants. The Cd-PC complex is considered to accumulate in the vacuole where low molecular weight PCs polymerize to form high molecular weight insoluble complexes which are thought to be detected in the MRG fraction (Adams et al. 1997). This might explain that Cd proportion in MRG increased by 300% in our analyzed root sample.
Protection against oxidative stress is also an important component of heavy metal resistance in plants (Tkalec et al. 2014; Shahid et al. 2014; Yan et al. 2016). In the absence of heavy metals, no oxidative stress was recorded in salt-treated K. pentacarpos, confirming the halophytic nature of this species. This is also supported by the fact that Na was quite efficiently translocated to the shoots. NaCl was able to reduce MDA, carbonyl, and H2O2 in HM-treated plants. Chloroplasts are important sites for reactive oxygen species (ROS) synthesis, and Cherif et al. (2012) demonstrated that high Zn concentration exacerbates the negative effect of Cd on chloroplast structure. In our work, NaCl improved all fluorescence-related parameters (except NPQ), suggesting that salt contributes to protect the chloroplast structure. This protecting effect, together with the NaCl-induced increase in stomatal conductance and pigment concentration in HM-treated plants, probably contributed to the salt-induced improvement of net photosynthesis in plants challenging with Cd + Zn.
The total antioxidant activity slightly decreased in plants exposed to Cd + Zn in the presence of NaCl, suggesting that ROS synthesis rather than ROS detoxification was improved in the presence of salt. Glutathione, which is an important antioxidant, was, however, increased by NaCl but was depleted in HM-treated plants in the absence of salt. GSH assumes a dual function in the presence of heavy metals since it acts both as an antioxidant and as a precursor of PC (Schäfer et al. 1997). Hence, the recorded GSH depletion recorded in HM-treated plants could be attributed to the stimulation of these protective compounds, as previously reported in other species (De Vos et al. 1992; Sun et al. 2005). Our data, however, suggest that NaCl was able to restore GSH content which could be due both to a direct stimulation of its synthesis and to a lower requirement of this precursor for PC synthesis, considering the NaCl-induced decrease in HM accumulation.
Conclusion
These results demonstrate that 50 mM NaCl improved the ability of the halophyte plant species Kosteletzkya pentacarpos to cope with the simultaneous presence of Cd (10 μM) and Zn (100 μM). It increased plant growth in relation to a decrease of Cd and Zn absorption. The recorded decrease was higher for Cd than for Zn. NaCl also reduced heavy metal translocation to the shoot, which, in turn, allows to maintain photosynthetic activities. Exogenous NaCl increased GSH content and improved the antioxidative status of plants exposed to heavy metals. It compromises the use of K. pentacarpos for phytoextraction and heavy metal removal from contaminated soils, but its positive impact on plant growth could favor the use of this plant for revegetation of HM-polluted sites in salt marsh and coastal areas.
References
Adams SM, Shorey CD, Byrne M (1997) An ultrastructural and microanalytical study of metal-ionn content in granular concretions of the freshwater mussel Hydriella depressa. Micron 28:1–11
Benzie IF, Strain JJ (1996) The ferric reducing ability of plasma (FRAP) as a measure of “antioxidant power”: the FRAP assay. Anal Biochem 239:70–76
Blits KC, Gallagher JL (1990) Salinity tolerance of Kosteletzkya virginica 1. Shoot growth, ion and water relations. Plant Cell Environ 13:409–418
Cereser C, Guichard J, Drai J, Bannier E, Garcia I, Boget S, Parvaz P, Revol A (2001) Quantitation of reduced and total glutathione at the femtomole level by high-performance liquid chromatography with fluorescence detection: application to red blood cells and cultured fibroblasts. J Chromatogr B Biomed Sci Appl 752:123–132
Cherif J, Derbel N, Nakkach M, vopn Bergmann H, Jemal F, Ben Lakhdar Z (2012) Spectroscopic studes of photosynthetic responses of tomato plants to the interaction of zinc and cadmium toxicity. J Photochem Photobiol B Biol 111:9–16
De Vos CR, Vonk MJ, Vooijs R, Schat H (1992) Glutathione depletion due to copper-induced phytochelatin synthesis causes oxidative stress in Silene cucubalus. Plant Physiol 98:853–858
El Nemr A, El-Said GF (2017) Assessment and ecological risk of heavy metals in sediment and molluscs from the Mediterranean coast. Water Environ Res 89:195–210
Ghabriche R, Ghanya T, Mnasri M, Zaier H, Baioui R, Vromman D, Abdelly C, Lutts S (2017) Polyamine and tyramine involvement in NaCl-induced improvement of Cd resistance in the halophyte Inula chritmoides L. J Plant Physiol 6:136–144
Ghanem ME, Han RM, Classen B, Leclerq J, Mahy G, Ruan CJ, Pérez-Alfcoea F, Lutts S (2010) Mucilages and polysaccharides in the halophyte plant species Kosteletzkya virginica (L.) Presl.: localization and composition in relation to salt-induced physiological modifications. J Plant Physiol 167:382–392
Halchak JL, Seliskar DM, Gallagher JL (2011) Root system architecture of Kosteletzkya pentacarpos (Malvaceae) and belowground environmental influences on root and aerial growth dynamics. Am J Bot 98:163–174
Han RM, Lefèvre I, Ruan CJ, Qin P, Lutts S (2012) NaCl differently interferes with Cd and Zn toxicities in the wetland halophyte species Kosteletzkya virginica (L.) Presl. Plant Growth Regul 68:97–109
Han RM, Quinet M, André E, Van Elteren J, Destrebecq F, Vogel-Mikus K, Cui G, Debeljak M, Lefèvre I, Lutts S (2013a) Accumulation and distribution of Zn in the shoots and reproductive structures of the halophyte plant species Kosteletzkya virginica as a function of salinity. Planta 238:441–457
Han RM, Lefèvre I, Albacete A, Pérez-Alfocéa F, Baeba-Espín G, Díaz-Vivancos P, Quinet M, Ruan CJ, Hernández JA, Cantero-Navarro E, Lutts S (2013b) Antioxidant enzyme activities and hormonal status in response to Cd stress in the wetland halophyte Kosteletzkya virginica under saline conditions. Physiol Plant 147:352–368
Hazotte C, L ubie B, Rees F, Morel JL, Simonnot MO (2017) A novel process to recover cadmium and zinc from the hyperaccumulator plant Noccaea caerulescens. Hydrometallurgy 174:56–65
Heath RL, Packer L (1968) Photoperoxidation in isolated chloroplasts. II. Role of electron transfer. Arch Biochem Biophys 125:850–857
Lefèvre I, Vogel-Mikuš K, Jeromel J, Vavpetič P, Planchon s AI, Van Elteren J, Lepoint G, Gobert S, Renaut J, Pelicon P, Lutts S (2014) Cadmium and zinc distribution in relation to their physiological impact in the leaves of the accumulating Zygophyllum fabago L. Plant Cell Environ 37:1299–1320
Lefèvre I, Vogel-Mikuš K, Arčon I, Lutts S (2016) How do roots of the metal resistant perenial bush Zygophyllum fabago cope with cadmium and zinc toxicities? Plant Soil 404:193–207
Li T, Yang X, Lu L, Islma E, He Z (2009) Effects of zinc and cadmium interactions on root morphology and metal translocation in a hyperaccumulating species under hydroponic conditions. J Hazard Mater 169:734–741
Li D, Zhou D, Wang P, Li L (2011) Temperature affects cadmium-induced phytotoxicity involved in subcellular cadmium distribution and oxidative stress in wheat roots. Ecotoxicol Environ Saf 74:2029–2035
Lutts S, Lefèvre I (2015) How can we take advantage of halophyte properties to cope with heavy metal toxicity in salt-affected areas? Ann Bot 115:509–528
Lutts S, Qin P, Han RM (2016) Salinity influences biosorption of heavy metals by the root of the halophyte species Kosteletzkya pentacarpos. Ecol Eng 95:682–689
Mani D, Kumar C, Patel NK (2015) Integrated micro-biochemical approach for phytoremediation of cadmium and zinc contaminated soils. Ecotoxicol Environ Saf 111:86–95
Maxwell K, Johnson GN (2000) Chlorophyll fluorescence—a practical guide. J Exp Bot 51:659–668
Nawaz I, Iqbal M, Bliek M, Schat H (2017) Salt and heavy metal tolerance and expression levels of candidate tolerance genes among four extremophile Cochlearia species with contrasting habitat preferences. Sci Total Environ 584-585:731–741
Ogundele LT, Owoade OK, Hopke PK, Olise FS (2017) Heavy metals in industrially emitted particulate matter in Ile-Ife, Nigeria. Environ Res 156:320–325
Pellegrini N, Re R, Yang M, Rice-Evans CA (1999) Screening of dietary carotenoids and carotenoid-rich fruit extracts for antioxidant activities applying the 2,2′-azobis(3-ethylenebenzothiazoline-6-sulfonic) acid radical cation decolorization assay. Methods Enzymol 299:379–389
Qin P, Han RM, Zhou MX, Zhang HS, Fan LS, Seliskar DM, Gallagher JL (2015) Ecological engineering through the biosecure introduction of Kosteletzkya virginica (seashore mallow) to saline lands in China: a review of 20 years of activity. Ecol Eng 74:174–186
Qiu RL, Thangavel P, Hu PJ, Senthilkumar P, Ying RR, Tang YT (2011) Interaction of cadmium and zinc on accumulation and sub-cellular distribution in leaves of hyperaccumulator Potentilla griffithii. J Hazard Mater 186:1425–1430
Redondo-Gomez S, Andrades-Moreno L, Mateos-Naranjo E, Parra R, Valera-Burgos J, Aroca R (2011) Synergic effect of salinity and zinc stress on growth and photosynthetic responses of the cordgrass, Spartina densiflora. J Exp Bot 62:5521–5530
Reznick AZ, Packer L (1994) Oxidative damage to proteins: spectrophotometric method for carbonyl assay. Methods Enzymol 233:357–363
Różański S, Jaworska H, Matuszczak K, Nowak J, Hardy A (2017) Impact of highway traffic and the acoustic screen on the content and spatial distribution of heavy metals in soils. Environ Sci Pollut Res 24:12778–12786
Schäfer HJ, Greiner S, Rausch T, Haag-Kerwer A (1997) In seedlings of the heavy metal accumulator Brassica juncea Cu2+ differentially affects transcript amounts for γ-glutamylcysteine synthetase (γ-ECS) and metallothionein (MT2). FEBS Lett 404:216–220
Shahid M, Pourrut B, Dumat C, Nadeem M, Aslam M, Pinelli E (2014) Heavy-metal-induced reactive oxygen species: phytotoxicity and physicochemical changes in plants. Rev Environ Contam Toxicol 232:1–44
Sofo A, Bochicchio R, Amato M, Rendina N, Vitti A, Nuzzaci M, Altamura MM, Falasca G, Della Rovere F, Scopa A (2017) Plant architecture, auxin homeostasis and phenol content in Arabidopsis thaliana grown in cadmium- and zinc-enriched media. J Plant Physiol 216:174–180
Sun Q, Wang XR, Ding SM, Yuan XF (2005) Effects of interactions between cadmium and zinc on phytochelatin and glutathione production in wheat (Triticum aesticum L.). Environ Toxicol 20:195–201
Tattibayeva D, Nebot C, Miranda JM, Abuova AB, Baibatyrov TA, Kizatova MZ, Cepeda A, Franco CM (2016) A study on toxic and essential elements in wheat grain from the Republic of Kazakhstan. Environ Sci Pollut Res 23:5527–5537
Tkalec M, Štefanić PP, Cvjetko P, Šikić S, Pavlica M, Balen B (2014) The effect of cadmium-zinc interactions on biochemical responses in tobacco seedlings and adult plants. PLoS One 9:e87582
Vaughn SF, Moser BR, Dien BS, Iten LB, Thompson AR, Seliskar DM, Gallagher JL (2013) Seashore mallow (Kosteletzkya pentacarpos) stems as a feedstock for biodegradable absorbents. Biomass Bioenergy 59:300–305
Wali M, Fourati E, Hmaeid N, Ghabriche R, Poschenrieder C, Abdelly C, Ghnaya T (2015) NaCl alleviates Cd toxicity by changing its chemical forms of accumulation in the halophyte Sesuvium portulacastrum. Environ Sci Pollut Res Int 22:10769–10777
Wali M, Gunsé N, Llugany M, Corrales I, Abdelly C, Poschenrieder C, Ghnaya T (2016) High salinity helps the halophyte Sesuvium portulacastrum in defense against Cd toxicity by mainatining redox balance and photosynthesis. Planta 244:333–346
Wali M, Martos S, Pérez-Martín L, Abdelly C, Ghnaya T, Poschenrieder C, Gunsé B (2017) Cadmium hampers salt tolerance of Sesuvium portulacastrum. Plant Physiol Biochem 115:390–399
Wang SY, Jiao HJ, Faust M (1991) Changes in ascorbate, glutathione, and related enzyme activities during thidiazuron-induced bud break of apple. Physiol Plant 82:231–236
Wang Y, Wang S, Nan Z, Ma J, Zang F, Chen Y, Li Y, Zhang Q (2015) Effects of Ni stress on the uptake and translocation of Ni and other mineral nutrition elements in mature wheat grown in sierozems from northwest of China. Environ Sci Pollut Res 22:19756–19763
Weigel HJ, Jager HJ (1980) Subcellular distribution and chemical form of cadmium in bean plants. Plant Physiol 65:480–482
Yan H, Filardo F, Hu X, Zhao X, Fu DH (2016) Cadmium stress alters the redox reaction and hormone balance in oilseed rape (Brassica napus L.) leaves. Environ Sci Pollut Res 23:3758–3769
Zhang C, Sale PW, Tang C (2016) Cadmium uptake by Carpobrotus rossii (Haw.) Schwantes under different saline conditions. Environ Sci Pollut Res 23:13480–13488
Zhou B, Wang J, Guo Z, Tan H, Zhu X (2006) A simple colorimetric method for determination of hydrogen peroxide in plant tissues. Plant Growth Regul 49:113–118
Acknowledgements
We thank Mrs. Brigitte Vanpee (UCL, Louvain-la-– Neuve) for the technical assistance, and the PhD student Jiachen Wang (KUL, Leuven) for the management of the experiment. Mingxi Zhou is grateful to the China Scholarship Council (CSC) for the award of a research fellowship.
Funding
This study was partly supported by the National Natural Science Foundation of China (41403064, 41773081).
Author information
Authors and Affiliations
Corresponding author
Additional information
Responsible editor: Elena Maestri
Rights and permissions
About this article

Cite this article
Zhou, MX., Dailly, H., Renard, ME. et al. NaCl impact on Kosteletzkya pentacarpos seedlings simultaneously exposed to cadmium and zinc toxicities. Environ Sci Pollut Res 25, 17444–17456 (2018). https://doi.org/10.1007/s11356-018-1865-x
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s11356-018-1865-x