
Abstract
The effects of levofloxacin (LEV) on Microcystis flos-aquae and its mechanism were investigated by determining the responses of some parameters of M. flos-aquae to LEV stress, including growth inhibition ratio, chlorophyll a content, superoxide dismutase (SOD) and catalase (CAT) activities, malondialdehyde (MDA) content, F v/F 0 and F v/F m, etc. The results indicated that LEV at 0.001–0.1 μg L−1 could stimulate the growth of M. flos-aquae and increase the chlorophyll a content but did not induce a significant increase in the activity of antioxidant enzymes (SOD and CAT) and the content of MDA. When the LEV concentration exceeds 10 μg L−1, the growth of M. flos-aquae could be significantly inhibited (the highest inhibition ratio can be up to 88.38 % at 100 μg L−1) and chlorophyll a content, SOD and CAT activities, and MDA content also significantly decreased in a concentration-dependent manner, indicating that high concentrations of LEV caused a severe oxidative stress on algal cells, resulting in a large number of reactive oxygen species produced in algal cells and thereby inhibiting the growth of algae. At the same time, the F v/F m and F v/F 0 values of M. flos-aquae decreased significantly with both exposure time and increasing test concentration of LEV, showing that the process of photosynthesis was inhibited.
Similar content being viewed by others


Explore related subjects
Discover the latest articles, news and stories from top researchers in related subjects.Avoid common mistakes on your manuscript.
Introduction
In recent years, antibiotics have been widely used in medicine and health, aquaculture industry, food processing, poultry farming, etc. In China, 15,770 tons of antibiotics were used as non-prescription therapeutics in 2004 (Richardson et al. 2005). Wise (2002) calculated that antibiotic consumption has been estimated worldwide to lie between 100,000 and 200,000 tons per annum, among which about 40,000 tons were used in China. Because antibiotics have short half-lives in living organisms and low metabolic rate, about 70–80 % of antibiotics are excreted by organisms in their parent form and then enter the aquatic ecosystem through many ways. More than 50 kinds of antibiotics have been detected in various aquatic environments, including sewage, surface, ground, and drinking waters, and the detected concentration was in the range of nanograms per liter to micrograms per liter (Tan et al. 2007; Ye et al. 2007). Previous studies have shown that macrolide and sulfonamide antibiotics had the highest residue levels, followed by quinolone antibiotics (Lalumera et al. 2004; Yang and Carlson 2004; Santos et al. 2010). Therefore, the potential hazard of antibiotics upon aquatic ecosystems has attracted increasing attention among people.
Quinolone antibiotics have been widely used in human and animal disease therapeutics as well as agricultural industry because of their very broad spectrum antimicrobial activity and high efficacy. Their environmental behavior and ecological effects are of increasing concern. Levofloxacin (LEV) is the levo isomer of ofloxacin, one of the representatives of quinolone antibacterial drugs. It kills bacteria by inhibiting key bacterial enzymes (DNA gyrase and topoisomerase IV) involved in unwinding the DNA helix for replication and transcription (Robinson et al. 2005). Robinson et al. (2005) investigated the toxicity of levofloxacin to five aquatic organisms but only determined the EC50 value. Pan et al. (2009) reported the effects of levofloxacin hydrochloride (LH) on the photosystem II activity and heterogeneity of Synechocystis sp.; the results suggested that O2 evolution and the photosystem II (PSII) activity were clearly inhibited by LH. These studies showed that LEV has adverse effects on aquatic organisms.
Recently, the frequent outbreak of cyanobacterial blooms has become a ubiquitous phenomenon in freshwater ecosystems, leading to serious harm to fishery, ecological environment, and human health. Cyanobacterial blooms have become one of the important environmental problems. Formation of cyanobacterial blooms is normally regulated by various factors, including eutrophication, light intensity, trace metals, temperature, and salinity (Jiang et al. 2008; Davis et al. 2009). However, the correlation between the residue of antibiotics in water and the outbreak of algal blooms has not been reported. Thus, in this paper, we investigated the effects of LEV on growth in a cyanobacterium (Microcystis flos-aquae). M. flos-aquae is a common freshwater alga that causes water bloom. Also, physiological and biochemical parameters including chlorophyll a content, superoxide dismutase (SOD) and catalase (CAT) activities, malondialdehyde (MDA) content, F v/F 0, and F v/F m were analyzed. The objective of the present work is to study the effects and the physiological and biochemical mechanisms of antibiotics to a cyanobacterium. Results from this study will help us evaluate the effects of antibiotic residues to the frequent outbreak and die out of cyanobacterial blooms.
Material and methods
The algae material and culture condition
M. flos-aquae (FACHB-1028) was provided by the Freshwater Algae Culture Collection of the Institute of Hydrobiology (FACHB-Collection), Wuhan City, China. The algae were grown in 250-mL Erlenmeyer flasks containing 100 mL of BG11 medium, cultivated at 25 ± 1 °C and 50 μmol photons m−2 s−1 illumination (cool white fluorescent tube) with a 12-h light/dark cycle. In order to reduce any effect caused by minor difference in photon irradiance, the flasks were shaken manually three times each day and rearranged randomly.
Antibiotic treatment
LEV was purchased from Beijing Jorferin Bio-Technology Co., Ltd, Beijing, China, with purity >98 %. The stock solution was prepared with sterilized water and then diluted to various test concentrations before use. When the algae were at the logarithmic growth phase, 1 mL of sterilized water or LEV in various concentrations was added into the algal medium to make final concentrations of 0, 0.001, 0.01, 0.1, 1, 10, 40, 70, and 100 μg L−1. The algal medium with 0 μg L−1 was used as the control. Each test concentration was replicated three times, and all operations were carried out under sterile conditions to avoid contamination from bacteria. All cultures were cultivated according to the culture condition described above.
Algal growth rate
Cells were counted everyday using a hemocytometer under a microscope (Olympus CX41, Japan). Meanwhile, the optical density OD680 was also measured everyday using a spectrophotometer (UV1800, Shimadzu, Japan). The linear relationship was built up between cell density and optical density, and the result showed that the relevant index R 2 > 0.99, which is well in agreement with the previous report (Ge et al. 2010). The growth inhibition ratio was calculated as follows:
where N is the cell count of the treated group, N 0 is the cell count of the control group, and IR is the inhibition ratio.
Chlorophyll a content and photosynthetic activity
Chlorophyll a content and fluorescence parameters were determined using a pulse amplitude-modulated fluorometer (Phyto-PAM Walz, Effeltrich, Germany). In this study, chlorophyll a was recorded everyday, and fluorescence parameters were measured every other day. Reviews of fluorescence measurements are given by Schreiber et al. (1994). The minimal fluorescence yield F 0 and the maximal fluorescence yield F m were measured after the samples were dark-adapted for at least 15 min. As a measure of the chlorophyll a concentration, the fluorescence was at an irradiance of 32 μmol photons m−2 s−1 PAR. Based on such measurements, the photosynthetic activity parameters can be formulated mathematically as (Rohacek and Bartak 1999)
Analysis of antioxidant responses
The algal cells from 80 mL of culture medium were harvested after 7 days of exposure to LEV and centrifuged at 5,000 g at 4 °C for 15 min, after which the supernatant was discarded. Collected algae were washed twice with 5 mL of buffer (50 mM potassium phosphate and 150 mM potassium chloride, pH 7.5). The algae were then transferred into 5 mL of buffer solution, ground at 4 °C with quartz powder, and then centrifuged at 12,000 g at 4 °C for 10 min (Geoffroy et al. 2003). The supernatant was used for enzyme activity assays.
The SOD activity was determined by the nitro blue tetrazolium (NBT) photochemical reduction method (Giannoplitis and Ries 1977). One unit of SOD activity was defined as the amount of enzyme that caused a 50 % decrease of the SOD-inhibited NBT reduction. The activity of SOD was expressed as units per cell. The activity of CAT was measured by the method of Goth (1991). One unit of CAT activity was defined as the amount of enzyme which degraded 1 mmol H2O2 per minute at 37 °C. The activity of CAT was expressed as units per cell. The content of MDA was determined by the thiobarbituric acid method as described by Hegedüs et al. (2001) and expressed as nanomoles per cell.
Statistical analysis
All data presented were expressed as mean ± standard deviation. Statistical analysis was performed using SPSS statistical package version 17.0. One-way analysis of variance (ANOVA) followed by the least significant difference test and Tukey's test was used to establish differences between the control group and treatments. The difference from the control group was considered significant at P < 0.05 or very significant at P < 0.01. All figures were produced using Origin 7.5.
Results
Algal growth rate
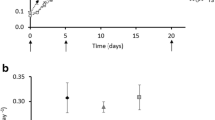
The effects of LEV on the growth curve and inhibition ratio of M. flos-aquae are depicted in Fig. 1a, b. As shown in Fig. 1, at 0.001–0.1 μg L−1 of LEV, the inhibition ratio of M. flos-aquae was negative, the growth was significantly stimulated (P < 0.05), and the promotion began to decrease with further increases in the concentration of LEV but still higher than the control. After 7 days of exposure, the average increments over the control values were 4.99, 3.13, and 1.52 %, respectively. The growth of M. flos-aquae slightly increased at the concentration of 1 μg L−1, but not obvious (P > 0.05). At 10 μg L−1 of LEV, the growth of M. flos-aquae increased firstly and then decreased. A statistically significant inhibition of growth occurred when the concentrations of LEV were equal to and higher than 40 μg L−1; the inhibition ratio increased in a concentration-dependent manner. At the concentration range of 10–100 μg L−1, the inhibition ratios were 11.92, 81.57, 83.22, and 88.38 % at 7 days of exposure, respectively. On the whole, the stress on M. flos-aquae by LEV increased with increasing concentration of LEV, and the inhibition presented a concentration-dependent trend.

Effects of levofloxacin on the growth curve (a) and inhibition ratio (b) of M. flos-aquae. Data shown are mean values±SE (n = 3). Error bars indicate the standard deviation
Chlorophyll a content
The effect of different concentrations of LEV on the chlorophyll a content of M. flos-aquae is depicted in Fig. 2. The content of chlorophyll a showed a significant increase (P < 0.01) compared with the control during 1–7 days of exposure to the LEV concentration of 0.001 μg L−1. There was a slight increase of chlorophyll a content by LEV at 0.01–1 μg L−1; the degree of increase gradually declined with time delays but remained higher than the control. The chlorophyll a content in response to LEV was inhibited at the test concentration of 10 μg L−1 after 6 days of exposure, showing a significant decrease (P < 0.01) compared with the control. A statistically significant inhibition of chlorophyll a content occurred when the concentration of LEV was equal to and higher than 40 μg L−1, and the inhibition ratio went up to 73.07, 76.28, and 85.6 % after 7 days of exposure, respectively.

Effects of levofloxacin on the chlorophyll a content of M. flos-aquae. Data shown are mean values±SE (n = 3). Error bars indicate the standard deviation
Photosynthetic activity
The F v/F m and F v/F 0 ratios of photosystem II were determined to assess the impact of LEV on the maximum photosynthetic capacity and the potential vitality of M. flos-aquae (Rohacek and Bartak 1999). As shown in Fig. 3a, b, the F v/F m and F v/F 0 ratios increased slightly with low LEV concentrations (≤1 μg L−1) during 1–7 days of exposure, only showing a significant increase at the third day (P < 0.01). The F v/F m and F v/F 0 ratios of M. flos-aquae showed an increase of 5.72 and 12.57 % relative to the control at the test concentration of 0.001 μg L−1 after 7 days of exposure. At a concentration range of 40–100 μg L−1, the value of F v/F m and F v/F 0 decreased sharply after 3 days of exposure. The F v/F m and F v/F 0 ratios in 10 μg L−1 LEV exposure increased firstly and then decreased, showing a significant decrease below the control after 5 days of exposure (P < 0.01).

Effects of levofloxacin on F v/F m (a) and F v/F 0 (b) of M. flos-aquae. Data shown are mean values±SE (n = 3). Error bars indicate the standard deviation
The activities of CAT and SOD
To better understand the biochemical basis of resistance in M. flos-aquae caused by LEV exposure, SOD and CAT activities were measured, and the results are displayed in Fig. 4. The CAT activity was induced after M. flos-aquae was exposed to different concentrations of LEV (Fig. 4a) and showed a dose-dependent relationship. There were significant increases in the CAT activity by LEV at higher concentrations (10–100 μg L−1), which were 5.47, 7.65, 25.09, and 29.44 times the control, respectively. As depicted in Fig. 4b, the SOD activity showed no significant difference with the control at low levels (0.001–10 μg L−1) of LEV exposure, whereas LEV at high levels (40–100 μg L−1) could significantly (P < 0.01) stimulate the SOD activity, which increased to 1.32, 2.65, and 3.07 times the control, respectively.

Effects of levofloxacin on SOD (a) and CAT (b) activities of M. flos-aquae. Data shown are mean values±SE (n = 3).*P < 0.05 (statistically significant difference), **P < 0.01 (statistical significance), when compared to the control (0 μg L−1). Error bars indicate the standard deviation
MDA content
In the present study, the effect of LEV on the MDA content of M. flos-aquae was shown in Fig. 5. The MDA content of each treatment group was stimulated in almost all treatments to LEV with different concentrations. The MDA content increased with increasing LEV concentrations, that is, increased lipid peroxidation. Exposure of LEV at a lower concentration (<1 μg L−1) augmented the MDA content slightly. However, significant increments in MDA level were observed at 10 μg L−1 and higher concentrations, showing a dose-dependent trend. When the concentration of LEV was 100 μg L−1, the MDA content was 3.84 times the control.

Effects of levofloxacin on the MDA content of M. flos-aquae. Data shown are mean values±SE (n = 3).*P < 0.05 (statistically significant difference), **P < 0.01 (statistical significance), when compared to the control (0 μg L−1). Error bars indicate the standard deviation
Discussion
Effects of LEV on the growth of M. flos-aquae
The present study indicated that LEV had a dual effect (promotion and inhibition) on the growth of M. flos-aquae. The growth was promoted by LEV at lower concentrations whereas inhibited at higher concentrations. Currently, this “low-promoting and high-repressing” phenomenon has been reported. Florfenicol was found to stimulate marine diatom Skeletonema costatum growth at concentrations of 0.5, 1.0, and 2.0 mg L−1 and significantly inhibit algal growth higher than 2.0 mg L−1 (Liu et al. 2012). Tetracycline at 0.5–10 mg L−1 could stimulate seed germination, cell mitotic division, and growth of wheat seedlings. However, tetracycline at high concentrations (10–300 mg L−1) could significantly inhibit these parameters in a concentration-dependent manner (Xie et al. 2011). LEV at lower concentrations (≤1 μg L−1) could promote the growth of M. flos-aquae, meanwhile the chlorophyll a content was also increased; this may result from certain enzymes involved in some physiological and biochemical reactions that were induced by the stress of LEV at certain concentrations. Moreover, it is possible that the algae may partially degrade LEV and absorbed LEV as nutrients instead of toxic xenobiotics (Yue et al. 2006). However, the precise mechanism remains to be investigated. At higher concentrations, LEV had inhibitory effects on the growth of M. flos-aquae (a possible reason is that the concentration of LEV exceeds the tolerance limit of algal cells), the cell structure begins to crack and disintegrate, and algal cell growth is in the state of zero growth or negative growth, which resulted in growth that cannot be fully restored (Nie et al. 2007).
Effects of LEV on the chlorophyll a content and photosynthetic activity of M. flos-aquae
Plant photosynthesis is the process of converting solar energy into chemical energy. It is known that chlorophylls play a key role in all aspects of primary photosynthesis, including light harvesting, energy transfer, and light energy conversion. Chlorophyll a content can be used to estimate the primary productivity and also is an important symbol of vitality of phytoplankton. Under a certain environmental stress, the chlorophyll a content of microalgae will also be affected (Liu et al. 2011; Qian et al. 2012). At lower concentrations of LEV, the synthesis of chlorophyll a was stimulated, which may have promoted the photosynthesis of algae. However, when LEV exceeded a certain range, which is over the self-adjustment range of algae, the inhibitory effects gradually increased with the increase of LEV dose, algal cell structure was damaged, algae liquid turned white, and chlorophyll a content decreased, affecting the light harvesting and finally may lead to the photosynthesis pathway being blocked. The possible reasons for the decrease of chlorophyll a content may be as follows: Firstly, antibiotic stress caused thylakoid membrane disintegration, which resulted in the loss of chlorophylls from the tissue (Gao et al. 2007). Secondly, the accumulation of intracellular reactive oxygen species caused the cell structure to be damaged and the chlorophyll synthesis to be blocked (Geoffroy et al. 2003). Thirdly, antibiotics directly combined with some ingredients of algae, which inhibited the synthesis of the light-harvesting chlorophyll a/b protein complex and resulted in reduced the efficiency of energy transformation (Alberte et al. 1981).
Chlorophyll fluorescence is closely linked with photosynthesis processes; it can not only reflect light absorption, excited energy transfer, and photochemical reactions of photosynthesis primary reaction process but is also connected with electron transfer, the establishment of proton gradient, ATP synthesis, and CO2 fixation. Thus, the chlorophyll fluorescence parameters can reflect changes in photosynthesis (Han et al. 2003). Adversity stress affects the photosynthesis of algae. The maximum quantum yield (F v/F m) indicates the maximum photochemical efficiency of PSII. The F v/F 0 ratio is a useful parameter in assessing the potential photosynthetic activity of PSII (Krause 1988). Under normal conditions, F v/F m and F v/F 0 remained relatively stable, freed from the influence of species and growing conditions, but changed significantly under stress. F v/F m and F v/F 0 decreased which meant that plants were photoinhibited (Razinger et al. 2007; Xing et al. 2010). In this study, the F v/F m and F v/F 0 ratios of M. flos-aquae increased slightly with low LEV concentrations (≤ 1 μg L−1) during 1–7 days of exposure, only showing a significant increase at the third day (P < 0.01). The F v/F m and F v/F 0 ratios were induced at the test concentration of 0.001 μg L−1 after 7 days of exposure. The results suggested that the low concentrations of LEV improved the maximum photochemical efficiency and the potential photosynthetic activity of PSII. The F v/F m and F v/F 0 ratios increased and then decreased at the concentration of 10 μg L−1, may be because M. flos-aquae rapidly started the synthesis of chlorophyll a to resist environmental stress during 1–3 days of exposure to LEV, increased the cell density, and accelerated the process of photosynthesis, but the inhibitory effect surpassed the defense effect with time delay; therefore, the F v/F m and F v/F 0 ratios started to decrease significantly after 5 days of exposure. At higher concentrations (40–100 μg L−1), F v/F m and F v/F 0 were significantly inhibited, which implied that the primary light energy conversion efficiency of PSII was reduced, damaged the potential active center, blocked the primary reaction process of photosynthesis, and finally inhibited the growth of algae. This may be ascribed to higher concentrations of LEV that interrupted the photosynthetic electron transport between Q A and Q B, thus forming more Q B-non-reducing PSII reaction centers, which resulted in the reduction of PSII reaction center oxygen evolution (Leu et al. 2002; Liang et al. 2006).
Effects of LEV on CAT and SOD activities and MDA content of M. flos-aquae
Under normal growth conditions, the generation and scavenging of reactive oxygen species (ROS) in plants are maintained in a dynamic equilibrium level, but under adverse conditions, this balance will be broken, which results in a large number of ROS generation and accumulation, triggers damaging effects to the membrane system of the organism, and finally inhibits growth (Tang and Li 2000). SOD and CAT are important enzymes associated with antioxidative stress in plants. SOD is an important antioxidative enzyme with free radicals as substrate, which can convert O2− to H2O2 and O2, thereby preventing the generation of superoxide anion radicals. CAT can catalyze H2O2 into H2O and O2 in order to alleviate the oxidative damage caused by H2O2. SOD and CAT make up the oxidation and antioxidant defense system which is indispensable for organisms for defense against the toxic effect of oxygen free radicals (Tripathi et al. 2006). At lower concentrations (≤10 μg L−1) of LEV, the CAT activity showed a slight increase, whereas the activity of SOD slightly decreased, both showing no significant differences with the control. This illustrated that M. flos-aquae tolerated LEV by other mechanisms, the low concentrations of LEV did not stimulate the antioxidant system to scavenge ROS, or ROS produced by LEV did not obviously affect the metabolisms of algae (Xie et al. 2011). The SOD activity increased significantly (P < 0.01) after exposure to 40 μg L−1 or higher concentrations of LEV. This may be attributed to the overproduction of superoxide, which is considered as the central component of the signal transduction, resulting in the activation of existing enzyme pools or increased expression of genes encoding SOD (Foyer et al. 1997; Mishra et al. 2006; Xie et al. 2011). In concurrence, the activity of CAT significantly increased, which was considered as an adaptive trait against the damage caused by oxidative stress (Rasheed and Mukerji 1991). In the present study, the activities of SOD and CAT were stimulated after exposure to higher concentrations of LEV and increased with increasing dose, indicating that the algal cells were under oxidative stress, resulting in the overproduction of ROS. However, the argument of SOD and CAT activities is still not enough to scavenge the ROS in algae, finally contributing to the inhibition of algal growth. A similar phenomenon was observed in the effect of tetracycline exposure on the growth of wheat (Triticum aestivum L.) (Xie et al. 2011) and the response of Chlorella vulgaris to trichloroisocyanuric acid (Nie et al. 2008).
MDA is an oxidized product of membrane lipids, and the content of MDA is commonly considered as an important index of lipid peroxidation to reflect cellular oxidative damage under environmental stress (Bailly et al. 1996; Chaoui et al. 1997). In the present investigation, the MDA content in algae exposed to 10 μg L−1 or higher concentrations of LEV was significantly higher than that in the control (P < 0.01). The possible reason is that under the stress caused by higher concentrations of LEV, the balance between production and scavenging of ROS in algal cells was disturbed, resulting the overaccumulation of ROS, which triggered the peroxidation of membrane lipids, increased lipid peroxidative products and oxidative stress in algae, damaged the membrane systems and functions, and increased the membrane permeability, finally leading to the death of cells (Alaiz et al. 1999). However, there was a slight increase in the MDA content at LEV concentrations lower than 1 μg L−1. The relatively low MDA content indicated less oxidative stress, which may account for no inhibition of algal growth (Xie et al. 2011).
Conclusion
The present study investigated the effects of LEV on M. flos-aquae and its mechanism. The results indicated that LEV could promote the growth of M. flos-aquae at lower concentrations and inhibit it at higher concentrations. After exposure to higher concentrations of LEV, the chlorophyll a content and photosynthetic activity decreased significantly, and the SOD and CAT activities and MDA content also increased obviously. These results may confirm that higher concentrations of LEV can cause photosynthesis inhibition and serious oxidative stress on M. flos-aquae. However, at lower concentrations, the photosynthetic activity increased slightly and the SOD and CAT activities and MDA content were not obviously different from those of the control. Therefore, further studies should be conducted to understand the mechanism of promotion on algae by lower concentrations of antibiotics.
References
Alaiz M, Hidalgo FJ, Zamora R (1999) Effect of pH and temperature on comparative antioxidant activity of nonenzymatically browned proteins produced by reaction with oxidized lipids and carbohydrates. J Agric Food Chem 47(2):748–752
Alberte RS, Friedman AL, Gustafson DL, Rudnick MS, Lyman H (1981) Light-harvesting systems of brown algae and diatoms. Isolation and characterization of chlorophyll a/c and chlorophyll a/fucoxanthin pigment–protein complexes. BBA 635(2):304–316
Bailly C, Benamar A, Corbineau F, Come D (1996) Changes in malondialdehyde content and in superoxide dismutase, catalase and glutathione reductase activities in sunflower seeds as related to deterioration during accelerated ageing. Physiol Plant 97:104–110
Chaoui A, Mazhoudi S, Ghorbal MH, Ferjani E (1997) Cadmium and zinc induction of lipid peroxidation and effects on antioxidant enzyme activities in bean (Phaseolus vulgaris L.). Plant Sci 127:139–147
Davis TW, Berry DL, Boyer GL, Gobler CJ (2009) The effects of temperature and nutrients on the growth and dynamics of toxic and non-toxic strains of Microcystis during cyanobacteria blooms. Harmful Algae 8(5):715–725
Foyer CH, Lopez-Delgado H, Dat JF, Scott IM (1997) Hydrogen peroxide and glutathione associated mechanism of acclamatory stress tolerance and signaling. Physiol Plant 100:241–254
Gao J, Sun MZ, Wang QY (2007) Effects of copper ion on the growth of Isochrysis Zhanjianggensis. Mar Fish Res 4(28):54–58 (in Chinese)
Ge F, Xu Y, Zhu RL, Yu FQ, Zhu MJ, Wong MH (2010) Joint action of binary mixtures of cetyltrimethyl ammonium chloride and aromatic hydrocarbons on Chlorella vulgaris. Ecotoxicol Environ Saf 73(7):1689–1695
Geoffroy L, Dewez D, Vernet G, Popovic R (2003) Oxyfluorfen toxic effect on S. obliquus evaluated by different photosynthetic and enzymatic biomarkers. Arch Environ Contam Toxicol 45(4):445–452
Giannoplitis CN, Ries SK (1977) Superoxide dismutase purification and quantitative relationship with water-soluble protein in seedlings. Plant Physiol 59:315–318
Goth L (1991) A simple method for determination of serum catalase activity and revision of reference range. Clin Chim Acta 196(2–3):143–151
Han BP, Han ZG, Fu X (2003) Algae photosynthesis mechanism and model. Science Press, Beijing (in Chinese)
Hegedüs A, Erdei S, Horvàth G (2001) Comparative studies of H2O2 detoxifying enzymes in green and greening barley seedling under Cd stress. Plant Sci 160:1085–1093
Jiang Y, Ji B, Wong RNS, Wong MH (2008) Statistical study on the effects of environmental factors on the growth and microcystins production of bloom-forming cyanobacterium—Microcystis aeruginosa. Harmful Algae 7(2):127–136
Krause GH (1988) Photoinhibition of photosynthesis: an evaluation of damaging and protective mechanisms. Physiol Plant 74:566–574
Lalumera GM, Calamari D, Galli P, Castiglioni S, Crosa G, Faneli R (2004) Preliminary investigation on the environmental occurrence and effects of antibiotics used in aquaculture in Italy. Chemosphere 54(5):661–668
Leu E, Krieger-Liszkay A, Goussias C, Gross EM (2002) Polyphenolic allelochemicals from the aquatic angiosperm Myriophyllum spicatum inhibit photosystem II. Plant Physiol 130(4):2011–2018
Liang Y, Feng LX, Tian CY, Yin CL (2006) Effects of salt stress on the growth and chlorophyll fluorescence of Pyramidomonas sp. Period Ocean Univ China 36(5):726–732 (in Chinese)
Liu BY, Liu WQ, Nie XP, Guan C, Yang YF, Wang ZH, Liao W (2011) Growth response and toxic effects of three antibiotics on Selenastrum capricornutum evaluated by photosynthetic rate and chlorophyll biosynthesis. J Environ Sci China 23(9):1558–1563
Liu WH, Ming Y, Huang ZW, Li P (2012) Impacts of florfenicol on marine diatom Skeletonema costatum through photosynthesis inhibition and oxidative damages. Plant Physiol Biochem 60:165–170
Mishra S, Srivastava S, Tripathi RD, Govindarajan R, Kuriakose SV, Prasad MNV (2006) Phytochelatin synthesis and response of antioxidants during cadmium stress in Bacopa monnieri L. Plant Physiol Biochem 44(1):25–37
Nie XP, Lu JY, Li X, Yang YF (2007) Toxic effects of norfloxacin on the growth and activity of antioxidase of Chlorella pyrenoidosa. Asian J Ecotoxicol 2(3):327–332 (in Chinese)
Nie XP, Wang X, Chen J, Ziko V, An T (2008) Response of the freshwater alga Chlorella vulgaris to trichloroisocyanuric acid and ciprofloxacin. Environ Toxicol Chem 27(1):168–173
Pan XL, Zhang DY, Chen X, Mu GJ, Li LH, Bao AM (2009) Effects of levofloxacin hydrochloride on photosystem II activity and heterogeneity of Synechocystis sp. Chemosphere 77(3):413–418
Qian HF, Li JJ, Pan XJ, Sun ZQ, Ye CB, Jin GQ, Fu ZW (2012) Effects of streptomycin on growth of algae Chlorella vulgaris and Microcystis aeruginosa. Environ Toxicol 27(4):229–237
Rasheed P, Mukerji S (1991) Changes in catalase and ascorbic acid oxidase activities in response to lead nitrate treatments in mungbean. Indian J Plant Physiol 34:143–146
Razinger J, Dermastia M, Drinovec L, Drobne D, Zrimec A, Koce JD (2007) Antioxidative responses of duckweed (Lemna minor L.) to short-term copper exposure. Environ Sci Pollut Res 14(3):194–201
Richardson BJ, Larn PKS, Martin M (2005) Emerging chemicals of concern: pharmaceuticals and personal care products (PPCPs) in Asia, with particular reference to Southern China. Mar Pollut Bull 50(9):913–920
Robinson AA, Belden JB, Lydy MJ (2005) Toxicity of fluoroquinolone antibiotics to aquatic organisms. Environ Toxicol Chem 24(2):423–430
Rohacek K, Bartak M (1999) Technique of the modulated chlorophyll fluorescence: basic concepts, useful parameters, and some applications. Photosynthetica 37(93):339–363
Santos LHMLM, Araujo AN, Faehini A, Pena A, Delerue-Matos C, Montenegro MCBSM (2010) Ecotoxicological aspects related to the presence of pharmaceuticals in the aquatic environment. J Hazard Mater 175(1–3):45–95
Schreiber U, Bilger W, Neubauer C (1994) Chlorophyll fluorescence as a nonintrusive indicator for rapid assessment of in vivo photosynthesis. In: Schulze E-D, Caldwell MM (eds) Ecophysiology of photosynthesis. Part A. Springer, Berlin, pp 49–70
Tan JH, Tang CM, Yu YY, Peng XZ (2007) Simultaneous analysis of multiple classes of antibiotics in urban river water by high performance liquid chromatography. Chin J Chromatogr/Zhongguo hua xue hui 25(4):546–549
Tang XX, Li YQ (2000) Inhibition of antioxidants on monocrotophos damage to Platymonas sp. Chin J Environ Sci 21(1):87–89
Tripathi BN, Mehta SK, Amar A, Gaur JP (2006) Oxidative stress in Scenedesmus sp. during short- and long-term exposure to Cu2+ and Zn2+. Chemosphere 62(4):538–544
Wise R (2002) Antimicrobial resistance: priorities for action. J Antimicrob Chemother 49(4):585–586
Xie XJ, Zhou QX, Lin DS, Guo JM, Bao YY (2011) Toxic effect of tetracycline exposure on growth, antioxidative and genetic indices of wheat (Triticum aestivum L.). Environ Sci Pollut Res 18(4):566–575
Xing W, Huang WM, Liu GH (2010) Effect of excess iron and copper on physiology of aquatic plant Spirodela polyrrhiza (L.) Schleid. Environ Toxicol 25(2):103–112
Yang S, Carlson KH (2004) Solid-phase extraction–high-performance liquid chromatography–ion trap mass spectrometry for analysis of trace concentrations of macrolide antibiotics in natural and wastewater matrices. J Chromatogr A 1038:141–155
Ye JP, Zhou SC, Zhang G, Xu WH (2007) Characteristics of selected antibiotics in the aquatic environment of the Pearl River Delta, south China. Ecol Environ 16(2):384–388 (in Chinese)
Yue XL, Zhang XP, Hu XW, Dong YY (2006) Effect of bensulfuron-methyl on growth of Chlorella pyrenoidosa. Sci Agric Sin 39(9):1823–1827 (in Chinese)
Acknowledgments
This study was supported by the National Natural Science Foundation of China (no. 20777021), the Science and Technology Research Key Project of the Ministry of Education (no. 210253), and the Natural Science Foundation Project of Fujian, China (no. 2010J01043, D0610012).
Author information
Authors and Affiliations
Corresponding author
Additional information
Responsible editor: Philippe Garrigues
Rights and permissions
About this article
Cite this article
Wan, J., Guo, P. & Zhang, S. Response of the cyanobacterium Microcystis flos-aquae to levofloxacin. Environ Sci Pollut Res 21, 3858–3865 (2014). https://doi.org/10.1007/s11356-013-2340-3
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s11356-013-2340-3