
Abstract
Pseudomonas aeruginosa strain GF31, isolated from a contaminated soil, can effectively degrade β-cypermethrin (β-CP), as well as fenpropathrin, fenvalerate, and cyhalothrin. The highest level of degradation (81.2 %) was achieved with the addition of peptone. Surprisingly, the enzyme responsible for degradation was mainly localized to the extracellular areas of the bacteria, in contrast to the other known pyrethroid-degrading enzymes, which are intracellular. Although intact bacterial cells function at about 30 °C for biodegradation, similar to other degrading strains, the crude extracellular extract of strain GF31 remained biologically active at 60 °C. Moreover, the extract fraction showed good storage stability, maintaining >50 % of its initial activity following storage at 25 °C for at least 20 days. Significant differences in the characteristics of the crude GF31 extracellular extract compared with the known pyrethroid-degrading enzymes indicate the presence of a novel pyrethroid-degrading enzyme. Furthermore, the identification of 3-phenoxybenzoic acid and 2,2-dimethylcyclopropanecarboxylate from the degradation products suggests the possibility that β-CP degradation by both the strain and the crude extracellular fraction is achieved through a hydrolysis pathway. Further degradation of these two metabolites may lead to the development of an efficient method for the mineralization of these types of pollutants.
Similar content being viewed by others

Explore related subjects
Discover the latest articles, news and stories from top researchers in related subjects.Avoid common mistakes on your manuscript.
Introduction
Pyrethroids, a type of insecticide previously considered to have relatively low toxicity, have been used worldwide in place of more toxic pesticides for nearly 30 years. However, recent evidence has shown their cumulative (Liu et al. 2010), reproductive (Ahmad et al. 2009), and neurotoxicological toxicity (Shafer et al. 2005; Wolansky and Harrill 2008) to humans and aquatic organisms. Beta-cypermethrin (β-CP), an important synthetic pyrethroid insecticide, shows better photostability than natural pyrethroids and a long effective time but is resistant to degradation. An investigation of the urban waterways and sediments of the Chinese Pearl River Delta showed that β-CP was the most abundant pyrethroid detected (Mehler et al. 2011; Li et al. 2011), representing a significant threat to the environment.
Biodegradation involves the use of living microorganisms or active enzymes to detoxify and degrade hazardous materials (Thouand 2014). Among the various pyrethroid-degrading microbes (Saikia and Gopal 2004; Chen et al. 2013), a few Serratia species (Zhang et al. 2010a), Pseudomonas aeruginosa (Li et al. 2009; Zhang et al. 2011), and Ochrobactrum lupini (Chen et al. 2011) strains can degrade β-CP at concentrations of 50–400 mg L−1 over a wide range of temperatures (18–40 °C) and pH (5–10), making them promising candidates for use in β-CP degradation. Cell-free enzyme extracts have several advantages over living organisms in biodegradation, including strong environmental adaptability, higher degradative capabilities at lower concentrations, and not being subject to limitations such as rate of substrate uptake and catabolite repression (Nannipier and Bollag 1991). Several previous studies have attempted to degrade pyrethroids using enzymes. Yu and Fan (2003) showed that the crude enzyme from Alcaligenes sp. strain YF11 could be used under a wider range of environmental conditions than the strain from which it was derived. Zhang et al. (2010b) employed cell-free extracts from Clostridium sp. ZP3 to degrade fenpropathrin, suggesting that it could be used for rapid removal of pesticide residues from the surface of fruits and vegetables. Although numerous reports suggest that the key enzyme in the biodegradation of contaminants is intracellular (Maloney et al. 1993; Liang et al. 2005; Guo et al. 2009), several other reports suggest otherwise. For example, Niebisch et al. (2010) showed that decolorization of the dye reactive blue 220 by novel fungal isolate Lentinus crinitus is associated with an extracellular laccase, while Mazotto et al. (2010) treated feather waste with extracellular keratinases and gelatinases derived from Bacillus species. However, extracellular enzymes involved in the biodegradation of pyrethroids are scarce.
A better understanding of metabolic pathways and key enzymes therein is needed to explore the biodegradation potential of microbes in practical bioremediation processes and for developing synthetic biological technologies. The first step in pyrethroid degradation involves hydrolysis of the ester bond, which is thought to be catalyzed by a carboxylesterase (Sogorb and Vilanova 2002), releasing carboxylic acid and alcohol. Since the first isolation of a microbial carboxylesterase from Bacillus cereus SM3 by Maloney et al. (1993), several hydrolases from Aspergillus niger ZD11 (Liang et al. 2005), Sphingobium sp. JZ-2 (Guo et al. 2009), and metagenomes (Li et al. 2008; Fan et al. 2012) have been purified to homogeneity and characterized. In all cases, the reported enzymes are endoenzymes.
In this study, we identified P. aeruginosa strain GF31, isolated from contaminated soil near Nanning, China, and determined that it could use β-CP as a substrate. Surprisingly, in contrast to other known pyrethroid-degrading enzymes, the enzyme responsible for degradation was mainly localized to the extracellular areas of the bacterial cell. These findings indicated that strain GF31 may possess a distinct substrate utilization pattern and that the degrading enzyme has specific performance characteristics.
Materials and methods
Chemicals and media
β-CP (97.2 %), fenpropathrin (91.5 %), fenvalerate (95.3 %), and cyhalothrin (97.8 %) were obtained from Guangxi Plant Protection General Station (Nanning, China). 3-Phenoxybenzoic acid (3-PBA), 3-phenoxy-benzaldehyde (3-PBH), and 3-(2,2-dichloroethenyl)-2,2-dimethylcyclopropanecarboxylate (DCVA) were purchased from Sigma-Aldrich Chemical Co. (Shanghai, China). All other chemicals were of analytical grade. Beef extract-peptone medium contained beef extract (5.0 g L−1), peptone (10.0 g L−1), and NaCl (5.0 g L−1) (pH 7.0). Mineral salts medium (MM) contained K2HPO4 (1.5 g L−1), KH2PO4 (0.5 g L−1), NaCl (0.5 g L−1), and MgSO4 (0.2 g L−1) (pH 7.0). Solid medium plates were prepared by adding 1.5 % (w/v) agar. Pyrethroids were dissolved in acetonitrile to a stock solution concentration of 20,000 mg L−1 (w/v) and sterilized by membrane filtration (pore size 0.22 μm). Stock solution was added directly to the medium at appropriate volumes to achieve the desired final concentration, and the concentration of acetonitrile in the final degradation system was up to 0.25 %.
Identification of the pyrethroid-degrading strain
The degrading strain was isolated from pyrethroid-contaminated soil near Nanning, China, through enrichment culture (Li et al. 2009). The strain was identified by standard laboratory procedures and those described in Bergey’s manual of determinative bacteriology (Holt et al. 1994). The 16S rRNA gene was amplified by PCR using the following primers: (f) 5′-AACACATGCAAGTCGAACG-3′; (r) 5′-GGTTACCTTGTTACGACTT-3′. The obtained sequence was aligned to known sequences in the GenBank database by BLAST analysis. Phylogenetic analyses were performed using MEGA version 5.2 software, and distances were calculated using the Kimura two-parameter distance model. Unrooted trees were built using the neighbor-joining method.
Cultivation of the bacteria and preparation of crude enzymes
A seed culture of strain GF31 was cultivated in a 250-mL Erlenmeyer flask containing 30-mL beef extract-peptone medium at 30 °C in a rotary shaker at 120 rpm. After 18 h, at which the stationary phase was reached (S1), the culture was centrifuged at 4500×g for 10 min at 4 °C; then, the pellet was washed three times in MM and resuspended in MM to form the cell suspension for the biodegradation (Li et al. 2009).
Crude enzyme was prepared using an osmotic-shock-based method described previously (Neu and Heppel 1965), with some modifications. Briefly, the culture supernatant was extracted by ultrafiltration with protein concentration adjusted to about 0.5 mg mL−1 and used as the extracellular crude enzyme. The cells were washed in a 25 % (w/v) sucrose solution and then resuspended in ice-cold distilled water and shacked in water bath for periplasmic enzyme preparation. After that, the cells were resuspended in 0.01 mol L−1 Tris-HCl buffer (pH 7.5) and disrupted by ultrasonication on ice to obtain the crude intracellular enzyme preparation (S2).
Biodegradation of β-cypermethrin by strain GF31
Flasks containing 10 mL MM and 50 mg L−1 β-CP were inoculated with bacterial cell suspension to a final cell density of approximately OD600 = 0.4. An air-permeable silica gel plug was used to reduce volatilization of the reaction phase. Flasks were incubated at 30 °C on a shaker at 120 rpm. The degradation was performed at pH 7.0, and flasks were collected at regular intervals. Samples were acidified to pH 2 using 1 M HCl and then extracted with 10 mL ethyl acetate (Li et al. 2009).
The influence of temperature and pH on biodegradation was studied by single factor analysis using a temperature range of 20–50 °C and a pH range of 5–9. The effect of β-CP concentration (50–300 mg L−1) on degradation was also examined, and the kinetic parameters were determined from Lineweaver-Burk plots under a linear regression. The effects of using glucose or peptone (each at 1 g L−1) as an extra carbon source on β-CP-degrading activity of strain GF31 were also investigated. Flasks were prepared as described above, and blank controls without the bacteria were included in all experiments. All experiments were performed at least in triplicate.
The ability of GF31 to degrade other pyrethroids was also investigated under the conditions described above.
Biodegradation of β-cypermethrin by crude enzyme
A reaction mixture containing 2.8 mL of phosphate buffer (50 mM, pH 7.0), 50 mg L−1 β-CP, and 0.2 mL of crude extracellular enzyme from strain GF31 was incubated at 30 °C on a shaker at 120 rpm for 48 h. The reactions were stopped by adding 0.2 mL of 1 M HCl. Residual β-CP was extracted by adding 3.0 mL of ethyl acetate, and the enzyme activity was assayed by measuring the decrease in the amount of β-CP.
A series of reactions were performed from pH 5.0 to 9.0 and from 20 to 70 °C, to observe the effects on the percentage degradation yield. Residual β-CP was extracted as described above, and the optimal conditions for degradation of β-CP were then determined. An additional assay was then performed under these optimal conditions to measure degradation yields. All experiments were performed in triplicate, and controls lacking crude extracellular enzyme were included.
Kinetic parameters were determined by measuring the enzyme activity when initial β-CP concentrations were 20–200 mg L−1. Each initial velocity was determined at a point when no more than 10 % of the substrate had been consumed, so that the decrease in the substrate concentration remained linear over time. Initial reaction velocities measured at various substrate concentrations were fitted to the Lineweaver-Burk transformation of the Michaelis-Menten equation.
Soluble protein concentration was determined according to the Bradford method (Bradford 1976), using bovine serum albumin as a standard. One unit of enzyme activity was defined as the amount required to catalyze the consumption of 1 μmol of substrate per minute.
Extraction and determination of pyrethroids
Pyrethroids were extracted with ethyl acetate, and then, the extracts were passed through a 0.45-μm filter prior to analysis. Pyrethroids were quantified using an ULTIMATE 3000 high-performance liquid chromatography system (Idstein, Germany) equipped with an ultraviolet detector. Assays were conducted at room temperature using a LiChrospher 5-μm C18 column (250 × 4.6 mm). The mobile phase was 85:15 (v/v) acetonitrile and water. The sample injection volume was 20 μL, and the mobile phase was programmed at a flow rate of 1 mL min−1 and detected at a wavelength of 235 nm (Galera and Vidal 1996).
Isolation and identification of metabolites
The metabolites of β-CP, 3-PBA and DCVA, were detected by gas chromatography-mass spectrometry (SHIMADZU GCMS-QP2010 Plus, Kyoto, Japan). The samples were extracted with ethyl acetate following acidification to pH 2 with 1 M HCl and then filtered using a 0.45-μm membrane. The GC-MS system was equipped with an autosampler, a split/splitless capillary injection system, and a DB-5MS capillary column (30.0 m × 0.32 mm × 0.25 μm). Compounds in the samples were characterized by array detection from 50 to 450 nm (total scan). The operating conditions were an initial column temperature of 100 °C for 1 min, which was then ramped at 10 °C min−1 to 250 °C, and finally held at 250 °C for 20 min. The temperatures of the transfer line and ion trap were 250 and 200 °C, respectively, and the ionization energy was 70 eV. The inlet temperature was 220 °C, and a 1-μL sample was injected. The flow rate of the carrier gas (helium) was 3.0 mL min−1, and the split ratio was 10:1 (Tallur et al. 2008).
Results
Identification of strain GF31
Strain GF31 was characterized using morphological, physiological, and biochemical techniques, as well as 16S ribosomal DNA (rDNA) gene analysis. Strain GF31 is a Gram-negative, aerobic, rod-shaped, non-sporulating bacterium. Colonies were circular, smooth, convex, and semitransparent on bouillon culture medium agar plates. The medium was stained green by GF31 at the initial stage of culture but changed to red brown in the later stages. Strain GF31 exhibited catalase and oxidase activities and could grow in MM supplemented with d-glucose, fructose, lactin, and citric acid. Lysine decarboxylase, phenylalanine deaminase, and arginine dihydrolase tests were all negative, and rhamnose, starch, and o-nitrophenyl-β-d-galactoside did not support growth of strain GF31. Analysis of the partial 16S rDNA region of strain GF31 showed that it was most closely related to Pseudomonas species (100 % similarity to P. aeruginosa LMG 1242) (Fig. 1). The 1450-bp 16S rDNA gene sequence was deposited in the GenBank database under accession number KF432838.

Phylogenetic tree based on the 16S rRNA gene sequences of strain GF31 and related species. The GenBank accession number for each microorganism used in the analysis is shown after the species name. The scale bar indicates 0.005 substitutions per nucleotide position. Bootstrap values obtained with 1000 resamplings are indicated as percentages at all branches
Based on the morphological, physiological, and biochemical and 16S rDNA gene analyses, strain GF31 was identified as P. aeruginosa. P. aeruginosa strain GF31 was assigned culture collection number CGMCC 7173 in the China General Microbiological Culture Collection (CGMCC).
Biodegradation of pyrethroids by strain GF31
Single-factor tests were used to examine the effects of independent variables, including temperature, pH, and substrate concentration, on the biodegradation of β-CP. Figure 2a shows the effect of pH on β-CP degradation at day 4. Under acidic (pH 5.0) and alkaline (pH 9.0) conditions, less than 10 % degradation was observed. However, at neutral pH, the degradation yield rose to 33.1 %, with pH 7.0 determined as the optimal pH for degradation. The results indicate that the effect of pH on the degradation activity of strain GF31 is highly significant. As the temperature decreased below 30 °C, the percent degradation yield increased (Fig. 2a). In contrast, degradation yields decreased slowly at temperatures above 30 °C. The optimal temperature was 30 °C. As shown in Fig. 2b, strain GF31 could degrade 200 mg L−1 β-CP at a maximum rate of 25.1 μmol L−1 day−1. The degradation rate catalyzed by strain GF31 increased as the substrate concentration increased from 50 to 200 mg L−1. However, at concentrations higher than 200 mg L−1, the degradation rate did not increase. These results showed that the degradation reaction meets the Michaelis-Menten kinetics, with kinetic parameters of V max = 46.9 μmol L−1 day−1 and K m = 204.5 mg L−1, and the R 2 value was 0.9872, demonstrating that the experimental data are well correlated with the model. Similar to O. lupini DG-S-01 (Chen et al. 2011), with the addition of glucose or peptone, the removal of β-CP was highly effective, and the percent degradation yield reached 53.8 and 81.2 % within 7 days, respectively (Fig. 2c), while the percent degradation yield of β-CP in MM only reached 37.3 %. However, this result contrasts with previous findings describing that addition of other nutrients led to a lag phase followed by accelerated biodegradation. Our findings revealed that GF31 preferred to utilize β-CP even in a nutrient-rich environment. In non-inoculated controls, β-CP degradation was negligible. Strain GF31 also degraded other pyrethroids, albeit less efficiently (Table 1). This result indicates that strain GF31 can degrade a series of analogue compounds, although the chemical and structural properties of pyrethroids can influence degrading efficiency.

Degradation of β-CP by strain GF31. a Biodegradation of β-CP by strain GF31 at different pHs (black squares) and different temperatures (black circles) (4 days). b Biodegradation speed of β-CP by strain GF31 at different β-CP concentrations under optimum conditions. c Effect of glucose or peptone on β-CP degradation by strain GF31 under optimum conditions. Control (white diamonds), no other added carbon source or nutrients (black squares), glucose (black circles), peptone (black triangles). Values are the mean ± SD of three replicates
Biodegradation of β-cypermethrin by crude enzyme
As crude extracellular enzyme from strain GF31 degraded β-CP more efficiently than crude intracellular enzyme and periplasmic enzyme (Table 2), we used crude extracellular enzyme in the subsequent experiments. Other reports concerning enzymatic degradation of pyrethroid pesticides focus on the intracellular enzyme. However, compared with intracellular enzyme, extracellular enzyme is more easily obtained and purified. Some reports show that certain enzyme fractions do not contain any degradation activity unless grown in the presence of a pyrethroid (Wang et al. 2011; Zhang et al. 2010b). However, the results shown here indicate that β-CP is degraded rapidly by crude extracellular enzyme of strain GF31 without any preculture in the presence of pyrethroid pesticides, suggesting that the degrading enzyme can be constitutively expressed.
Crude extracellular enzyme degraded β-CP at pH 5–9, with an optimal pH of 7.0. The percent removal of β-CP was >30 % at all points within this range, and the pH activity profile of the enzyme was very flat (Fig. 3a). These findings indicated that the crude extracellular enzyme from strain GF31 has excellent catalytic activity over a wide pH range. Temperature is another important factor that significantly influences the degradation ability of the enzyme. β-CP was degraded by crude extracellular enzyme at temperatures from 20 to 70 °C. The percent degradation yield increased rapidly with increasing temperature and peaked at 60 °C. Activity began to decline at 70 °C; however, the percent degradation yield at 70 °C was still over half of that at the highest point (Fig. 3a). These findings indicated that the crude extracellular enzyme from strain GF31 had unexceptionable thermal stability. Compared with β-CP degradation with strain GF31, degradation with the isolated fractions displayed wider pH tolerance and a higher optimal temperature, because biodegradation with cell-free enzymes excludes the influences and limitations brought by bacteria growth and metabolism. This is an important feature for bioremediation in a variable environment. Crude extracellular enzyme from strain GF31 degraded β-CP efficiently under optimal conditions to the point of removing it completely by day 7 (Fig. 3b). Kinetic parameters were determined by measuring the initial reaction velocities at various concentrations of the substrates (S3). The V max and K m for the crude enzyme were 176.7 μmol L−1 day−1 and 27.5 mg L−1, respectively. The K m for the crude enzyme was lower than that of the corresponding whole bacterial cell, indicating a stronger affinity for the substrate. In addition, crude extracellular enzyme had excellent storage stability. Relative β-CP degradation of crude extracellular enzyme was 51.9 % after storage for 20 days at 25 °C (Fig. 4).

Enzymatic degradation capacity of β-CP. a Enzymatic degradation capacity of extracellular crude enzyme at different pHs (black squares) and different temperatures (black circles) (2 days). b Enzymatic degradation capacity of β-CP under optimal conditions. Control (white diamonds), extracellular crude enzyme (black squares). Values are the mean ± SD of three replicates

The storage stability of the extracellular crude enzyme fraction at 25 °C
Isolation and identification of metabolites
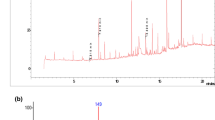
The metabolites produced following degradation of β-CP by strain GF31 and its crude extracellular enzyme extract were analyzed by GC-MS. The mass spectra results revealed the presence of several compounds, which corresponded well with DCVA (Fig. 5) and 3-PBA (Fig. 6). The retention times of the compounds were 13.48 and 20.99 min, respectively. It is generally believed that pyrethroid metabolism mainly occurs through hydrolysis and oxidation (Sogorb and Vilanova 2002). Our results indicated that β-CP is degraded through an initial hydrolysis of its ester linkage, yielding DCVA and cyano-3-phenoxybenzylalcohol, with the latter being rapidly converted into 3-PBH (Tallur et al. 2008). Moreover, we determined that 3-PBH can be rapidly converted into 3-PBA by the bacteria (data not shown), which explains why 3-PBH was not obviously detectable from the reaction system. The hydrolysis of the ester linkage destroys the insecticidal activity of β-CP, leading to its detoxification.

Mass spectrum of DCVA

Mass spectrum of 3-phenoxybenzoic acid
The curves showing the appearance and disappearance of DCVA and 3-PBA shared the same trend. The highest DCVA and 3-PBA concentrations were observed within the first 4 days of the degradation process in the presence of cells, with their respective concentrations increasing from days 1 to 4. After 4 days, the concentrations of DCVA and 3-PBA were distinctly decreased (Fig. 7). In the presence of extracellular crude enzyme, the highest DCVA and 3-PBA concentrations were observed within 3 days and then gradually declined (Fig. 8). These results indicate that strain GF31 and its crude extracellular enzyme fraction can degrade both β-CP and its degradation products.

Time course of the appearance and disappearance of substrate and products from the degradation of β-CP in the presence of strain GF31. β-CP (black squares), 3-PBA (black circles), DCVA (black triangles). Values are the mean ± SD of three replicates

Time course of the appearance and disappearance of substrate and products from the degradation of β-CP in the presence of the extracellular enzyme fraction of strain GF31. β-CP (black squares), 3-PBA (black circles), DCVA (black triangles). Values are the mean ± SD of three replicates
Discussion
Because of their extensive and long-term use in both agricultural and domestic situations, a substantial amount of residual pyrethroids has accumulated in rivers and soil and can even be found on the surface of fruits and vegetables. P. aeruginosa strain GF31, isolated from contaminated soil and identified in this study, can degrade several kinds of pyrethroids in addition to β-CP. It can also further degrade the catabolite 3-PBA to a similar extent as P. aeruginosa strain CH7 (Zhang et al. 2011), suggesting that these two species share certain characteristics. In addition, the decrease of another metabolite, DCVA, was observed during the degradation process. Thus, use of strain GF31 can be beneficial for the complete mineralization of β-CP, as 3-PBA, classified as an endocrine-disrupting chemical owing to its antiestrogenic activity (Meeker et al. 2009), has a higher mobility than its parent compounds. Other than Ochrobactrum sp. DG-S-01 (Chen et al. 2011), P. aeruginosa CH7 (Zhang et al. 2011), Stenotrophomonas sp. ZS-S-01 (Chen et al. 2010), and Streptomyces sp. HU-S-01 (Lin et al. 2011), few other microorganisms have been identified as having this PBA-degrading ability.
For hydrophobic organic compounds (HOCs) such as pyrethroids, direct adherence to the bacterial cell surface is the first and key step in the process of degrading hydrophobic organic pollutants by microbes (Zita and Hermansson 1997). Many reports have shown that the degrading efficiency can be improved by changing the hydrophobicity of the substance or the microorganism. Zhang et al. (2010a) attributed the better β-CP degradation ability of strain JC1 to its higher cell surface hydrophobicity than strain JCN13, even though they are the same species. Adding surfactants is also a common strategy to modify hydrophobicity (Lanzon and Brown 2013). In cases where the microorganism itself can secrete extracellular active enzymes, the conversion of the HOCs into hydrophilic small molecular compounds prior to sorption could enhance the bioavailability of HOCs. For example, Kwak et al. (2013) generated an engineered strain for biodegradation of tolclofos-methyl that could secrete extracellular organophosphorus hydrolase. To date, the only crude enzymes described for the degradation of pyrethroids, such as those from Clostridium sp. ZP3 and Alcaligenes sp. YF11, are endoenzymes, while the degrading enzymes obtained by purification, such as those from B. cereus SM3, A. niger ZD11, and Sphingobium sp. JZ-2, are also intracellular. Therefore, GF31 is the first reported example of a bacterial strain that can secrete extracellular hydrolase for pyrethroid degradation. In addition, the secreted enzyme can hydrolyze β-CP into DCVA and 3-PBA, which are then further degraded by other microorganisms. This degradation pathway was explored to supply a new theoretical basis for the bioremediation of organic pollutants such as pyrethroids in the environment.
The catalytic characteristics of an enzyme are the most important factors influencing its practical application. The optimal temperature for the activity of the crude extracellular enzyme extract from GF31 was 60 °C, much higher than temperatures reported for Clostridium sp. ZP3 (35 °C) and Alcaligenes sp. YF11 (32.5 °C). Moreover, the crude enzyme from strain GF31 showed good storage stability, maintaining more than 50 % initial activity after storage at 25 °C for 20 days. These findings mean that the crude extracellular enzyme can theoretically maintain activity for environmental bioremediation for a long period. This study also suggested that the crude extracellular enzyme from strain GF31 had very good thermal stability. Importantly, thermostable enzymes are often used in solvents and detergents, giving these enzymes considerable potential for applications in industry and household detergent manufacturing. For example, Xu et al. (2008) reported the use of cell-free extracts for removal of pesticides on fruits and vegetables. Finally, the crude extracellular enzyme from GF31 displayed a good range of pH tolerance, with no appreciable change in the percent degradation yield between pH 5 and pH 9. This feature is helpful when dealing with the changeable environments often encountered during bioremediation.
In summary, P. aeruginosa strain GF31 can degrade β-CP with high efficiency and further degrade the resulting metabolites DCVA and 3-PBA. The degradation results from the action of an extracellular hydrolase secreted by GF31, and a crude extracellular enzyme extract shows good thermal stability and pH tolerance, making it a promising target for industrial applications. In addition, relative to the intracellular enzymes, which require complex extraction protocols and present issues in terms of easy deactivation and low production, extracellular enzymes are more useful for industrial production and applications. The degrading enzyme reported here is not a monooxygenase or a peroxidase, based on metabolite analysis, and is significantly different from the previously reported carboxylesterase, as it has a higher thermal stability and is located extracellularly. The above findings suggest that the unidentified enzyme from GF31 can degrade pyrethroids and should be investigated further.
References
Ahmad M, Hussain I, Khan A, Najib ur R (2009) Deleterious effects of cypermethrin on semen characteristics and testes of dwarf goats (Capra hircus). Exp Toxicol Pathol 61(4):339–346
Bradford MM (1976) A rapid and sensitive method for the quantitation of microgram quantities of protein. Anal Biochem 72(1–2):248–254
Chen SH, Yang L, Hu M, Liu J (2010) Biodegradation of fenvalerate and 3-phenoxybenzoic acid by a novel Stenotrophomonas sp. strain ZS-S-01 and its use in bioremediation of contaminated soils. Appl Microbiol Biotechnol 90(2):755–767
Chen SH, Hu M, Liu J, Zhong G, Yang L, Rizwan-ul-Haq M, Han H (2011) Biodegradation of beta-cypermethrin and 3-phenoxybenzoic acid by a novel Ochrobactrum lupini DG-S-01. J Hazard Mater 187(1–3):433–440
Chen SH, Dong YH, Chang C, Deng Y, Zhang XF, Zhong G, Song H, Hu M, Zhang LH (2013) Characterization of a novel cyfluthrin-degrading bacterial strain Brevibacterium aureum and its biochemical degradation pathway. Bioresour Technol 132:16–23
Fan XJ, Liu XL, Huang R, Liu YH (2012) Identification and characterization of a novel thermostable pyrethroid-hydrolyzing enzyme isolated through metagenomic approach. Microb Cell Factories 11(1):33–43
Galera M, Vidal JLM (1996) Determination of cypermethrin, fenvalerate and cis-and trans-permethrin in soil and groundwater by high-performance liquid chromatography using partial least-squares regression. J Chromatogr A 727(1):39–46
Guo P, Wang B, Hang B et al (2009) Pyrethroid-degrading Sphingobium sp. JZ-2 and the purification and characterization of a novel pyrethroid hydrolase. Int Biodeterior Biodegrad 63(8):1107–1112
Holt JG, Krieg NR, Sneath PHA, Staley JT, Williams ST (1994) Bergey’s manual of determinative bacteriology. Williams and Wilkins, USA
Kwak YY, Rhee IK, Shin JH (2013) Expression pattern of recombinant organophosphorus hydrolase from Flavobacterium sp. ATCC 27551 in Escherichia coli. Appl Microbiol Biotechnol 97:8097–8105
Lanzon JB, Brown DG (2013) Partitioning of phenanthrene into surfactant hemi-micelles on the bacterial cell surface and implications for surfactant-enhanced biodegradation. Water Res 47(13):4612–4620
Li G, Wang K, Liu YH (2008) Molecular cloning and characterization of a novel pyrethroid-hydrolyzing esterase originating from the metagenome. Microb Cell Factories 7(1):38–47
Li QY, Gu BQ, Liu YY, Zhou MZ, Li C, Qin YM, Zhong SJ (2009) Isolation, identification and characteristics of a cypermethrin-degrading bacterium GF31. Microbiology 36(9):1334–1339 (in Chinese)
Li H, Mehler TW, Lydy MJ, You J (2011) Occurrence and distribution of sediment-associated insecticides in urban waterways in the Pearl River Delta, China. Chemosphere 82(10):1373–1379
Liang WQ, Wang ZY, Li H, Wu PC, Hu JM, Luo N, Cao LX, Liu YH (2005) Purification and characterization of a novel pyrethroid hydrolase from Aspergillus niger ZD11. J Agric Food Chem 53(19):7415–7420
Lin QS, Chen SH, Hu MY, Rizwan-ul-Haq M, Yang L, Li H (2011) Biodegradation of cypermethrin by a newly isolated actinomycetes HU-S-01 from wastewater sludge. Int J Environ Sci Technol 8(1):45–56
Liu P, Liu Y, Liu Q, Liu J (2010) Photodegradation mechanism of deltamethrin and fenvalerate. J Environ Sci 22(7):1123–1128
Maloney SE, Maule A, Smith AR (1993) Purification and preliminary characterization of permethrinase from a pyrethroid-transforming strain of Bacillus cereus. Appl Environ Microbiol 59(7):2007–2013
Mazotto AM, de Melo ACN, Macrae A et al (2010) Biodegradation of feather waste by extracellular keratinases and gelatinases from Bacillus spp. World J Microbiol Biotechnol 27(6):1355–1365
Meeker JD, Barr DB, Hauser R (2009) Pyrethroid insecticide metabolites are associated with serum hormone levels in adult men. Reprod Toxicol 27(2):155–160
Mehler WT, Li H, Lydy MJ, You J (2011) Identifying the causes of sediment-associated toxicity in urban waterways of the Pearl River Delta, China. Environ Sci Technol 45(5):1812–1819
Nannipier P, Bollag JM (1991) Use of enzymes to detoxify pesticide-contaminated soils and waters. J Environ Qual 20:510–517
Neu HC, Heppel LA (1965) The release of enzymes from Escherichia coli by osmotic shock and during the formation of spheroplasts. J Biol Chem 204(9):3685–3692
Niebisch CH, Malinowski AK, Schadeck R et al (2010) Decolorization and biodegradation of reactive blue 220 textile dye by Lentinus crinitus extracellular extract. J Hazard Mater 180(1–3):316–322
Saikia N, Gopal M (2004) Biodegradation of α-cyfluthrin by fungi. J Agric Food Chem 52(5):1220–1223
Shafer TJ, Meyer DA, Crofton KM (2005) Developmental neurotoxicity of pyrethroid insecticides: critical review and future research needs. Environ Health Perspect 113(2):123–136
Sogorb MA, Vilanova E (2002) Enzymes involved in the detoxification of organophosphorus, carbamate and pyrethroid insecticides through hydrolysis. Toxicol Lett 128:215–228
Tallur P, Megadi V, Ninnekar H (2008) Biodegradation of cypermethrin by micrococcus sp. strain CPN 1. Biodegradation 19(1):77–82
Thouand G (2014) Biodegradability assessments of organic substances and polymers. Environ Sci Pollut Res 21:9443–9444
Wang BZ, Ma Y, Zhou WY et al (2011) Biodegradation of synthetic pyrethroids by Ochrobactrum tritici strain pyd-1. World J Microbiol Biotechnol 27(10):2315–2324
Wolansky MJ, Harrill JA (2008) Neurobehavioral toxicology of pyrethroid insecticides in adult animals: a critical review. Neurotoxicol Teratol 30(2):55–78
Xu G, Zheng W, Li Y et al (2008) Biodegradation of chlorpyrifos and 3,5,6-trichloro-2-pyridinol by a newly isolated Paracoccus sp. strain TRP. Int Biodeterior Biodegrad 62(1):51–56
Yu Y, Fan D (2003) Preliminary study of an enzyme extracted from Alcaligenes sp. strain YF11 capable of degrading pesticides. Bull Environ Contam Toxicol 70(2):367–371
Zhang C, Jia L, Wang S, Qu J, Li K, Xu L, Shi Y, Yan Y (2010a) Biodegradation of beta-cypermethrin by two Serratia spp. with different cell surface hydrophobicity. Bioresour Technol 101(10):3423–3429
Zhang SB, Yin L, Liu Y, Zhang D, Luo X, Je C, Cheng F, Dai J (2010b) Cometabolic biotransformation of fenpropathrin by Clostridium species strain ZP3. Biodegradation 22(5):869–875
Zhang C, Wang S, Yan Y (2011) Isomerization and biodegradation of beta-cypermethrin by Pseudomonas aeruginosa CH7 with biosurfactant production. Bioresour Technol 102(14):7139–7146
Zita A, Hermansson M (1997) Effects of bacterial cell surface structures and hydrophobicity on attachment to activated sludge flocs. Appl Environ Microbiol 63:1168–1170
Acknowledgments
This work was support in part by the National Natural Science Foundation of China (No. 51108098), the Graduate Student Innovation Projects of Guangxi (No. GXU11T31090), and the Scientific Research Foundation of Guangxi University (No. XJZ120278).
Conflict of interest
The authors declare that they have no conflict of interest.
Author information
Authors and Affiliations
Corresponding author
Additional information
Responsible editor: Angeles Blanco
Rights and permissions
About this article

Cite this article
Tang, A., Wang, B., Liu, Y. et al. Biodegradation and extracellular enzymatic activities of Pseudomonas aeruginosa strain GF31 on β-cypermethrin. Environ Sci Pollut Res 22, 13049–13057 (2015). https://doi.org/10.1007/s11356-015-4545-0
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s11356-015-4545-0