
Abstract
The evaluation of radiation exposure in 14 species of herbaceous plants from the East-Ural Radioactive Trace (EURT) zone was performed, using the ERICA Tool, v. 1.2. Recent (up to 2015) levels of radionuclide activity concentration were measured in soil and vegetative plant mass. 239,240Pu content was used for the first time to estimate external dose rates for herbaceous plant species along the pollution gradient. In addition, a new approach to assessing the geometry of objects was adopted, including not only aboveground but also underground plant organs. This improved approach to the evaluation of radiation exposure confirms previous findings that herbaceous plant populations currently exist under low-level chronic exposure in the EURT area. This reassessment based on new data suggests a 48–977-fold increase in the total dose rate per plant organism at the most polluted site compared to background areas. The highest capacity for the transfer of 90Sr and 137Cs was observed in Taraxacum officinale and Plantago major. In these species, the total dose rate per plant exceeded 150 μGy h−1 due to 90Sr + 137Cs + 239,240Pu radionuclide anthropogenic pollution in the EURT zone. All estimated total dose rates per plant were below the dose rate screening value of 400 μGy h−1.
Similar content being viewed by others
Explore related subjects
Discover the latest articles, news and stories from top researchers in related subjects.Avoid common mistakes on your manuscript.
Introduction
The Kyshtym accident occurred at the Mayak Production Association (Mayak PA) in 1957 in the Southern Urals, USSR. It resulted in the release of 7.4 PBq (10% of the radioactive waste total volume) of man-made radionuclides to the environment in a northeast direction, dispersing across a territory of 23,000 km2 (Romanov et al. 1990). At present, the East-Ural Radioactive Trace (EURT) zone is a convenient test site to evaluate the influence of radioactivity on natural ecosystems since living organisms here have been exposed to technogenic ionising radiation for over half a century (Antonova et al. 2015; Pozolotina et al. 2010). The Population Radiobiology laboratory (Institute of Plant and Animal Ecology, Russia) investigates the long-term effects of radiation on plants at the population, organism, and cell levels. Within the last decade, a wide range of biological effects were observed in wild plant populations in the EURT area: rare and unique enzyme alleles or morphs (Pozolotina and Antonova 2009; Pozolotina et al. 2007), expanding the range of variability of germination rates of seed progeny (Karimullina et al. 2015; Pozolotina and Antonova 2017), and a large number of different morphoses revealed at the early stages of ontogenesis in populations of Stellaria graminea (Pozolotina et al. 2010), Melandrium album (Antonova et al. 2013), and Bromus inermis (Antonova et al. 2014). The assessment of radiation doses is necessary to explain these radioecological and radiobiological effects due to elevated levels of technogenic radionuclides in the wild.
The potential of the ERICA Tool in assessing radiation exposure on non-human biota has been demonstrated in numerous studies (Karimullina et al. 2013; Kubota et al. 2015; Mazeika et al. 2016; Sotiropoulou et al. 2016; Wood et al. 2008). Because the accumulation of radionuclides in the phytomass depends on the chemical properties of the emitters, the chemical characteristics of the soil, and the biological features of individual species and their growing conditions, a unique combination of factors is formed in each area of operating atomic energy industrial enterprises. A considerable area of the Ural region, experiencing radioactive contamination stress, as a result of radiation accidents and routine operation of nuclear facilities, is of particular interest (Karavaeva et al. 2010). It was previously shown that the dose received by herbs exceeded the background level by several orders of magnitude in the most contaminated sites of the EURT area (Pozolotina et al. 2007; Pozolotina et al. 2012b). Later, we conducted a new assessment of doses comparing novel approaches such as the ERICA Tool 1.0 (Beresford et al. 2007; Brown et al. 2008; Larsson 2008) and RandD128/SP1a (Copplestone et al. 2001; Copplestone et al. 2003) for four herbaceous plant species growing in the EURT zone (Karimullina et al. 2013). In herbs, the interspecific accumulation capability of radionuclides may vary by four orders of magnitude (Kabata-Pendias 2010; Pozolotina et al. 2012b; Solecki et al. 2003). Therefore, the spectrum of herb species, common in temperate environments, was expanded by ten more species in our new study. Recent (up to 2015) field data of 90Sr + 137Cs radionuclide concentration and additional radionuclide 239,240Pu content in soil and plants has become available due to continuous ecological monitoring of the EURT zone. Species-specific concentration ratios of the main contaminants of the EURT area, therefore, need further evaluations which are provided in the present study.
The ERICA Tool is a continually evolving program developed by an international group of scientists (Avila et al. 2014; Brown et al. 2013; Copplestone et al. 2013; Hosseini et al. 2013). Researchers have sought to improve and modify the ERICA Tool, including efforts at consistency with developments originating from the International Atomic Energy Agency (IAEA) and International Commission on Radiological Protection (ICRP) (Brown et al. 2016).
The aim of this study was the evaluation of a radionuclide uptake and dose assessment of 14 species of herbaceous plants from the EURT zone based on (1) new data measured directly in the EURT zone, (2) using geometry of root and shoot for each plant species and (3) the analysis of the contribution of 239,240Pu in addition to the activity concentrations of 90Sr + 137Cs in soil and plants.
The ERICA Tool version 1.2, released in November 2014 (Brown et al. 2016), was used to perform dose assessment for natural herb populations at six sites along the EURT pollution gradient. The most common herb species within the area were involved in the analysis.
Materials and methods
Characterisation of sites
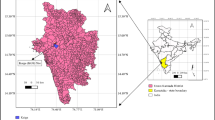
The EURT zone is located within the Trans-Ural forest-steppe, characterised by the alternation of steppe meadows, small groves of birch and birch aspen, and pine forests (Gorchakovsky 1968). The detailed biogeochemical and geobotanical description of the area was given previously (Karimullina et al. 2013; Pozolotina et al. 2012b). We investigated areas with diverse levels of radionuclide contamination in the EURT area and beyond. The map (Fig. 1) and the characterisation of sites is summarised here but is discussed elsewhere (Karimullina et al. 2013; Pozolotina et al. 2012b).

Location of the sample sites within the main part of the East-Ural Radioactive Trace and background areas. From the closest location of Mayak PA: imp-2 Impact 2 site, imp-1 Impact 1 site, buf-2 Buffer 2 site, buf-1 Buffer 1 site, bg-2 background-2 site, bg-1 background-1 site (Karimullina et al. 2013)
Two background plots are located outside the EURT area. Background 1 site [56°41′N61°02′E] is located in the birch-pine forest about 112 km from the centre of the accident. Live vegetative cover is multi-layer. The projective cover of the grass, i.e. the proportion of the ground covered by the vertical projection of vegetation (Lindenmayer and Burgman 2005), is 95%. The soil type is brown forest. Close proximity to a village area has led to a high abundance of ruderals, i.e. plant species with high survival rates on lands disturbed by natural and anthropogenic factors (Grime 1977). Background 2 site [56°47′N 61°18′E] is located in a birch-pine forest mixed with some aspen 125 km away from the centre of the accident. The projective cover of the grass is 90–95%.
Buffer 1 site [55°52′N 60°54′E] is located 15–18 km from the centre of the accident at the western border of the EURT zone. The site has a secondary forb-grass meadow, i.e. re-grown vegetation containing herbaceous flowering plants in addition to grasses, which includes 40–45 species. The projective cover of the grass is 100%. Buffer site 2 55°49′N 60°55′E] is located 15–18 km from the centre of the accident on the west bank of Lake Uruskul. The site has a secondary forb-grass meadow with birch trees. The total projective cover is 75–80%. The projective cover of the grass is 50–60%.
Impact 1 site [55°46′N 60°53′E] is located 9–12 km from the centre of the accident on the south-west bank of Lake Berdenish. Leached chernozem and subtypes of grey forest soils predominate at this site. The involvement of ruderal species in meadows and forest communities of the impact zone is reduced, while plants typical to the communities play significant roles. The total projective cover is 80–90%. Impact 2 site [55°45′N 60°50′E] is the closest to the centre of the accident, located at a distance of 6–8 km. Soils are brown forest. The most transformed plant community includes ruderals, lungwort and thistle. The total projective cover is 100%.
Field sampling and preparation
Field sampling of soil and vegetation was undertaken during the period from 2006 to 2015. Soil cores were collected randomly in 4–25 replicates at each site. To determine the level of radionuclide contamination of soil, samples were taken from soil profiles, placing them at the corners of a triangle with sides 10 m in length. The area of the soil samples was approximately and not more than 0.1 m2. Our independent preliminary studies have shown that the sampling within this scheme representatively characterises soil contamination in sites of this size. Samples of plants were taken in the immediate vicinity of the soil profile. Vegetation samples (2–10 replicates) were obtained by cutting with garden shears above the soil surface. The level of surface contamination was controlled visually, so that there was no physical contamination on the surface of the plants (soil particles easily visible to the naked eye). In a natural environment, the surface of the soil is covered with a layer of dead vegetation. Therefore, large particles of soil do not normally reach the surface of the leaves. The pubescence or stickiness of the leaves does not allow the microparticles to be washed away in some plant species. In most cases, with enhanced flushing, not only radionuclides that have entered the plant surface from the atmosphere or as a result of wind transfer with dust particles but also a part of the radionuclides taken up by other plant tissues will be lost. It is of fundamental importance not to rinse, but to take into account finely dispersed radionuclides because, being on the surface of leaves for a long time (3–4 months), they also contribute to the total dose.
Radiochemical analysis was performed to determine the 90Sr content in soil and vegetation samples. The method was based on the leaching of radiostrontium with 6 N HCl-solution after preliminary preparation of samples. Separated 90Sr was kept in balance with its daughter product of decay, 90Y, after precipitation of 90Sr in the oxalate form. Measurements of 90Y were established using an α-β-radiometer with a siliceous detector (Russia). The detection limit was 0.2 Bq.
The radionuclide analysis of 137Cs content was performed using γ-ray spectroscopy, for which the Canberra-1510 multichannel analyser and GENIE-PC software (Canberra-Packard, USA) germanium detectors were used. The detection limit was 0.1 Bq.
Radiochemical determination of 239,240Pu included several steps, such as leaching isotopes by mixed concentrated acids (HNO3 + HCl) from preliminary prepared samples, precipitating plutonium on ion-exchange resin, purification, stripping from the ion-exchange column by H2O and 0.5 N HNO3 solution in succession and electro-deposition on disks made of stainless steel (Chen et al. 1993). The system used for the measurements was an α-spectrometer Alpha Analyst (Canberra-Packard, USA) with semiconductor detectors (PIPS) and GENIE-2000 software. The lower limit of detection was 0.001 Bq. The procedural error of the methods did not exceed 20%. A correction for radioactive decay was applied for samples collected in different years.
Test organisms
Fourteen common herbaceous plant species capacity to accumulate radionuclides was analysed. Dominant or codominant herbs, analysed in this study, include little starwort (Stellaria graminea L.), white campion (Melandrium album (Mill.) Garcke (= Silene latifolia)), smooth brome (Bromopsis inermis Leyss. (= Bromus inermis)), quinquelobate motherwort (Leonurus quinquelobatus Gilib.), creeping thistle (Cirsium setosum (= Cirsium arvense s.l.), yellow sweet clover (Melilotus officinalis L.), common nettle (Urtica dioica L.), woolly burdock (Arctium tomentosum Mill.), common dandelion (Taraxacum officinale s.l.), broadleaf plantain (Plantago major L.), Russian dock (Rumex confertus Willd.), zigzag clover (Trifolium medium L.), meadow pea (Lathyrus pratensis L.) and common wormwood (Artemisia vulgaris L.). These plant species represent 14 genera and 8 families in our study.
Data analysis
To estimate the doses of external and internal radiation exposure using the ERICA Tool program, the average values of parameters (weight, height, width, length) were used for 14 species of plants commonly found in the surveyed territory (see supplementary material, Tables 1 and 2) The following assumptions characterise the model used in this study:
-
1.
The dimensional parameters of the vegetation mass (see supplementary material, Table 1) of plants of different species were averaged for each species and are in good agreement with botanical literature for this region (Gorchakovsky et al. 1964).
-
2.
The average rates of activity concentrations of man-made radionuclides were used in each species of herbaceous plant for each investigated site, because the content of radionuclides in plants varies widely, depending on the type and age of the plants, the weather conditions during the vegetation period, the way radionuclides enter the soil, the properties of the soil and other factors (Molchanova et al. 2014a; Shcheglov and Tsvetnova 2001; Willey and Tang 2006).
-
3.
One of three ellipsoid dimensions, the height of the roots, was taken as 0.1 m for all herbaceous species (see supplementary material, Table 2), as the bulk of the roots is located in this the most contaminated layer of soil. The other two ellipsoid dimensions, width and length of the roots, forming a square area, were taken as equal to the projective vegetation mass; this parameter was conditional and unified for all herb species. This assumption was made according to previous findings that in the forest-steppe zone, the ratio of underground and aboveground phytomass in the out-of-place meadows, steppes and agricultural lands is close to 1 (Ravkin et al. 2011). We also observed that the mass of living and dead roots and the aboveground vegetation mass (living and dead) per one square meter were approximately the same (our unpublished data).
The ERICA Tool uses activity concentrations of vegetation on a fresh weight basis. The radionuclide content measurement procedure uses dried samples of vegetation. Therefore, we computed the fresh weight using the dry/wet ratio for Stellaria graminea = 1/2.5; Bromus inermis, Melilotus officinalis, Urtica dioica, and Leonurus quinquelobatus = 1/2; Melandrium album, Arctium tomentosum, Artemisia vulgaris, Cirsium setosum, Lathyrus pratensis, and Rumex confertus = 1/3; Plantago major, Taraxacum officinale, and Trifolium medium = 1/4. The difference was determined by weighing freshly harvested samples of vegetation before and after complete drying. This allowed us to calculate the concentration ratio for the wild organism (CRwo-soil) for each of the 14 species of herbaceous plants. The standard equation (Beresford et al. 2007; Karimullina et al. 2013) was used for this purpose. Data are presented as the arithmetic mean (AM) ± standard deviation (SD). Dose assessment was performed using the ERICA Tool 1.2 software (Brown et al. 2016).
Results and discussion
Contamination gradient of soil
The spatial distribution of radionuclides in the EURT area was estimated in our previous works (Molchanova et al. 2009, 2011, 2014b). Within the central axis, the contamination densities of radionuclides change with increasing distance from the centre of the accident and satisfactorily approximate a power function (for example, stock of 90Sr changes from 69 MBq m−2 to 0.4 MBq m−2). On the eastern periphery of the trace zone, the value of 90Sr and 137Cs stocks are almost identical and with increasing distance reduce from 100 to 10 kBq m−2. On the western periphery of the trace, the content of 137Cs in soil, regardless of the distance, is kept at a level (5–10) kBq m−2. The stocks of 239,240Pu are much lower and change from 5 to 0.2 kBq m−2 with distance from the pollution source. On the whole territory of the EURT, the 20 cm soil layer keeps 80–92% of the total 90Sr, 137Cs and 239,240Pu stocks. Thus, these radionuclides are characterised by low mobility in the soil profiles.
In our previous work (Karimullina et al. 2013), two main EURT contaminant radionuclides in soil, 90Sr and 137Cs, were used to calculate the external dose rates. In the present study, we additionally included 239,240Pu content in soil and vegetation to estimate its impact on the total exposure of plant populations to technogenic radionuclides. Our present data demonstrate an increase in 90Sr levels in soil at the Impact 2 site to 15,500 or even 92,700 times the pollution levels observed at the two background sites. Notably, the Impact 2 site is located closest to the site of the Kyshtym accident (Table 1). 239,240Pu- and 137Cs-specific activity in Impact 2 soil samples was 2 and 3 orders of magnitude higher than those from background sites respectively. Soil samples from buffer sites had intermediate radionuclide activity concentrations rates.
Activity concentrations in 14 herbaceous species in the EURT area
Radionuclide activity concentrations in biota are required for Tier 2 and 3 assessments using the ERICA Tool (Beresford et al. 2007). As for radionuclide concentrations in soil and plants, sufficient data are available in our case.
The activity concentrations of 90Sr, 137Cs and 239,240Pu were used to calculate the transfer parameters in 14 species of herbaceous plants from the EURT zone and background area. These data were determined empirically and are presented in Table 2. The lowest levels of 90Sr and 137Cs specific activity were observed in A. Tomentosum and L. quinquelobatus vegetative mass, especially at background sites. The minimum activity concentration of 239,240Pu was observed in L. pratensis. The highest activity concentrations were in T. officinale (90Sr and 137Cs) and two species from the Caryophyllaceae family (M. album and S. graminea) (239,240Pu) collected in impact plots. Previously, we observed a low capacity for biological transition to herbaceous plants for 137Cs at the EURT zone (Karimullina et al. 2013; Pozolotina et al. 2012b). The major contribution to internal dose levels is made by 90Sr radionuclide in the technogenic EURT area. In the background sites, the ratio of activity concentrations of 90Sr/137Cs and 90Sr/239,240Pu radionuclides was approximately 19:1 and 300:1 in the vegetative mass, respectively. In the EURT impact plots, that ratio reached 700:1 and 5000:1, respectively.
According to different studies, the ratio of concentrations of 90Sr and 137Cs in the roots of plants and plant organs above the ground varies greatly. This ratio may be either larger or smaller than one. Moreover, there are data indicating that when the conditions of mineral nutrition of plants change, the ratio of radionuclide concentrations in the aboveground vegetation mass and roots may change significantly even within the same species (Willey and Tang 2006). There are no such data for plutonium. However, there is evidence of an accumulation of its iron macro-analogue (Afanasyeva and Kashin 2015; La Toya et al. 2010), which allows the use of a root/shoot ratio equal to 1. Our spatial empirical data on the accumulation of radionuclides in the roots of herbaceous plants from the EURT zone allow us to proceed from the assumption that the concentrations in the aboveground mass and in the roots of the plants may be the same.
Input CRwo-soil values data
A standard equation (Beresford et al. 2007; Karimullina et al. 2013) was used to define the concentration ratios (CRwo-soil) of 14 herbaceous species for the 90Sr, 137Cs and 239,240Pu radionuclides (Table 3). The highest uptake of 90Sr was observed in T. officinale. The next highest uptake was observed in P. major followed by M. album. U. dioica and T. medium, and C. arvense had intermediate CRwo-soil values. The lowest uptake was revealed in B. inermis, S. graminea and A. tomentosum species. The 137Cs CRwo-soil was low in most species. The weakest transfer of 137Cs was identified in L. quinquelobatus, B. inermis, A. tomentosum, M. officinalis and S. graminea species. M. album, T. medium, L. pratensis and U. dioica are grouped with ascending values of 137Cs CRwo-soil. The highest transfer capacity for 137Cs was observed in T. officinale. Minimal CR values of 239,240Pu were determined in L. pratensis, which were two orders of magnitude lower than the maximal rates observed in S. graminea and M. officinalis. Overall, radionuclide 90Sr showed higher soil-to-organism transfers than 137Cs and 239,240Pu in 14 herb species in the EURT area.
Patterns of transfer of 90Sr, 137Cs and 239,240Pu radionuclides from soil to plant were assessed by grouping herb taxa by their taxonomic families. The groups of families such as Asteraceae, Caryophyllaceae and Fabaceae presented by several species had high variability in interspecific CR rates and had no significant pattern that would indicate potent or weak uptake of radionuclides in the whole family. The absolute maximum CR rate of 137Cs and the minimum CR rate of 90Sr belong to members of Asteraceae. The maximum and minimum CR level of 239,240Pu was observed in the Fabaceae family (see Table 3).
Dose assessment using empirically derived data in soil and vegetation of 14 herbaceous species
The ERICA Tool has an input model of herbs as an ellipsoid with an on-soil occupancy factor. Therefore, previously, we used this model to analyse total dose rates. However, this approach misses the fact that the root system, representing in most cases half of the plant organism, is exposed to external doses at increased rates compared to that above the soil. Since the application of realistic occupancy factors will lead to less conservative dose estimates, we entered 28 new geometries as 28 new vegetation organisms: 14 geometries for shoots with occupancy factor on-soil equal to 1.0 and roots with occupancy factor in-soil equal to 1.0. This was performed for 14 herb species characterised by various root systems, mass, and ellipsoid dimensions (see supplementary material, Tables 1 and 2). In the wildlife group of plants, the specimen was chosen as “vegetation,” and the “herb” subtype was picked. Internal and external dose conversion coefficient (DCC) values were calculated. Since the only way to calculate DCC for root and shoot separately in the ERICA Tool is to enter them into the software as two new organisms (Biermans et al. 2014), these models were used to analyse doses at each site using the Tier 2 approach. Soil concentrations used for the analysis and CR values are presented in Tables 1 and 3, respectively. The weighting factor for alpha radiation was set as 10 by default; the low-level β-radiation was set constant at 3, and for high-energy γ- and β-radiation, it was set at 1.
The external dose rates for terrestrial herb species were mainly due to the 137Cs radionuclide (see supplementary material, Table 3). The external dose rate increased by 3–4 orders of magnitude compared to the background rate along the pollution gradient due to anthropogenic 90Sr + 137Cs + 239,240Pu radionuclide pollution. The main technogenic radionuclide contributing to the estimated internal dose rate was 90Sr at the Impact sites, while in background plots, all three radionuclides contributed mainly equally (see supplementary material, Table 4). Due to external and internal contributing radiostrontium and radiocesium, the values of the total dose rate per root or shoot model derived from these two radionuclides were higher by 1–3 orders of magnitude compared with the values of 239,240Pu at the polluted sites (see supplementary material, Table 5). The values of the total dose rate calculated using the ERICA Tool Tier 2 approach (Table 4) varied slightly between root and shoot ellipsoids within each plant due to slightly different dimensions used for input (see supplementary material, Tables 1 and 2). At the most polluted Impact 2 site, total dose rates of T. officinale and P. major were the highest (see Table 4).
Discussion
The gradual moving from the principle that the observance of radiation dose limits established for humans is a sufficient guarantee for the protection of biota is already declared in publication 103 of the International Commission on Radiological Protection (ICRP 2007). In the subsequent publication 108 (ICRP 2008), the ICRP expanded this concept and introduced the concept of “reference” organisms, which are hypothetical objects with biological characteristics generalised at the family level, derivatives of the “reference levels,” as expressed by the dose rate of radiation and which are proposed for each of them based on the data on the radiation effects observed at different doses. For the purposes of protecting the environment under irradiation conditions, the ICRP proposes to consider the species directly exposed to radiation as representative organisms. The doses of irradiation of representative organisms are compared with the derived reference levels established for close reference organisms. There are limited possibilities of ensuring the radiation safety of biota in the situation of radioactive contamination compared a set of methods developed for human protection (ICRP 2009, 2014). Doses of irradiation on animals and plants living in radioactively contaminated areas cannot be reduced by the resettlement, removal or removal of contaminated soil, etc. At the same time, the protection of non-human biota can be based on measures that while not directly affecting the source of radiation contribute to the conservation of biological diversity. In addition, when considering the possible approaches to ensuring the radiation safety of biota, it is necessary to study the mechanisms of the adaptation of animals and plants, relating them to the dose loads on specific populations, and also taking into account the interaction of the radiation factor with other environmental conditions.
In the first years after the Kyshtym disaster, environmental protection was not raised. The accident was classified as communal (agrarian), since agricultural and forest products containing radionuclides were the main source of human exposure. Fundamental studies of the migration and accumulation of radionuclides in living and stagnant components of ecosystems were associated with applied problems, with the development of recommendations for reducing radionuclide intake into plants and animals. These works were extremely important in the future in the aftermath of accidents at the nuclear power plants in Chernobyl (1986) and Fukushima (2011).
At present, the assessment of the consequences of a radiation accident in the Urals shows that the forests and meadows in the EURT area have recovered completely. Community biodiversity is assessed even higher than in the adjacent areas (Lagunov and Smagin 2007; Sokolov and Krivolutskiy 1993). This is due to several reasons: (1) there is always a seed stock in the soil that can germinate when dose loads have decreased; (2) the territory of the zone is narrow (6–12 km wide), and the introduction of seeds from “clean” areas, like the migration of animals, is possible; (3) perennial rhizome hemicryptophytes, with good protection of the buds of renewal, predominate in the EURT zone, and annuals are almost never found. The only countermeasure that contributed to the restoration of natural communities was the lack of anthropogenic load on this area (Pozolotina et al. 2012b). However, this does not exclude the possibility of active measures contributing to the restoration of natural communities in radioactive contamination.
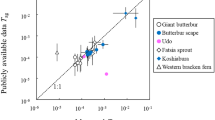
The mean values of the strontium soil-to-plant transfer factor for herbs in the temperate environment were reported in Table 5 of TRS-479 as 9.8E−01 for all soil types (IAEA 2014). Our CR data are in the same order of magnitude for most species (see Table 3) including Artemisia vulgaris, Cirsium arvense, Taraxacum officinale, Melandrium album, Lathyrus pratensis, Melilotus officinalis, Trifolium medium, Leonurus quinquelobatus, Rumex confertus, Plantago major, and Urtica dioica. The other three species, Arctium tomentosum, Stellaria graminea, and Bromus inermis, had lower rates of CR by one order of magnitude. In contrast, no trend was observed in species specificity for 137Cs. Radiostrontium CR rates were two orders of magnitude lower for all studied species than the rate published in TRS-479 for grasses and herbs (1.2E+0) (IAEA 2014). It should be noted that the TRS-479 report does not specify CR values for each isotope giving instead averaged data for all isotopes of one element. Our mean CR values of 239,240Pu were in good agreement for most of the investigated herbaceous species with the mean transfer factor value of plutonium (1.6E−02) for grasses and herbs given in TRS-479 (IAEA 2014). There was only Lathyrus pratensis which had a mean value (see Table 3) two orders of magnitude lower than that calculated by IAEA.
At the same time, the mean CR values of 239,240Pu reported by Strand et al. (2009) were in agreement with our CR data gathered from Melilotus officinalis, Stellaria graminea, Melandrium album, and Arctium tomentosum. We observed decreased CR rates of 90Sr and 137Cs compared to ICRP data for herbs. With a rare exceptions (e.g., CR value of 90Sr in Taraxacum officinale), the distribution of our new measured CR data was below the rates for soil-to-plant transfer factor values of radiostrontium and radiocesium for the temperate environment (Fig. 2) given in TRS-479 (IAEA 2014) and the ICRP report for CR values of plutonium in wild grasses in terrestrial ecosystems (Strand et al. 2009).

All the estimated total dose rates per plant (see Table 4) were below the dose rate screening value of 400 μGy h−1 (USDoE 2002) and higher than no-effect dose rate of 10 μGy h−1 predicted in the ERICA risk assessment (Garnier-Laplace et al. 2008) for all herbaceous plants at the most polluted EURT zone. The ERICA Tool includes a tab summarising the radiobiological effects data as determined in conjunction with information from the FREDERICA radiation effects database (Copplestone et al. 2008). These data indicate the various effects in the Taraxacum species (dandelion) with rate ranges of 4–70 μGy h−1. In our assessment, the total dose rate for the Taraxacum species was 93 and 199 μGy h−1 at the Impact 1 and Impact 2 sites, respectively. In addition, another approach using Derived Consideration Reference Levels (DCRLs) indicated the possible radiation effects within the described exposure situation in the EURT zone. The ICRP described its approach to the protection of the environment (Copplestone et al. 2016) and derived DCRL for grass as 1–10 mGy/day (i.e., 42 to 420 μGy/h) (ICRP 2008). In fact, the total dose per 6 out of 14 species and 9 out of 14 species at the Impact 1 and Impact 2 sites, respectively, is within DCRL for grass.
At the buffer EURT sites, the average contribution to the radiation dose due to 90Sr was 67%, 137Cs − 31% and 239,240Pu − 1.7%. At the impact EURT sites, the domination of 90Sr in the formation of the radiation dose increased (93%), while the contribution of 137Cs and 239,240Pu was minimal (6 and 1%, respectively). However, the 239,240Pu decay period is considerably higher than that of 90Sr and 137Cs. Therefore, the impact of radiation on non-human biota will persist for a long time in the EURT area.
The ERICA Integrated Approach should be used to assess the incremental doses from human activities only (Beresford et al. 2007). However, natural background radiation acts on all ecosystems. Its contribution to the background sites was significant for herbs (0.1–0.2 μGy h−1), while the 90Sr, 137Cs and 239,240Pu total dose rates ranged from 0.001 to 0.01 μGy h−1 (see Table 4). The opposite pattern was observed at the impact sites. Considering the summarised dose rates due to natural radiation and anthropogenic radionuclide pollution, our results point to a 48–977-fold increase in the dose rate per plant organism at the most polluted site compared to the background. These findings extend our understanding of the link between the anthropogenic impact and plant populations growing in the wild.
The biological effects of technogenic radiation were not the goal of this work; however, without assessing the dose rates on natural communities from the zones of radioactive contamination, it is difficult to understand the effects of radiation on non-human biota. One of the crucial challenges we undergo in our field studies to assess dose rates is that it is generally impossible to separate individual plants in natural populations of rhizome plants (cereals, for example) because they form a single cover, namely the sod. The roots of the individual parts are interconnected by rhizomes making the exchange of nutrients, and therefore radionuclides, possible.
Earlier, we compared the dose rates of β-radiation measured with a natural grass stand and after cutting these plants off at sites in the EURT zone (Pozolotina et al. 2012a). In the herbage, two processes are superimposed. On the one hand, plant biomass causes the weakening of radiation from the soil surface (screening effect). On the other hand, radionuclides accumulated in plants contribute to radiation. Both morphological features of the species and their ability to accumulate radionuclides play an important role in the manifestation of the total doses. On sites where the Bromus inermis dominates, significant differences with undisturbed grass and without it are absent. At the same time, Urtica dioica leaves at an altitude of 10–50 cm create a significant screening effect (see Table 5). The exposure dose of γ radiation with height has no change.
Based on data on radionuclide concentrations in different parts of plants, we calculated the contribution of incorporated radionuclides to the total irradiation dose of the herbage. Radionuclides accumulated in the rump biomass were added from 1.4 to 4.2% to the total dose, while the contribution of radionuclides in leaves and stems of nettle at an altitude of 40–100 cm reached 22–27%. These results are in good agreement with the literature data (Bensen and Sparrow 1971; Prister 2008). The total dose of β-radiation from the loss of primary products of nuclear fission decreased by a factor of two regarding the height of cereal crops. In the lower tier, the contribution of irradiation from the soil surface predominated, and radiation from radionuclides accumulated by biomass prevailed in the upper tier (Prister 2008). A survey of a meadow community polluted with 137Cs showed that the accumulation of radionuclides in the biomass leads to the formation of a volumetric radiation source (Bensen and Sparrow 1971).
Therefore, as a promising future direction, we would like to recommend ERICA developers to provide a user-friendly option for a dose calculation of the dose rates not only in individual plants, but in a whole grass stand.
Conclusions
This improved approach to the evaluation of radiation exposure confirms previous findings that herbaceous plant populations in the EURT area currently exist under low-level chronic exposure. A reassessment based on new expanded data suggests a 48–977-fold increase in the total dose rate per plant at the most polluted site as compared to the background. The highest capacity for the transfer of 137Cs and 90Sr was observed in T. officinale and P. major. The total dose rate per plant exceeded 150 μGy h−1 in T. officinale and P. major due to 90Sr + 137Cs + 239,240Pu radionuclide anthropogenic pollution in the EURT area.
At the buffer EURT sites, the average contribution to the radiation dose due to 90Sr was 67%, 137Cs 31% and 239,240Pu 1.7%. At the impact EURT sites, the domination of 90Sr in the formation of the radiation dose increased (93%), while the contribution of 137Cs and 239,240Pu was minimal (6 and 1%, respectively).
As a possible recommendation, we suggest the introduction into the program of an approach that takes into account the total doses not only for individual plants but for a grass stand. The calculation of the geometry of individual plants is very conditional, and grasses form in nature, as a rule, an organic whole, which is a grass stand of different specific density, and they can irradiate each other due to radionuclides accumulated in the aboveground mass.
References
Afanasyeva LV, Kashin VK (2015) The chemical composition and productivity of Vaccinium myrtillus L. under influence of industrial pollution. J Sib FU Biol 8:333–346
Antonova E, Karimullina E, Pozolotina V (2013) Intraspecific variation in Melandrium album along a radioactive contamination gradient at the eastern Ural radioactive trace. Russ J Ecol 44:18–27
Antonova E, Pozolotina V, Karimullina E (2014) Variation in the seed progeny of smooth brome grass, Bromus inermis Leyss., under conditions of chronic irradiation in the zone of the eastern Ural radioactive trace. Russ J Ecol 45:508–516
Antonova EV, Pozolotina VN, Karimullina EM (2015) Time-dependent changes of the physiological status of Bromus inermis Leyss. seeds from chronic low-level radiation exposure areas. Biol Rhythm Res 46:587–600
Avila R, Beresford N, Brown J, Hosseini A (2014) The selection of parameter values in studies of environmental radiological impacts. J Radiol Prot 34:260
Bensen DW, Sparrow AH (1971) Survival of food crops and livestock in the event of nuclear war. Proc Confer S Springfield:509–520
Beresford N, Brown J, Copplestone D, Garnier-Laplace J, Howard B, Larsson C-M, Oughton D, Prohl G, Zinger I (2007) D-ERICA: an integrated approach to the assessment and management of environmental risk from ionising radiation. Description of purpose, methodology and application. p. 82
Biermans G, Horemans N, Vanhoudt N, Vandenhove H, Saenen E, Van Hees M, Wannijn J, i Batlle JV, Cuypers A (2014) An organ-based approach to dose calculation in the assessment of dose-dependent biological effects of ionising radiation in Arabidopsis thaliana. J Environ Radioact 133:24–30
Brown JE, Alfonso B, Avila R, Beresford NA, Copplestone D, Pröhl G, Ulanovsky A (2008) The ERICA tool. J Environ Radioact 99:1371–1383
Brown JE, Beresford NA, Hosseini A (2013) Approaches to providing missing transfer parameter values in the ERICA tool–how well do they work? J Environ Radioact 126:399–411
Brown J, Alfonso B, Avila R, Beresford N, Copplestone D, Hosseini A (2016) A new version of the ERICA tool to facilitate impact assessments of radioactivity on wild plants and animals. J Environ Radioact 153:141–148
Chen Q, Aarkrog A, Nielsen S, Dahlgaard H, Nies H, Yixua Y, Mandrup K (1993) Determination of plutonium in environmental samples by controlled valence in aniion exchange. J Radioanal Nucl Chem 172:281–288
Copplestone D, Bielby S, Jones S (2001) Impact assessment of ionising radiation on wildlife. Environment Agency, London
Copplestone D, Wood MD, Bielby S, Jones SR, Vives J, Beresford NA (2003) Habitat regulations for Stage 3 assessments: radioactive substances authorisations. R&D Technical Report P3-101/SP1a. Environment Agency, Bristol
Copplestone D, Hingston J, Real A (2008) The development and purpose of the FREDERICA radiation effects database. J Environ Radioact 99:1456–1463
Copplestone D, Beresford N, Brown J, Yankovich T (2013) An international database of radionuclide concentration ratios for wildlife: development and uses. J Environ Radioact 126:288–298
Copplestone D, Larsson C-M, Strand P, Sneve MK (2016) Protection of the environment in existing exposure situations. Ann ICRP 45:91–105
Garnier-Laplace J, Copplestone D, Gilbin R, Alonzo F, Ciffroy P, Gilek M, Agüero A, Björk M, Oughton DH, Jaworska A, Larsson CM, Hingston JL (2008) Issues and practices in the use of effects data from FREDERICA in the ERICA integrated approach. J Environ Radioact 99:1474–1483
Gorchakovsky PL (1968) The European broad-leaved forest plants at their eastern distribution limits. Academy of Science USSR, Sverdlovsk (in Russian)
Gorchakovsky PL, Shurova EA, Knyazev MS, Marina LV, Morozova LM, Nikonova NN, Pryamonosova SA, Salmina HP, Shlykova NA, Belyaeva IV, Balandin SV, Vasfilova ES, Famelis TV, Trotsenko GV, Zueva VN, Meling EA (1964) The determinant of vascular plants of the middle Urals. Nauka, Moscow (in Russian)
Grime JP (1977) Evidence for the existence of three primary strategies in plants and its relevance to ecological and evolutionary theory. Am Nat 111(982):1169–1194
Hosseini A, Stenberg K, Avila R, Beresford NA, Brown JE (2013) Application of the Bayesian approach for derivation of PDFs for concentration ratio values. J Environ Radioact 126:376–387
IAEA (2014) Handbook of parameter values for the prediction of radionuclide transfer to wildlife. Technical Reports Series. International Atomic Energy Agency, Vienna
ICRP (2007) Recommendations of the international commission on radiological protection (users edition). Ann ICRP 37 (2-4)). ANN ICRP 37 publication 103 (users edition): 2-4
ICRP (2008) Environmental protection—the concept and use of reference animals and plants. ICRP publication 108 ANN ICRP 38: 4-6
ICRP (2009) Environmental protection: transfer parameters for reference animals and plants ANN ICRP 6
ICRP (2014) Protection of the environment under different exposure situations ANN ICRP 1
Kabata-Pendias A (2010) Trace elements in soils and plants. CRC press, Boca Raton, p 548
Karavaeva Y, Mikhailovskaya L, Molchanova I, Pozolotina V (2010) Accumulation of 90Sr and 137Cs by plants from soils contaminated due to the operation of the enterprises of the nuclear energy sector. Problems of biogeochemistry and geochemical. Ecology 1:85–90
Karimullina E, Antonova E, Pozolotina V (2013) Assessing radiation exposure of herbaceous plant species at the east-Ural radioactive trace. J Environ Radioact 124:113–120
Karimullina E, Antonova E, Pozolotina V, Tokarev A, Minko S (2015) The toxicity of engineered nanoparticles on seed plants chronically exposed to low-level environmental radiation. Russ J Ecol 46:236–245
Kubota Y, Takahashi H, Watanabe Y, Fuma S, Kawaguchi I, Aoki M, Kubota M, Furuhata Y, Shigemura Y, Yamada F, Ishikawa T, Obara S, Yoshida S (2015) Estimation of absorbed radiation dose rates in wild rodents inhabiting a site severely contaminated by the Fukushima Dai-ichi nuclear power plant accident. J Environ Radioact 142:124–131
La Toya TK, Jacob DL, Otte ML (2010) Multi-element accumulation near Rumex crispus roots under wetland and dryland conditions. Environ Pollut 158:1834–1841
Lagunov A, Smagin A (2007) Role of the east-Ural state reservation in the system of highly guarded natural territories of the Chelyabinsk region. Prob Radiat Saf S:45-67 (in Russian)
Larsson C-M (2008) An overview of the ERICA integrated approach to the assessment and management of environmental risks from ionising contaminants. J Environ Radioact 99:1364–1370
Lindenmayer D, Burgman M (2005) Practical conservation biology. Csiro Publishing, p 624
Mazeika J, Marciulioniene D, Nedveckaite T, Jefanova O (2016) The assessment of ionising radiation impact on the cooling pond freshwater ecosystem non-human biota from the Ignalina NPP operation beginning to shut down and initial decommissioning. J Environ Radioact 151:28–37
Molchanova I, Pozolotina V, Karavaeva Y, Mikhaylovskaya L, Antonova E, Antonov K (2009) Radioactive inventories within the east-Ural radioactive state reserve on the southern Urals. Radioprotection 44(5):747–757
Molchanova IV, Pozolotina VN, Antonova EV, Mikhaylovskaya LN (2011) The impacts of permanent irradiation on the terrestrial ecosystems of the eastern-Ural radioactive trace. Radioprotection 46:567–572
Molchanova I, Mikhaĭlovskaia L, Pozolotina V, Antonova E (2014a) Man-made radionuclides and their accumulation by plants of different taxonomic groups from the soils of the eastern Ural radioactive trace. Radiats Biol Radioecol 54:77–84
Molchanova I, Mikhailovskaya L, Antonov K, Pozolotina V, Antonova E (2014b) Current assessment of integrated content of long-lived radionuclides in soils of the head part of the east Ural radioactive trace. J Environ Radioact 138:238–248
Pozolotina V, Antonova E (2009) Intrapopulation variation in the quality of dandelion seed progeny in zones of chemical and radioactive contamination. Russ J Ecol 40:361–367
Pozolotina VN, Antonova EV (2017) Temporal variability of the quality of Taraxacum Officinale seed progeny from the east-Ural radioactive trace: is there an interaction between low level radiation and weather conditions? Int J Radiat Biol 93:330–339
Pozolotina V, Molchanova I, Karavaeva E, Mihkaylovskaya L, Antonova E, Karimullina E (2007) Analysis of current state of terrestrial ecosystems in the east-Ural radioactive trace. The issues of the radiation safety (special issue ‘the east Ural radioactive trace marks its 50 year anniversary’) pp 32-44
Pozolotina VN, Antonova EV, Karimullina EM (2010) Assessment of radiation impact on Stellaria graminea cenopopulations in the zone of the eastern Ural radioactive trace. Russ J Ecol 41:459–468
Pozolotina VN, Antonova EV, Onishchenko AD, Seleznev AA (2012a) Assessment of external irradiation doses for herbage in the east-Ural radioactive trace area. Problems of biogeochemistry and geochemical. Ecology 4:71–78
Pozolotina VN, Molchanova IV, Mikhailovskaya LN, Antonova EV, Karavaeva EN (2012b) In: Gerada JG (ed) The current state of terrestrial ecosystems in the eastern Ural radioactive trace. Nova Science Publishers, Inc., New York, pp 1–21
Prister BS (2008) The problems of agricultural radioecology and radiobiology at radiation environmental pollution by a young mixture of nuclear fission products. Ins. for safety problems of nuclear power plants of NAS of Ukraine, Chornobyl, p 320 (in Russian)
Ravkin YS, Sedel’nikov V, Sergeev M, Titlyanova A, Khmelev V, Bogomolova I, Tsybulin S (2011) Spatial-typological differentiation of ecosystems of the west Siberian plain. Communication V: terrestrial ecosystems. Contemp Probl Ecol 4:568–581
Romanov GN, Nikipelov BV, Drozhko EG (1990) The Kyshtym accident: causes, scale and radiation characteristics. Seminar on comparative assessment of the environmental impact of radionuclides released during three major nuclear accidents: Kyshtym, Windscale, Chernobyl. Commission of the European Communities, Luxemburg 1:25–40
Shcheglov AI, Tsvetnova OB (2001) Biogeochemical migration of technogenic radionuclides in forest ecosystems: by the materials of a multiyear study in the areas severely contaminated due to the Chernobyl accident. Cornell University, Nauka, p 234
Sokolov V, Krivolutskiy D (1993) Ecological consequences of radioactive contamination in the southern Urals. Nauka, Moscow (in Russian)
Solecki J, Reszka M, Chibowski S (2003) 90Sr and 137Cs radioisotopes and heavy metal concentrations in pharmaceutical herbal plants from the Lublin (Poland) region. J Radioanal Nucl Chem 257:261–265
Sotiropoulou M, Florou H, Manolopoulou M (2016) Radioactivity measurements and dose rate calculations using ERICA tool in the terrestrial environment of Greece. Environ Sci Pollut Res:1–11
Strand P, Beresford N, Copplestone D, Godoy J, Jianguo L, Saxén R, Yankovich T, Brown J (2009) Environmental protection: transfer parameters for reference animals and plants. Ann ICRP 39:1–111
USDoE A (2002) Graded approach for evaluating radiation doses to aquatic and terrestrial biota, technical standard DOE-STD-1153-2002. USDOE, Washington, DC
Willey N, Tang S (2006) Some effects of nitrogen nutrition on caesium uptake and translocation by species in the Poaceae, Asteraceae and Caryophyllidae. Environ Exp Bot 58:114–122
Wood MD, Marshall WA, Beresford NA, Jones SR, Howard BJ, Copplestone D, Leah RT (2008) Application of the ERICA integrated approach to the Drigg coastal sand dunes. J Environ Radioact 99:1484–1495
Funding
This study was performed within the frameworks of state contract with the Institute of Plant and Animal Ecology, Ural Branch, Russian Academy of Sciences, and partly supported by the Program of UB RAS (projects 15-2-4-21, 18-4-4-9).
Author information
Authors and Affiliations
Corresponding author
Additional information
Responsible editor: Georg Steinhauser
Electronic supplementary material
ESM 1
(DOCX 43 kb)
Rights and permissions
About this article

Cite this article
Karimullina, E.M., Mikhailovskaya, L.N., Antonova, E.V. et al. Radionuclide uptake and dose assessment of 14 herbaceous species from the east-Ural radioactive trace area using the ERICA Tool. Environ Sci Pollut Res 25, 13975–13987 (2018). https://doi.org/10.1007/s11356-018-1544-y
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s11356-018-1544-y